
二氧化碳在华北大黑鳃金龟幼虫定位寄主植物根系中的作用
2022-02-10 23:33刘香雨赵旭王志敏秦健辉屈亚飞李克斌曹雅忠尹姣
植物保护 2022年1期
刘香雨 赵旭 王志敏 秦健辉 屈亚飞 李克斌 曹雅忠 尹姣





摘要
为探究二氧化碳(CO2)在华北大黑鳃金龟Holotrichia oblita幼虫定位寄主植物根系时所发挥的作用,对其开展了触角电位生理反应及行为生测试验。研究结果表明,CO2浓度为0.75%、1.5%、3%、6%时均能显著激发幼虫的触角电位和趋性反应,且随着浓度增加幼虫对CO2的趋向爬行所用时间显著缩短;根系分泌物同样能引起华北大黑鳃金龟幼虫的触角电位和趋性行为反应,且与1.5% CO2间无显著差异;将1.5% CO2分别与棕榈酸甘油酯、2丁烯酸、肉豆蔻酸、十四烷、4甲基2氧戊酸、十二烷、环己六醇、邻二甲苯和对羟基苯甲酸等9种根系分泌物混合后对试虫引诱效果增强,其趋向爬行时间显著缩短。以上结果显示,CO2在华北大黑鳃金龟幼虫定位寄主植物根系过程中发挥了重要的指引和增效作用,试验结果可为华北大黑鳃金龟幼虫的绿色防治提供新思路。
关键词
华北大黑鳃金龟幼虫;二氧化碳;植物根系分泌物;触角电位;行为反应
中图分类号:
S433.5
文献标识码:A
DOI:10.16688/j.zwbh.2020575
The role of carbon dioxide in the orientation of Holotrichia oblita larvae to the host plant root
LIU Xiangyu,ZHAO Xu,WANG Zhimin,QIN Jianhui,QU Yafei,
LI Kebin,CAO Yazhong,YIN Jiao*
(State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant
Protection, Chinese Academy of Agricultural Sciences, Beijing100193, China)
Abstract
In order to explore the role of carbon dioxide (CO2) in locating the roots of host plants by the larvae of Holotrichia oblita, the electroantennogram (EAG) and behavioral responses were assayed. The results showed that the stimulation of 0.75%, 1.5%, 3%, 6% CO2 elicited a strong reaction of the larvae, and their crawling speed became faster with increasing concentration of CO2. The root exudates could also induce an EAG and chemotaxis response of H.oblita larvae, and the results were similar to that of the group of 1.5% CO2. Furthermore, mixing 1.5% CO2 with nine kinds of root exudates, i.e., tripalmitin, 