
粗壮脉纹孢菌发酵前后豆渣多糖理化性质和结构特性
2022-02-11 11:26:26黄文利欧阳耀铭邓泽元范亚苇
食品工业科技 2022年3期
黄文利,欧阳耀铭,卢 玲,邓泽元,范亚苇
(南昌大学,食品科学与技术国家重点实验室,江西南昌 330047)
豆渣是指在加工生产自制豆浆、豆腐等其他豆制品类的过程中产生的粗糙物或残渣,是大豆的副产物[1]。由于其易腐坏、有刺激性味道和质地差,常用做动物饲料或直接丢弃在田地里当肥料[2],但它不仅含有丰富的营养成分,如多种蛋白质、脂肪、矿物质等[3],还含有多种膳食纤维,其中约含有30%的可溶性多糖[4],可以有效促进人的肠道蠕动并可预防慢性便秘和结肠癌,减少脂肪在体内的沉积[5],减少糖尿病患者对胰岛素的消耗,并促进肠道有益菌群的生长、降低血液中胆固醇含量等功效[6]。目前,对豆渣中膳食纤维的提取处理方法有很多,如微波提取法[1]、酶水解法[7]、发酵等,其中,以豆渣为原料提取膳食纤维,利用羊肚菌[2]、酵母菌等微生物对豆渣进行发酵,可以提高发酵豆渣中营养成分的含量[8],相对于其他方法更能提高豆渣的利用率,而且节约资源,避免环境污染[9]。膳食纤维作为“第七类营养物质”成为当今社会食品保健方面重要的营养物质和食品研发对象[10]。
粗壮脉纹孢菌,属真菌门[11],分生孢子颜色呈红至棕黄色或淡黄至淡红橙色,球形或卵状椭圆形至卵状长形或球形,直径6~8 μm[12]。它是从江西传统食品中提取纯化出来的菌种[13],富含类胡萝卜素[14],能够产生高活性纤维素酶,能够在发酵过程中利用豆渣中的蛋白质、淀粉等物质,产生大量的有机酸和酶,使一些不溶性纤维可以降解,使豆渣口感变得细腻[15]。目前对粗壮脉纹孢菌发酵豆渣的研究主要集中在发酵后豆渣营养成分的测定和功能性糖的提取工艺[16],而对发酵前后豆渣理化性质及多糖结构解析的研究很少。有研究报道[17]用粗壮脉纹孢菌发酵的豆渣产品口感细腻,粗糙感明显降低,大部分纤维素被降解为功能性糖组分。发酵后由其多糖含量明显增加,且具有较高的抗氧化活性,可作为功能性食品使用[18]。然而,发酵对其多糖的结构是否发生变化仍不清楚。为了有效地利用发酵豆渣多糖,需要对其结构特性进行评估,对促进豆渣资源的深度开发至关重要。
本研究用粗壮脉纹孢菌发酵豆渣,对发酵前后豆渣中可溶性总糖、蛋白质、总酚、总黄酮理化成分进行测定;利用气相色谱-质谱联用仪分析发酵前后豆渣中多糖的单糖组成,并用微观扫描电子显微镜、X-射线衍射、傅里叶变换红外光谱、差热-热重分析对发酵前后多糖的结构进行进一步的分析鉴定,为今后发酵豆渣功能性糖的进一步研究提供依据。
1 材料与方法
1.1 材料与仪器
新鲜豆渣 南昌市江大南路农贸市场;粗壮脉纹孢菌 南昌大学国家重点实验室保存;考马斯亮蓝G-250 生兴生物技术有限公司;D-葡萄糖、L-鼠李糖、L-岩藻糖、D-半乳糖、L-甘露糖、L-阿拉伯糖
标准品,上海阿拉丁生化科技有限公司;DEAE-52纤维素 索莱宝生化科技有限公司;无水乙醇、硫酸、盐酸、苯酚、福林酚、氢氧化钠、氯化钠、碳酸钠、氯仿、正丁醇等 均为国产分析纯,广东省湛江科铭科技有限公司。