2butenoic acid, myristic acid, tetradecane, 4methyl2oxovaleric acid, dodecane, inositol, oxylene and phydroxybenzoic acid, respectively, could enhance the attraction effect and significantly shorten the crawling time. These results suggested that CO2 played an important role in guiding and enhancing the locating capability of the larvae of H.oblita for the host plant roots, which provides a new idea for green control of H.oblita larvae.
Key words
Holotrichia oblita larvae;carbon dioxide;plant root exudates;EAG;behavioral responses
蛴螬为鞘翅目金龟子幼虫的统称,是我国地下害虫中种类最多、发生为害最严重的类群[1]。其取食苗木根系,使其生长发育不良甚至死亡;或取食种子新萌发出的胚根、胚茎,使其不能萌发出土成苗,发生严重时可毁坏整块苗圃[2],而且苗木被咬食后,伤口处易感染病菌。据统計,植物地下部分受害有86%是蛴螬为害造成的[3]。另外,由于种植结构调整、免耕、浅耕、秸秆还田、有机肥推广等措施的实施,蛴螬在田间的为害日益严重[4]。华北大黑鳃金龟Holotrichia oblita对农作物、林木苗圃等为害的严重性以及广泛性在金龟子类群中居于首位,也是土栖金龟子中的一个优势种[5]。该虫一般1年发生1代,其3龄幼虫食量大,活动能力强,多在花生、大豆等油料和其他作物田的土壤中发生为害[6]。从东北到西北到西南再到华南,几乎全国范围都有华北大黑鳃金龟的分布[7]。由于蛴螬生活在土壤中,药剂难以直接接触到虫体,多年来主要采用高毒、高残留的化学药剂进行种子或土壤处理,对土壤和环境造成了巨大的污染,因此目前生产上急需绿色技术对蛴螬进行有效防控[8]。
害虫引诱技术由于具有对环境安全、选择性高、兼容性好等特点,已经成为当前植保领域研究热点[9- 10],并在多种重要农业害虫的田间种群监测和防治中得到了应用。目前引诱剂分为视觉引诱剂和嗅觉引诱剂[11]。视觉引诱主要是利用昆虫对某些颜色的趋性,比如荧光红色和黄色[12];嗅觉引诱则是利用一些植物挥发物、性信息素等化学物质对昆虫进行引诱[13 -14],比如,丙酸苯乙酯+丁香酚+香叶醇(3 ∶7 ∶3)的混合物在监测和诱杀日本金龟子Popillia japonica成虫中被广泛使用[13]。李晓峰等利用植物挥发物、性信息素、聚集信息素混配获得多个引诱华北大黑鳃金龟成虫的高效配方[15],为华北大黑鳃金龟的诱集防控提供了技术支持。但是,华北大黑鳃金龟主要通过幼虫取食植物根系造成危害,研究表明其对玉米、花生以及大豆根系都具有显著的趋向性[16],其中根系分泌物发挥了重要作用。植物根系分泌物是指在植物生长过程中,由根部释放到介质中的有机物质[17]。根系分泌物在植物与环境的互作中起传递信息的作用。不同作物根系分泌物的种类以及同种作物在不同生育期的根系分泌物的数量和种类都有差异[18]。已有研究表明,释放到根际的次生植物代谢物(如低分子量的醇类、酯类和酸类)对地下昆虫具有“引诱”特性,可以帮助土壤昆虫对其寄主植物进行定位和识别[19-20]。
二氧化碳(CO2)作为一种非特异引诱剂在土壤昆虫对寄主植物的定位中发挥着重要作用[21]。最典型的例子是玉米根萤叶甲 Diabrotica virgifera virgifera 对寄主植物的定位,Bernklau等[22]的研究表明, CO2对玉米根萤叶甲的最佳吸引浓度为2.51~4.20 mmol/mol,与玉米根附近土壤中CO2浓度(4.36±0.31) mmol/mol基本一致,300 mmol/mol 或900 mmol/mol的CO2浓度对玉米根萤叶甲表现出毒性。大量试验结果表明,玉米根萤叶甲仅根据CO2的浓度即可完成对玉米根的定位,并不需要其他物质的参与[23]。另外,针对浅黄根瘤象Sitona lepidus、甘蓝地种蝇 Delia radicum 等多种地下害虫的研究表明,害虫在检测到相应浓度的CO2后会引发其对寄主植物根系更为密集的搜索行为[19- 20, 24]。已有研究也表明,CO2是蛴螬寻找食物、定位寄主植物根系的重要线索[25],但是在对CO2和寄主植物根系的双向选择过程中,暗黑鳃金龟 Holotrichia parallela 和铜绿丽金龟 Anomala corpulenta 幼虫更趋向于植物根系[26]。目前对于CO2在华北大黑鳃金龟幼虫定位寄主植物过程中的作用尚未明确。
因此,本文旨在通过研究华北大黑鳃金龟幼虫对CO2和根系分泌物刺激的电生理反应和行为选择,探索CO2在华北大黑鳃金龟幼虫定位植物根系过程中发挥的作用,为开发新的华北大黑鳃金龟幼虫等地下害虫引诱剂提供理论依据。
1材料与方法
1.1供试昆虫
华北大黑鳃金龟取自中国农业科学院植物保护研究所地下害虫组实验室孵化的幼虫,置于养虫箱 (长36 cm×宽25 cm×高22 cm)内,将厚度为10 cm的灭菌土壤均匀地铺满箱底,湿度保持在(18±1)%, 室温控制在(25±1)℃,以马铃薯块及小麦苗为饲料进行饲养。
1.2供试材料
MY15 型触角电位仪(北京市科安劳保新技术公司);臂长25 cm的直管;“Y”形嗅觉仪主臂长20 cm,两个侧臂长18 cm,内径为3 cm(北京兴运科诺科贸中心);QC3大气采样仪(北京市科安劳保新技术公司);0.75%、1.5%、3%、6%(氮气填充)的CO2混配气体购于北京上通宏化工有限公司。
根系分泌化合物的选取参照马艳华等[16],共37种;分别购于上海阿拉丁生化科技股份有限公司、北京中创宏达科技有限公司、上海源叶生物科技有限公司、北京伊诺凯科技有限公司、北京百灵威科技有限公司、赛谱锐思(北京)科技有限公司、北京依珊汇通科技有限公司、北京凯国科技有限公司、生工生物工程(上海)股份有限公司,均为纯度≥95%的分析纯试剂。配制成浓度为0.1%的溶液,溶剂为石蜡油。
1.3华北大黑鳃金龟幼虫触角电位反应及行为试验
1.3.1试虫选择
选择健康、活泼、大小相同的3龄幼虫进行试验,试验前进行8~10 h的饥饿处理。
1.3.2 华北大黑鳃金龟幼虫对CO2和根系分泌物的触角电位反应
用镊子将幼虫触角完整地取下后用导电胶将触角轻轻地粘在两个电极上,保证只有触角两端与导电胶接触。分别观测华北大黑鳃金龟幼虫触角对CO2和植物根系分泌物的电生理反应。以石蜡油为对照,取20 μL(1 μg/mL)的待测化合物溶液(由石蜡油配制)涂于滤纸条上,并将滤纸条放入1 mL的蓝色枪头内,将枪头固定在电位仪上;每种化合物测试3根触角,同一化合物在1根触角上重复记录3次[27]。进行CO2浓度梯度试验时,分别将0.75%、1.5%、3%、6%的CO2气体装入一次性灭菌注射器中备用,每次给予刺激前,按照从低浓度到高浓度的顺序,依次将2 mL CO2气体打入枪头即可,每个CO2浓度重复测试10根触角。