海能K 9860全自动凯氏定氮仪 厦门精艺兴业科技有限公司;LGJ- 1 AZ 真空冷冻干燥机 北京亚泰科隆仪器技术有限公司;Nicolet 5700 智能型傅里叶变换红外光谱仪 美国热电尼高力公司;JSM 6701 F 场发射扫描电镜带能谱仪 日本电子仪器有限公司;L6 s紫外可见分光光度计 上海光谱仪器有限公司;Agilent 6890 N气相色谱仪 安捷伦科技有限公司(中国)。
1.2 实验方法
1.2.1 粗壮脉纹孢菌发酵前后豆渣多糖的提取和测定 菌种斜面培养:称取40 g马铃薯葡萄糖琼脂(PDA)溶于100 mL蒸馏水中,水浴加热15 min,将灭菌过的琼脂倒入试管中制成斜面,然后挑取要接种的菌丝,将菌丝呈Z字型轻轻划斜面试管完成接种,最后放入温度30 ℃、湿度70%的恒温恒湿箱光照培养96 h,备用。
豆渣发酵:称取40 g豆渣于发酵瓶中,以豆渣:蒸馏水:菌液比1:2.1:0.4加入蒸馏水和菌液,然后放于恒温恒湿箱培养72 h,然后烘干粉碎备用。参照魏长浩等[19]的方法。
多糖的提取:称取适量用粗壮脉纹孢菌发酵前后的豆渣样品,分别将豆渣样品和蒸馏水以料液比1:20于95 ℃的水浴中浸提2 h,以转速4200 r/min离心5 min取上清液,将离心后沉淀继续浸提,重复上述操作,连续浸提三次。合并提取的溶液,将提取液快速浓缩至原溶液体积的1/4,加入4倍量75%的无水乙醇,4 ℃醇沉后经过夜24 h,醇沉后溶液进行离心得出多糖沉淀,然后将离心出的沉淀冷冻干燥后即可得到粗多糖[20],测定、计算粗多糖的得率。

1.2.2 发酵前后豆渣组成成分及多糖理化性质的测定
1.2.2.1 组成成分的测定 对粗壮脉纹孢菌发酵前后豆渣中总糖、蛋白质、总黄酮、总酚含量进行分析测定。采用苯酚-硫酸法[21]测总糖含量,以葡萄糖为标准品,得标准曲线y=0.0081x-0.0128,R2=0.9987;采用凯氏定氮法[22]测定蛋白质的含量;亚硝酸钠-硝酸铝法[23]测定总黄酮的含量,以芦丁为标准品,得标准曲线为y=0.0007x+0.002,R2=0.9997;采用福林酚试剂法[24]测定总酚的含量,以没食子酸为标准品,得标准曲线y=0.0066x+0.1163,R2=0.9992。
1.2.2.2 酶-Sevag法除粗多糖中的蛋白 取适当的粗多糖样品于锥形瓶中,料液比1:20添加蒸馏水,添加2%木瓜蛋白酶,在50 ℃摇床振摇1 h,使粗多糖中的蛋白质在蛋白酶的作用下充分水解,90 ℃水浴锅灭酶活15 min[25],离心取上层清液,然后添加1/4倍氯仿和正丁醇的混合液(氯仿:正丁醇的比例为4:1,v/v),振摇25 min,4500 r/min下离心5 min,将水相与氯仿相分开,水相部分再加入氯仿-正丁醇溶液,重复上述过程,直至水相和氯仿相之间无蛋白质为止,收集水相部分,浓缩、醇沉后离心取沉淀,冷冻干燥后即为脱完蛋白后的豆渣多糖(SRP),通过上述方法得到的未发酵豆渣的多糖记录为U-SRP,发酵豆渣的多糖记录为F-SRP。