1.3.3 華北大黑鳃金龟幼虫对CO2和植物根系分泌物的趋性行为反应
参照Honda[28]的方法,利用“Y”形嗅觉仪观测华北大黑鳃金龟幼虫对化合物和CO2的选择趋性。CO2与化合物的选择试验按照图1 顺序连接仪器,吸取20 μL(1 μg/mL)的标样化合物滴于滤纸上,放置在“Y”形管一侧臂端,另一侧臂端通1.5%的 CO2,“Y”形管两侧臂端分别通入经洗气瓶去除杂质和活性炭干燥的空气,气流保持在60 mL/min;每处理测试80头幼虫,各处理重复3次。将试虫从嗅觉仪的主臂口2 cm处引入,检查试虫位置,试虫若超过两侧臂结合中点到达某一侧臂 5 cm处时,即认为试虫选择了测试化合物或CO2;停留在主臂或未达到侧臂5 cm处的试虫定为未作出选择。
利用直管试验装置观测不同化合物或不同浓度CO2诱集对华北大黑鳃金龟幼虫爬行时间的影响。直管试验装置按照图2连接。 除不同浓度CO2的试验外,其余试验的CO2浓度为1.5%(即选择最接近植物根系的CO2浓度)[29]。气流保持在60 mL/min ,将饥饿处理后的幼虫放在距管口2 cm处,另一管口放置化合物或通CO2气体,试虫与化合物或CO2放置的距离为20 cm(界限),记录幼虫趋向爬行20 cm所需要的时间,为避免管中化合物残留以及幼虫爬行轨迹对于试验的影响,每观测10头幼虫要更换1次直管,更换化合物的同时也要进行直管的更换,每处理测试80头幼虫,重复3次。
1.3.4 华北大黑鳃金龟幼虫对CO2与植物根系分泌物混合体的趋性行为反应
利用“Y”形管观测根系分泌物添加CO2后对诱集华北大黑鳃金龟幼虫的影响(图1)。“Y”形管两臂内放入加有20 μL(1 μg/mL)相同化合物溶液的滤纸条,但“Y”形管一侧臂通入1.5% CO2,另一侧臂通入过滤后空气,其他试验方法同1.3.3。
利用直管试验装置观测根系分泌物添加CO2后(图2),两者的混合体在诱集过程中对试虫趋向爬行时间的影响,具体方法同1.3.3。
1.4数据处理
试验数据采用Excel 2016以及SPSS 16.0软件进行处理。采用t测验进行处理数据间的差异显著性分析。
2结果与分析
2.1华北大黑鳃金龟幼虫对不同浓度CO2的趋性反应
2.1.1 华北大黑鳃金龟幼虫对不同浓度CO2的触角电位反应
结果表明,华北大黑鳃金龟幼虫触角对4种不同浓度的CO2均有明显的电生理反应(图3a),随着CO2浓度的增加,触角电位反应逐渐增强,其中3%CO2与6%CO2均能引起华北大黑鳃金龟幼虫较强的触角电位 反应,并显著高于0.75%CO2引起的触角电位反应(P<0.05),但是与1.5%CO2无显著差异(P>0.05)。
2.1.2 华北大黑鳃金龟幼虫对不同浓度CO2的趋向爬行时间
幼虫对不同浓度CO2趋向爬行20 cm所需要的时间不同,从图3b中可以看出,CO2浓度越高,幼虫趋向爬行20 cm所需要的时间越短,其中6%CO2处理下爬行时间最短,仅需要75.4 s,极显著低于其在0.75%、1.5%和3% CO2浓度刺激下爬行20 cm所用时间(P<0.01)。该结果与幼虫对不同浓度CO2触角电位反应结果基本一致。
2.2华北大黑鳃金龟幼虫对植物根系分泌物的趋性反应
2.2.1 华北大黑鳃金龟幼虫对不同植物根系分泌物的触角电位反应
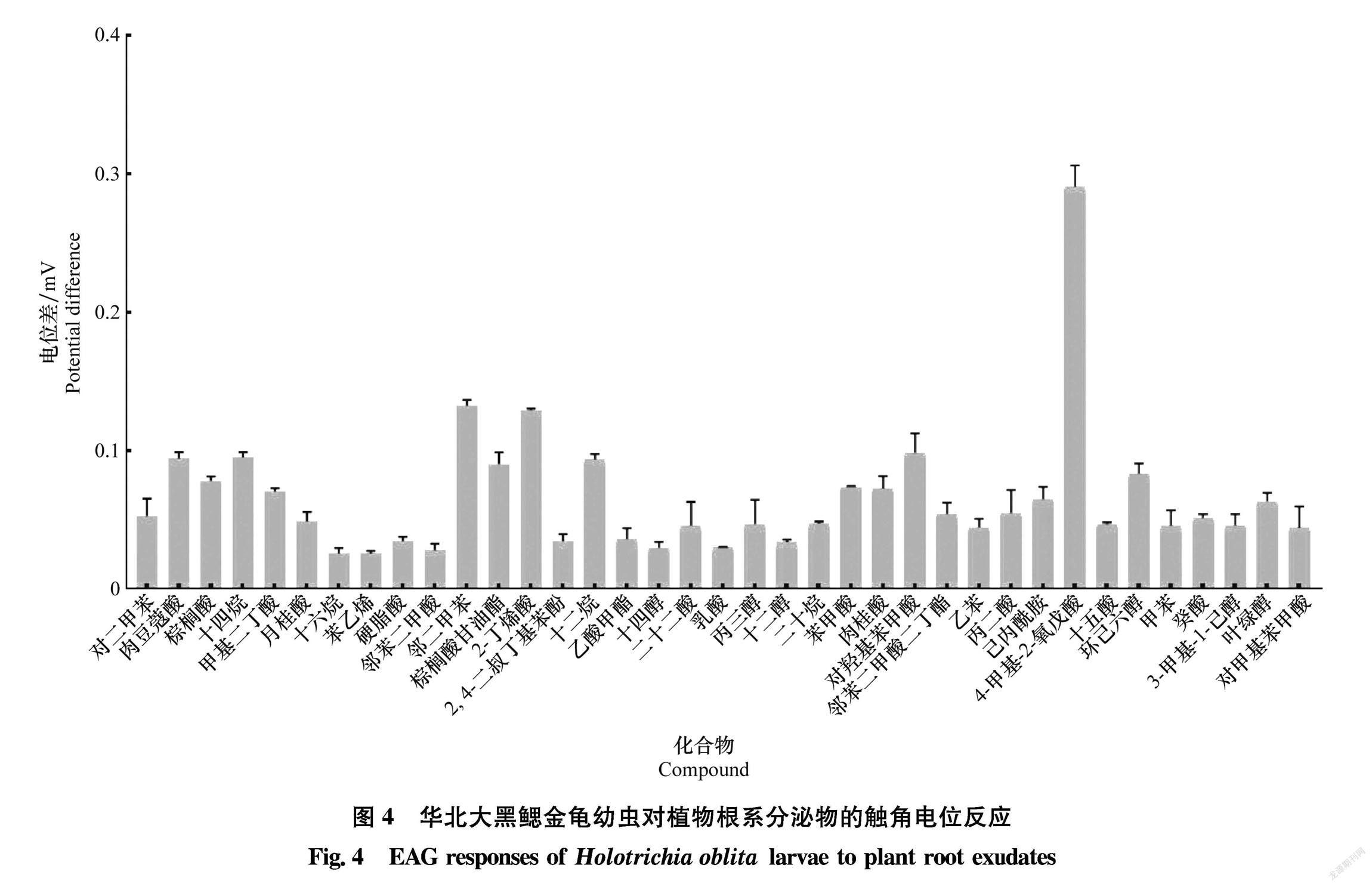
华北大黑鳃金龟幼虫对37种根系分泌物的电生理反应试验结果表明(图4),华北大黑鳃金龟幼虫对于不同根系分泌物的触角电位反应存在明显差异。其中,对于4甲基2氧戊酸、 邻二甲苯、2丁烯酸、对羟基苯甲酸、十二烷、十四烷、肉豆蔻酸、棕榈酸甘油酯、环己六醇等9种化合物的反应较明显(电位反应值均达到0.1左右),这些化合物中4甲基2氧戊酸引起的触角电位反应值最高(达到0.29左右)。
2.2.2不同植物根系分泌物对华北大黑鳃金龟爬行时间的影响
将上述能引起较强触角电位反应的9种化合物置于直管试验装置中对华北大黑鳃金龟幼虫进行诱集,试验结果表明,对羟基苯甲酸对华北大黑鳃金龟幼虫引诱效果最好(图5),试虫向其爬行20 cm仅需要(81±2)s,其次为十二烷,为(83±3)s。棕榈酸甘油酯诱集时幼虫爬行最慢,爬行20 cm需要133 s,顯著长于其他8种化合物诱集时幼虫的爬行时间(P<0.05),说明其引诱效果较差。
2.3华北大黑鳃金龟幼虫对CO2与植物根系分泌物混合物的趋性反应
2.3.1 华北大黑鳃金龟对CO2与根系分泌物的趋性选择
华北大黑鳃金龟幼虫对根系分泌物与1.5%CO2的趋性选择试验结果(图6)表明,幼虫除对环己六醇趋向性显著强于1.5%CO2,对其他8种化合物和CO2间的趋性选择没有显著差异(P>0.05),表明1.5%的CO2与绝大多数的植物根系分泌物之间对于试虫的诱集性无显著差异。
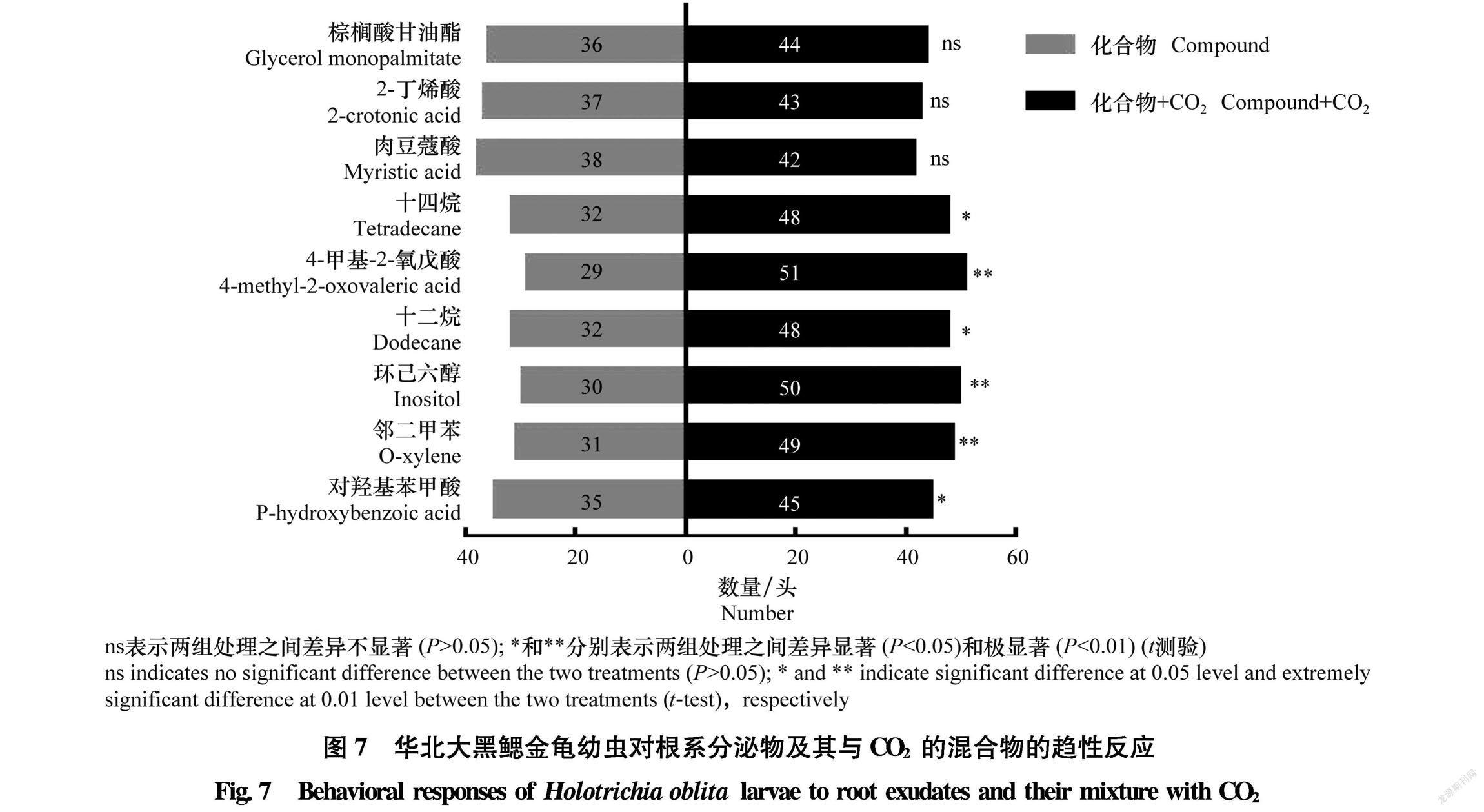
2.3.2 华北大黑鳃金龟对CO2与根系分泌物混合物的趋性选择
华北大黑鳃金龟幼虫对根系分泌物与添加CO2后混合物的双向选择试验结果(图7)表明,其明显更喜欢加入CO2的根系分泌物,且对6种添加CO2后的根系分泌物的趋向性显著(P<0.05)高于单纯的根系分泌物,其中,4甲基2氧戊酸、环己六醇、
邻二甲苯与CO2混合后对试虫的诱集 效果极显著高于相应化合物(P<0.01);说明CO2对根系分泌物引诱试虫具有显著的增效作用。
2.3.3 CO2与根系分泌物的混合物对华北大黑鳃金龟幼虫引诱爬行时间的影响
图8结果显示,将CO2添加到 9种植物根系分泌物中后均能够缩短幼虫趋向混合物爬行20 cm所需要的时间。其中,十二烷和对羟基苯甲酸添加CO2后幼虫爬行时间分别缩短20 s和24 s,与单一根系分泌物差异显著(P<0.05),而十四烷、4甲基2氧戊酸、环己六醇和邻二甲苯添加CO2后幼虫爬行时间分别缩短39、29、30 s和36 s,与不添加CO2差异极显著(P<0.01)。4甲基2氧戊酸在加入CO2气体后幼虫的趋向爬行时间最短,爬行20 cm仅需58 s。
3结论与讨论
本研究通过触角电生理反应和行为生测试验,初步探索了CO2对大黑鳃金龟幼虫的引诱作用,并比较了不同浓度下幼虫的反应差异,试验结果表明,低浓度(0.75%)的CO2对于华北大黑鳃金龟幼虫具有引诱作用,且随着CO2浓度的增加,对试虫的引诱效果也越强。此结论与张鑫鑫等[26]对暗黑鳃金龟和铜绿丽金龟的研究结果基本一致,表明植食性蛴螬对低浓度的CO2有明显的趋向性。目前针对玉米根萤叶甲、Ctenicera destructor等多种土壤昆虫定位寄主根系的研究也表明,昆虫主要利用CO2来定位寄主植物的根[19]。也有研究认为,CO2仅在昆虫寄主根系的定位过程中发挥辅助作用,如Zhang等[30]的研究认为CO2仅导致为害三叶草的浅黄根瘤象运动和搜索行为的变化,而并非是直接定位三叶草位置的信号;植物根系分泌物在某些土壤昆虫对寄主植物定位过程中发挥主导作用[20]。