式中:A0表示样品除蛋白前样品蛋白含量;A1表示样品除蛋白后蛋白含量;B0表示样品除蛋白前样品中多糖含量;B1表示样品除蛋白后样品中多糖含量。
1.2.3 发酵前后豆渣中多糖结构性能的测定
1.2.3.1 扫描电镜微观结构 用扫描电子显微镜可以观察发酵前后豆渣多糖的微观分子结构。将导电带贴在样品台上,将样品轻轻散落在导电带上,直至绿豆粒大小即可。用洗耳球轻轻吹去多余的样品粉末,然后用金属离子溅射仪进行镀金。用电子发射激光扫描电子显微镜(SEM)可以观察实验样品粉末的外观表面形貌,选择放大倍数2000×对样品进行表面结构观察并拍照。
1.2.3.2 X-射线衍射 利用X-射线衍射对发酵前后多糖的晶体结构进行分析。称取0.1 g发酵前后的多糖样品,试样测定条件为:管内电压40 kV、管内电流40 mA、衍射角范围为5°~50°。
1.2.3.3 傅里叶变换红外光谱 取1 mg发酵前后多糖样品粉末,在加热灯下加入2 mg KBr粉末,在玛瑙研钵中研磨成粉,尽量使粒度在2.0 μm以下,将研磨好的粉末在专用压片模具上加压成均匀透明的薄片,使其厚度约1 mm,然后进行红外光谱扫描,对在4000~500 cm-1波数内的峰型进行观察,得到扫描曲线。
1.2.3.4 单糖组成的测定 采用气质-联用色谱法[26]检测发酵豆渣多糖的单糖组成。
多糖的水解:精密称取多糖样品2 mg于10 mL比色管中,加入2 mL(2 mol/L)TFA溶液,充入一定量氮气后密封,在110 ℃的低温烘箱中进行水解4 h,冷却数分钟后用氮气吹干去除其中残留的TFA溶液,然后将比色管密封备用。
水解物的衍生:盐酸羟胺10 mg,99.5% 的吡啶0.5 mL,分别加入到多糖水解物、各单糖标准品和混合单糖标准品中。密封后在90 ℃的高温热水浴中对其进行快速振荡即可使其快速反应30 min。冷却至室温后,加入0.5 mL 99% 的羟基乙酸酐,在90 ℃的高温热水浴中连续进行振荡30 min,反应产物为乙酸乙腈衍生物。将得到的多糖衍生物过0.22 μm Ny有机滤膜后待测,随后准备进行气质分析。
GC-MS条件:色谱柱 Aglient Hp-5毛细管柱(30 m×0.32 mm×0.25 μm),进样前用乙醇洗针3次;进样口温度280 ℃,柱温为220 ℃,FID检测器温度设为250 ℃,进样量为1.0 μL,分流比20 :1载气为氦气,流速设为1 mL/min;程序升温初始温度设定为160 ℃,保持恒温2 min,然后以2 ℃/min的速度升温至200 ℃,然后保持恒温3 min;离子源温度230 ℃,质量扫描范围m/z 35~600。
1.2.3.5 差热-热重分析 采用热重分析法分析多糖样品性能与温度间的关系。取5~10 mg左右发酵前后的多糖粉末样品,放入差热-热重联用分析仪(DSC-TGA)样品池中进行热稳定性测定。温度范围25~600 ℃,升温速度10 ℃/min,氮气排气流速50 mL/min。
1.3 数据处理
所有单次统计实验中的数据统计分为3次,数据以平均数值±标准差值的方式进行表示,采用单因素方差分析(ANOVA)和采用SPSS22.0软件进行单次实验统计检验数据性状分析,采用Duncan多重极差方法进行统计检验,确定各实验数据性状均值的显著性(P<0.05)。
2 结果与分析
2.1 粗壮脉纹孢菌发酵前后粗多糖的测定
采用热水浸提法提取粗壮脉纹孢菌发酵前后豆渣粗多糖,其粗多糖的得率由2.10%增加到了17.20%,豆渣经粗壮脉纹孢菌发酵后粗多糖含量是原物料的8.19倍,张玮等[27]研究发现,粗壮脉纹孢菌发酵豆皮产纤维素酶可降解纤维素,使还原糖产量提高了81.74%。说明发酵可以提高豆渣中的糖含量,由于豆渣发酵过程中,会产生了大量的有机酸和酶,而粗壮脉纹孢菌可以分泌纤维素酶和半纤维素酶[28],使一些不溶性的纤维素得以降解,从而增加了豆渣中可溶性糖的含量。
2.2 发酵前后豆渣组成成分分析
发酵前后豆渣中总糖、蛋白质、总黄酮、总酚含量见表1,经粗壮脉纹孢菌发酵后豆渣中各理化组分含量都显著升高(P<0.05)。叶俊等[3]发现,利用粗壮脉纹孢菌发酵豆渣可以使豆渣中粗纤维含量降低,其中可溶性总糖、蛋白质等营养物质含量均显著提高。Vong等[29]发现,利用Yarrowia lipolytica发酵豆渣可以提高豆渣的营养水平,还可以使一些促鲜味物质琥珀酸、谷氨酸显著增加。余玮等[28]研究发现粗壮脉纹孢菌通过产生纤维素酶来分解和转化纤维素,使一些大分子物质在酶的作用下分解为小分子物质,从而使发酵后一些物质的营养成分得到提高。说明发酵可能通过纤维素酶和蛋白酶的作用使豆渣中的粗纤维降解[19]。同时发酵会影响豆渣中异黄酮的结构[30],豆渣中的异黄酮都是以丙二酰基配糖体的形式存在,豆渣经过粗壮脉纹孢菌发酵后,产生纤维素酶可降解其配糖体生成异黄酮配基,从而发酵后会增加总黄酮的含量。经过酶-Sevag法对粗多糖脱蛋白得到蛋白质的去除率为88.5%;多糖的保留率为86.25%,经脱蛋白后的多糖纯度更高,为后面更好的研究发酵前后多糖的结构性能奠定基础。

表1 发酵前后豆渣中基本成分测定结果(%)Table 1 Determination of basic components in soybean dregs before and after fermentation (%)
2.3 发酵前后豆渣中多糖结构性能分析
2.3.1 扫描电子显微镜(SEM) 图1分别为U-SRP和F-SRP的SEM照片。扩大2000×的SEM图像可以清晰地显示U-SRP和F-SRP的表面微观结构变化。从图1可以观察到F-SRP的表面微观结构与U-SRP相比出现了明显的变化。图1a中的U-SRP电镜图可以看出,豆渣多糖由许多致密的小颗粒组成,就像蓬松的头发,小颗粒被大颗粒包裹,聚集成一团,表面蓬松柔软的结构更有利于微生物的接触;FSRP表面结构如图1b所示,发酵后多糖分布较分散、不聚团、较松散,可能是由于发酵过程中微生物生长或酶的代谢产生了小分子物质,使豆渣变得疏松柔软,增加了豆渣多糖的比表面积,这可能对提高多糖、多酚等生物活性物质的提取率有一定的贡献。
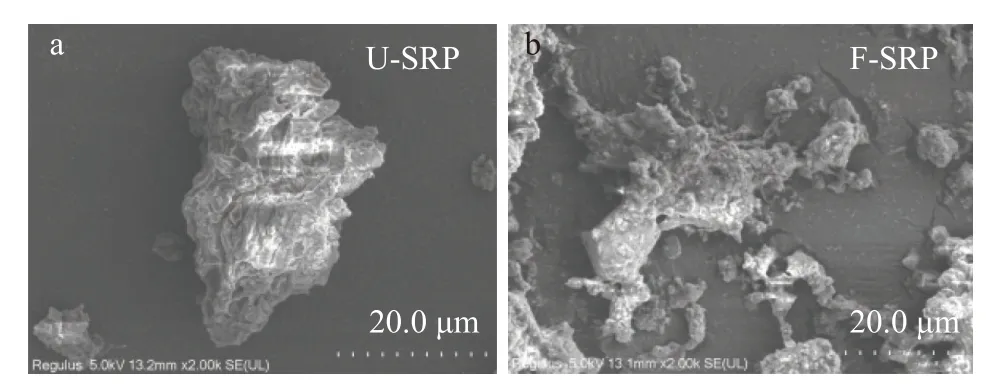