通过本研究也可以发现,植物根系分泌物如4甲基2氧戊酸、十二烷、环己六醇、十四烷、邻二甲苯、对羟基苯甲酸、棕榈酸甘油酯、2丁烯酸、肉豆蔻酸等化合物无论在触角电位反应还是趋性行为反应中,均对蛴螬表现出较强的刺激活性。因此,我们推测这些化合物可能在华北大黑鳃金龟幼虫识别寄主植物根系的过程中也发挥作用。此外,部分根系分泌物还具有驱避和抑制作用,它们在植物根受到破坏后被释放,可以阻止或减少食根昆虫对寄主植物根的取食[19]。还有研究认为,增加CO2浓度后,咀嚼式口器的昆虫为获得足够的氮素营养会增加取食[31]。
Doane等的研究表明,植物根系附近土壤中CO2的浓度大约为1.5%[29]。在华北大黑鳃金龟幼虫对1.5% CO2与9种植物根系分泌物的双向选择试验中发现,除环己六醇外,对1.5% CO2与其他8种化合物间的趋性选择并无显著差异。对多种土壤害虫的研究表明,CO2作为植物根释放量最大的气体[32],对土壤害虫具有远距离的引诱作用[33],而寄主植物根系分泌物则对华北大黑鳃金龟幼虫有近距离的引诱作用[16]。本文通过幼虫的行为选择和爬行时间的测定试验可知,CO2可增强根系分泌物对华北大黑鳃金龟幼虫的引诱效果,同时缩短其趋向爬行时间。因此,我们推测CO2作为非特异的化学信号,开启或者增强了华北大黑鳃金龟幼虫在土壤中对寄主植物根系的搜索行为,主要由于植物根系附近CO2浓度较没有根系的土壤中浓度要高,其指引幼虫向植物根系定向运动,在接近根系后植物根系分泌物开始发挥引诱作用,引导蛴螬等地下害虫定位并取食植物根部。这与浅黄根瘤象、甘蓝地种蝇、五月鳃金龟Melolontha melolontha 等土壤害虫对CO2和植物根系的定位搜索行为类似[19-20,24],即“CO2 是地下害虫觅食行为的触发器”。
本试验主要探究了CO2对于华北大黑鳃金龟幼虫定位寄主植物根系过程中的作用,结果表明,CO2对华北大黑鳃金龟幼虫有引诱或引导作用,且可以增强其对植物根系分泌物的趋向行为,此研究结果为探讨华北大黑鳃金龟幼虫对寄主根系的定位机制奠定了基础,也为华北大黑鳃金龟幼虫的绿色防控提供了新的思路。但是,本研究仅仅针对单一根系分泌物及其与CO2混合后对幼虫的吸引作用进行了研究,对于多种根系分泌物的混合物以及其与CO2混合后,对大黑鳃金龟幼虫的引诱作用及其作用程度等尚需开展深入研究。另外,本试验采用的CO2浓度约等于植物根系附近土壤中的CO2浓度。但事实上,由于CO2是无处不在的,作物地下块根、块茎、花生荚等也是土壤害虫主要为害的部位,其呼吸强度会高于其他根系,产生的CO2可能更多;还有土壤微生物释放的CO2等,都有可能对于土壤昆虫定位植物根系的行为产生影响,这些因素也有待后续开展进一步研究。
参考文献
[1]曹雅忠, 李克斌, 尹姣. 浅析我国地下害虫的发生与防治现状[C]∥成卓敏. 农业生物灾害预防与控制研究. 北京: 中国农业科学技术出版社, 2005: 5.
[2]吴瑾, 范颖博. 金龟子类林木害虫的发生与防治[J]. 现代农村科技, 2020(9): 38.
[3]张美翠, 尹姣, 李克斌, 等. 地下害虫蛴螬的发生与防治研究进展[J]. 中国植保导刊, 2014, 34(10): 20-28.
[4]INNOCENT N, ZHANG Shuai, SUN Xiulian, et al. Transition challenge to organic agriculture: A course for advancing belowground insect pest management [J/OL]. Applied Soil Ecology, 2020, 148: 103476. DOI: 10.1016/j.apsoil.2019.103476.
[5]劉福顺, 王庆雷, 刘春琴, 等. 大黑蛴螬活动规律及对农作物幼苗的取食趋性研究[J]. 环境昆虫学报, 2014, 36(4): 635-639.
[6]胡宏云, 许春远. 华北大黑鳃金龟生物学特性研究[J]. 安徽农业科学, 1986(4): 69- 70.
[7]王哲, 钟涛, 赵彤华, 等. 重要地下害虫东北大黑鳃金龟研究进展[J]. 环境昆虫学报, 2019, 41(5): 1023- 1030.
[8]陈建明, 俞晓平, 陈列忠, 等. 我国地下害虫的发生为害和治理策略[J]. 浙江农业学报, 2004, 16(6): 389-394.
[9]HARRIS C R. Factors influencing the effectiveness of soil insecticides [J]. Annual Review of Entomology, 1972, 17: 177-198.
[10] 蔡晓明, 边磊, 李兆群, 等. 田间背景气味对植物源引诱剂的干扰及相应研发策略[J]. 中国生物防治学报, 2020, 36(5): 646-655.
[11] WILLBRAND B N, PFEIFFER D G. Brown rice vinegar as an olfactory field attractant for Drosophila suzukii (Matsumura) and Zaprionus indianus Gupta (Diptera: Drosophilidae) in Cherimoya in Maui, Hawaii, with implications for attractant specificity between species and estimation of relative abundance [J/OL]. Insects, 2019, 10(3): 80. DOI: 10.3390/insects10030080.
[12] KIRKPATRICK D M, MCGHEE P S, HERMANN S L, et al. Alignment of spotted wing Drosophila (Diptera: Drosophilidae) on odorless disks varying in color [J]. Environmental Entomology, 2016, 45(1): 185- 191.
[13] LADD T L, MCGOVERN T P. Japanese beetle: a superior attractant, phenethyl propionate + eugenol + geraniol, 3 ∶7 ∶3 [J]. Journal of Economic Entomology, 1980, 73(5): 689 -691.
[14] RUTHER J, TOLASCH T. Attraction of garden chafer, Phyllopertha horticola, to floral Japanese beetle lure [J]. Journal of Economic Entomology, 2004, 128(2): 158 -160.