图1 发酵前后豆渣多糖扫描电镜图Fig.1 SEM graphs of soybean residue polysaccharide before and after fermentation
2.3.2 X-射线衍射 通过分析X射线在实验晶体中衍射产生的晶体衍射效应现象,来帮助检测实验样品是否具有的晶体结构。由如上图2可以明显看出,U-SRP在5°~50°衍射范围内无较明显的吸收峰出现,说明SRP中纤维分子具有牢固的结晶构型,有文献报道[31]纤维素由晶体和非晶体组成。在纤维素的结晶部分,纤维分子整齐的排列、有排列规律,葡萄糖纤维分子排列中的羟基与纤维分子内外的氢离子相互结合,形成对于纤维分子相对稳定的网状结晶分子结构。发酵后F-SRP在衍射角为17.6°时出现明显特征衍射峰,说明发酵对样品颗粒内部的晶体结构产生了一定程度的破坏,说明发酵水解了豆渣中的纤维素或半纤维素,使SRP变得蓬松,晶体结构发生改变,这与微观扫描电镜结果一致。

图2 发酵前后多糖的X-射线衍射图Fig.2 X- ray diffraction of polysaccharides before and after fermentation
2.3.3 傅里叶变换红外光谱 用傅里叶红外光谱分析多糖等大分子有机物中红外光与官能团的振动频率有关。当多糖被红外光照射时,在红外光谱图上可以看到各官能团会出现不同的吸收峰,根据吸收峰所处的红外光波段,可推测出多糖具有的官能团种类[32]。图3所示的为U-SRP和F-SRP的傅里叶变换红外光谱,傅里叶红外光谱分析结果表明,复合材料中存在多种特征官能团。从曲线上可以看出,U-SRP和F-SRP的光谱峰吸收带现状相似,但吸收带的相对强度不同,F-SRP的吸收带强度更大。多糖样品在3420.81 cm-1处的宽峰是多糖内部发生一定程度的O-H的伸缩振动峰[33];2928.53 cm-1处的一组峰是由于糖类C-H键的伸缩振动而形成的;1647.11 cm-1为-COOH中的C-O吸收峰;1418.63 cm-1为C-H的弯曲振动峰;1000~1200 cm-1以环振动为主,可能与C-O-H和C-O-C的伸缩振动重叠有关;1031.11 cm-1处的峰证实了吡喃环构象的存在,说明两者都含有吡喃糖环;在890 cm-1有微弱的吸收峰,说明U-SRP和F-SRP内部结构有β-型糖苷键存在[34];696.70 cm-1峰为醇或酚O-H键的面外弯曲振动。在3420.81、2928.534、1418.63 cm-1这三个波段的吸收峰,即为糖类化合物的特征吸收峰[35]。由U-SRP和F-SRP的傅立叶红外光谱图可以看出,粗壮脉纹孢菌发酵前后对多糖的化学结构和官能团没有显著影响。

图3 发酵前后豆渣多糖的红外光谱Fig.3 IR spectra of soybean residue polysaccharide before and after fermentation