[15] 李晓峰, 曹雅忠, 尹姣, 等. 华北大黑鳃金龟植物源引诱剂配方筛选及引诱效果[J]. 植物保护学报, 2020, 47(1): 35- 45.
[16] 马艳华, 李雪, 曹雅忠, 等. 华北大黑鳃金龟幼虫对3种寄主植物根系分泌物的趋性反应[J]. 应用生态学报, 2018, 29(11): 3838- 3846.
[17] 王会会, 李前正, 王川, 等. 根系分泌物介导的根际效应及在水体生态修复中的应用潜力[J]. 水生生物学报, 2020, 44(2): 453- 460.
[18] 朱丽霞, 章家恩, 刘文高. 根系分泌物与根际微生物相互作用研究综述[J]. 生态环境, 2003, 12(1): 102- 105.
[19] JOHNSON S N, GREGORY P J. Chemicallymediated host plant location and selection by rootfeeding insects [J]. Physiological Entomology, 2006, 31(1): 1-13.
[20] JOHNSON S N, NIELSEN U N. Foraging in the darkchemically mediated host plant location by belowground insect herbivores [J]. Journal of Chemical Ecology, 2012, 38(6): 604-614.
[21] HAMILTON C C. The behaviour of some soil insects in gradients of evaporating powder of air, carbon dioxide and ammonia [J]. Biology Bulletin, 1917, 32(2): 159 -182.
[22] BERNKLAU E J, BJOSTAD L B. Behavioral responses of firstinstar western corn rootworm (Coleoptera: Chrysomelidae) to carbon dioxide in a glass bead bioassay [J]. Journal of Economic Entomology, 1998, 91(2): 444-456.
[23] BERNKLAU E J, BJOSTAD L B. Reinvestigation of host location by western corn rootworm larvae (Coleoptera: Chrysomelidae): CO2 is the only volatile attractant [J]. Journal of Economic Entomology, 1998, 91(6): 1331-1340.
[24] REINECKE A, MLLER F, HILKER M.Attractiveness of CO2 released by root respiration fades on the background of root exudates [J]. Basic Applied Ecology, 2008, 9(5): 568-576.
[25] HASLER T. Abundanzund dispersionsdynamik von Melolontha melolontha (L.) in intensivobstanlagen [M]. ETH zürich, 1986: 60-65.
[26] 张鑫鑫, 房迟琴, 刘丹丹, 等. 二氧化碳对三种蛴螬的引诱作用[J]. 植物保护, 2016, 42(6): 46-50.
[27] 李为争, 杨雷, 申小卫, 等. 金龟甲蓖麻源引诱剂的配方筛选及田间效果评价[J]. 中国生态农业学报, 2013, 21(4): 480-486.
[28] HONDA T, KAINOH Y, HONDA H. The persistence of a learned response in the egglarval parasitoid Ascogaster reticulatus Watanabe (Hymenoptera: Braconidae) [J]. Entomological Science, 1999, 2(3): 335-340.
[29] DOANE J F, LEE Y W, KLINGLER J, et al. The orientation response of Ctenicera destructor and other wireworms (Coleoptera: Elateridae) to germinating grain and to carbon dioxide [J]. The Canadian Entomologist, 1975, 107(12): 1233-1251.
[30] ZHANG Xiaoxian, JOHNSON S N, GREGORY P J, et al. Modelling the movement and survival of rootfeeding clover weevil, Sitona lepidus, in the rootzone of white clover [J]. Ecological Modelling, 2005, 190(1): 133-146.
[31] 孟玲, 李保平. 二氧化碳濃度升高对植食性昆虫的影响[C]∥李典谟, 康乐, 吴钜文, 等. 昆虫学创新与发展——中国昆虫学会2002年学术年会论文集. 北京: 中国科学技术出版社, 2002: 6.
[32] PAYNE D, GREGORY P J. The soil atmosphere [M]∥WILD A. Russell’s soil conditions and plant growth. Harlow, UK: Longman, 1988: 298-314.
[33] 涂书新, 孙锦荷, 郭智芬, 等. 植物根系分泌物与根际营养关系评述[J]. 土壤与环境, 2000, 9(1): 64 -67.
猜你喜欢
东方少年·阅读与作文(2020年2期)2020-06-11
小学科学(2019年7期)2019-08-27
小猕猴学习画刊(2019年4期)2019-05-08
扬子江(2019年1期)2019-03-08
新青年(2018年9期)2018-09-14
农民致富之友(2017年24期)2018-01-30
小朋友·快乐手工(2017年3期)2017-04-26
故事林(2017年5期)2017-03-18
作文评点报·作文素材小学版(2016年9期)2016-03-18
第二课堂(课外活动版)(2015年6期)2015-10-21