有文献报道[36]利用羊肚菌发酵豆渣,通过傅里叶红外光谱分析发酵前后豆渣多糖结构变化,发现发酵后不仅出现了上述的吸收峰,还发现了三个新的条带,在1456 cm-1处为脂肪族C-H的变形,表明豆渣多糖中存在芳香化合物。1364 cm-1处为CH3-的弯曲震动,616 cm-1处为硫酸盐的不对称变形。同时有研究发现利用乳酸菌和粗壮脉纹孢菌进行结合发酵豆渣,发酵前后多糖的吸收波段与本研究相似,发酵后结构没有发生变化,造成上述发酵后多糖结构发生差异的原因,可能是与不同菌种在发酵中所产生酶的种类、酶活力和发酵过程中的持续性等因素有关。
2.3.4 单糖组成分析 通过GC-MS对豆渣和发酵豆渣中多糖的单糖组分进行定性定量分析。图4为混合单糖标准品色谱图。从图5~图6中可以看出豆渣和发酵豆渣中主要含有鼠李糖、阿拉伯糖、岩藻糖、甘露糖、葡萄糖和半乳糖,U-SRP中含量最高的是半乳糖(42.80%),其次是阿拉伯糖、岩藻糖等,其各单糖含量比为Rham:Ara:Fuc:Man:Glc:Gal 为6.62:23.6:9.53:5.66:7.63:42.80;发酵豆渣中葡萄糖含量最高(26.83%),其次是阿拉伯糖、半乳糖等,各单糖组成百分比Rham:Ara:Fuc:Man:Glc:Gal为13.5:20.02:10.81:10.11:26.83:16.74。从表2中可以得知,豆渣多糖中含量相对较高的是半乳糖,有研究发现植物细胞壁有很多植物纤维,半纤维素是植物膳食纤维的其中一个主要构成组分,其化学结构中分子链较短,有些糖单元如 4-氧-甲基葡萄糖醛酸和半乳糖等有时以支链连接到主链上,经水解后可产生多种中性糖。而葡萄糖的合成主要是来自纤维素和淀粉的水解,发酵后由于纤维素的水解使得葡萄糖的含量最高。Romero等[37]研究发现经粗壮脉纹孢菌发酵豆渣可高产纤维素酶和半纤维素酶,发酵产生的纤维素酶可能水解豆渣中的纤维素产生葡萄糖,这与张玮等[27]研究的发酵糖中对单糖的分析相似,水解产物中大部分单糖为葡萄糖,本文的研究发现经过粗壮脉纹孢菌发酵后各单糖的组成重新排布,大部分单糖为葡萄糖。分析上述结果表明,发酵后多糖中的大部分单糖组分均在未发酵豆渣的基础上增加,这是由于发酵产生的纤维素酶会破坏纤维素,并使细胞壁产生破壁现象,将一些难溶或不溶性的组分进行水解,使细胞壁多糖和胞内多糖可溶性组分溶出,如果胶和一些可溶性中性糖等,使得发酵后多糖各组分含量增加,由于细胞壁纤维中葡萄糖组分最多,所以发酵后葡萄糖含量较高。

图4 混合单糖标准品色谱图Fig.4 GC-MS chromatogram of mixed monosaccharide standards
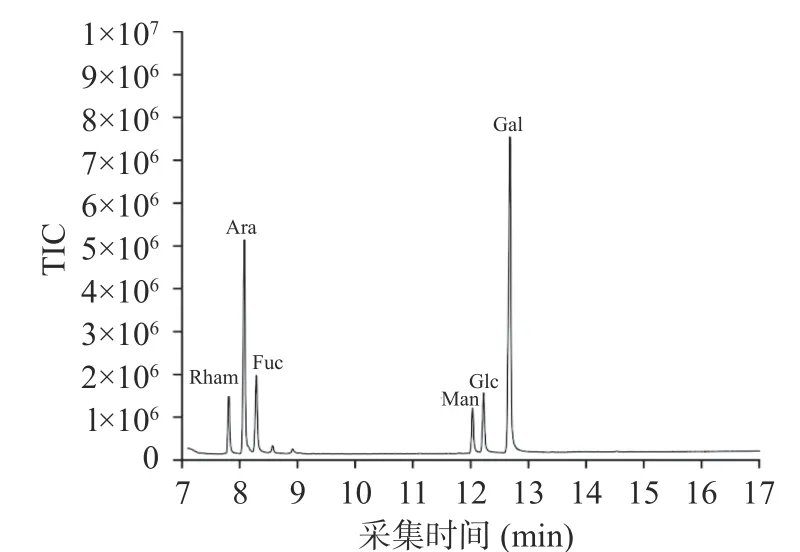
图5 未发酵豆渣多糖色谱图Fig.5 GC-MS Chromatogram of U-SRP

图6 发酵豆渣多糖色谱图Fig.6 GC-MS Chromatography of F-SRP

表2 发酵前后豆渣多糖的单糖组成Table 2 Monosaccharide composition of soybean dregs polysaccharides before and after fermentation
2.3.5 热稳定性 用热重分析评价样品的热稳定性,通过观察多糖样品随温度的变化而发生物理或化学的变化[38]。测量结果如图7所示,样品均呈多步热分解,热分解过程可以划分为两个阶段,第一阶段是200 ℃以前为初始热分解阶段,U-SRP在200 ℃之前失重为7.86%,F-SRP失重为10.4%。在100 ℃左右F-SRP损失比U-SRP多,在此期间是由于吸收水的蒸发和半水合物的脱水而失重,200 ℃时两者失重率相同。第二阶段发生在200~400 ℃,两者曲线都较陡,说明样品失重急剧降低,说明是样品中碳水化合物的降解所致,U-SRP比F-SRP失重较快,两者残余质量都有降低,但F-SRP残余质量相对较高,说明F-SRP比U-SRP具有较高的热稳定性。从图7中可以看出无论是U-SRP还是F-SRP在高温下质量都会有一定的损失。因此,建议食品加工温度应低于200 ℃,以避免改变样品的物理性能指标。

图7 发酵前后豆渣多糖热重分析Fig.7 TGA of soybean residue polysaccharides before and after fermentation
3 结论
本研究结果表明,采用粗壮脉纹孢菌发酵豆渣可以提高豆渣中多糖的含量,并且各理化组分含量都明显升高。根据扫描电镜和X-射线图谱,F-SRP较疏松分散、不聚团,发酵会破坏SRP的晶体结构。傅里叶红外光谱图可以看出发酵后含有多糖特征吸收峰,有β-型糖苷键存在,发酵对SRP化学结构和官能团没有显著影响。通过GC-MS对多糖中单糖组分进行定性定量分析,结果表明豆渣多糖中含量最高的是半乳糖,发酵豆渣中葡萄糖含量最高,发酵在酶的催化下使各种单糖组分发生改变,根据热重分析得到,发酵豆渣多糖比豆渣多糖具有较高的热稳定性。
本研究通过探究发酵前后豆渣中的组成变化,以及对发酵前后豆渣中多糖的结构进行解析,表明发酵后豆渣组成成分得到提高,多糖的微观结构得到了改善,具有作为优质膳食纤维的潜力。为粗壮脉纹孢菌发酵过程中所产生的植物多糖的实际应用研究提供了科学理论基础,对其进一步的应用研究以及今后开发出具有科学针对性的多种有机功能性保健食品起到了重要的研究指导意义。
猜你喜欢
东北农业大学学报(2020年3期)2020-05-14 13:18:12
粮食科技与经济(2020年2期)2020-05-09 10:13:58
祝您健康·文摘版(2019年12期)2019-12-13 08:24:29
生物技术通报(2019年2期)2019-03-15 05:52:02
天然产物研究与开发(2019年1期)2019-03-01 05:41:20
四川农业大学学报(2018年5期)2018-11-09 07:22:10
天然产物研究与开发(2016年1期)2016-06-05 10:29:25
现代面粉工业(2016年2期)2016-02-23 00:27:05
中南民族大学学报(自然科学版)(2015年2期)2015-12-16 12:11:10
中南民族大学学报(自然科学版)(2015年2期)2015-12-16 12:11:08
