
防褐化处理对蒙古栎插穗生根及其生理生化的影响
2022-02-10 12:35虞家昕王思进王天宠陆秀君
西北林学院学报 2022年1期
虞家昕,战 昊,王思进,王天宠,陆秀君
(沈阳农业大学 林学院,辽宁 沈阳 110866)
蒙古栎(Quercusmongolica)是壳斗科(Fagaceae)栎属(Quercus)落叶乔木,别名柞树、蒙栎,国家二级珍贵树种,是我国东北林区中主要次生林树种,其材质优良,也是我国30个主要工业用材林树种之一[1-2]。蒙古栎具有重要的生态建设价值,尤其材质优良的蒙古栎的橡实和柞叶更具有经济利用和很高的开发价值。
我国目前主要的蒙古栎林分绝大部分是采伐后萌蘖更新的次生林,其遗传改良程度差,通过优树选择和杂交选育应用无性繁殖的方法,是增加生产量、提高改良程度,加速良种化的重要手段[3]。栎树属于极难生根树种,其体内含有大量酚类等阻碍生根物质[4],其中主要抑制生根物质为单宁,单宁中含有很多的酚羟基,对光敏感,易被氧化成黑色的氧化物,插条基部褐化,从而导致插穗死亡。
蒙古栎属愈伤组织和皮部混合生根型且极难生根型树种。对难生根树种常采用激素处理、黄化处理、防褐化处理等方式提高扦插成活率。黄秦军等[3]将2种生长调节剂IBA和6-BA混合施用浸蘸插穗,使蒙古栎的生根率达到16.67%。祝亚云等[5]在研究弗吉尼亚栎(Q.virginiana)嫩枝扦插中发现,400 mg·L-1NAA+400 mg·L-1IBA 混合液处理效果最佳,生根率达82.6%。吕秀立等[6]对苏玛栎(Q.shumardii)扦插繁殖研究表明:采用0.5 mg·L-1的IBA浸泡1 h处理插穗效果最佳,生根率为24.2%。夏兴宏等[7]在麻栎(Q.acutissima)无性扦插繁殖的研究中表明,统一采用300 mg·L-1IBA+3 mg·L-16-BA处理插穗,以珍珠岩为基质的生根效果最佳,生根率为81.00%;正交试验结果显示,用300 mg·L-1IBA+ABT1处理0.5 h,扦插生根率达到88.67%。在实生苗上搭建遮阴网获得黄化的蒙古栎插穗,采用IBA速蘸插穗,生根率最高达到84.6%[4]。但是采用黄化处理需要建设大量遮阴棚为实生苗遮阴,成本高,处理时间长,有一定的推广局限性。李先民[8]采用丙酮、抗坏血酸对青钱柳(Cyclocaryapaliurus)插穗进行了防褐化处理,使生根率最高达到17.8%,显著高于对照处理,取得了良好的成果。朴楚炳等[9]采用酒精和高锰酸钾对红松(Pinuskoraiensis)插穗进行防褐化处理,使得生根率达到66.7%,显著高于对照。李明红等[10]采用250 mg·L-1丙酮+200 mg·L-1ABT处理杨梅(Myricarubra)插穗,生根率可达88.3%。
树种不同,提高扦插成活率的方法各异。本研究针对蒙古栎插穗褐化严重的现象,对其嫩枝插穗采用乙醇、AgNO3、抗坏血酸3种方法防褐化处理,并结合激素处理探究其对蒙古栎嫩枝扦插生根及其生根过程中相关酶活性及内源激素的影响,以掌握嫩枝扦插繁殖技术和影响扦插成活的内在因子,为扦插育苗提供技术支撑和科学依据。
1 材料与方法
1.1 试验材料
试验材料采自位于辽宁省沈阳市高坎镇的沈阳农业大学试验基地,采穗母株为3年生实生苗。2020年7月上旬选取生长良好的当年生半木质化枝条的中上部,上、下口平剪成10~15 cm的插穗,保留上部2片1/3叶。基质按V草炭∶V蛭石∶V珍珠岩=1∶2∶2混合后均匀铺在沙床上。塑料拱棚上搭建遮阴网,透光度35%~40%,拱棚内温度控制在28~30 ℃、湿度85%~95%,定期喷施杀菌剂与营养液。
1.2 试验方法
1.2.1 防褐化处理对插穗生根的影响 防褐化处理的试剂选用1%乙醇、2% AgNO3和不同浓度的抗坏血酸。将插穗在上述防褐化药剂中浸泡1 h后,再将插穗基部2~3 cm处采用500 mg·L-1IBA-K(吲哚丁酸钾)浸泡2 h后扦插(表1)。分别以500 mg·L-1IBA-K和清水浸泡2 h处理为对照。每种处理3次重复,每次重复30个插穗。采用完全随机区组设计。扦插后90 d调查生根率、愈伤率、愈伤组织平均重量、生根数、平均根长、最长根长及根系效果指数等指标。
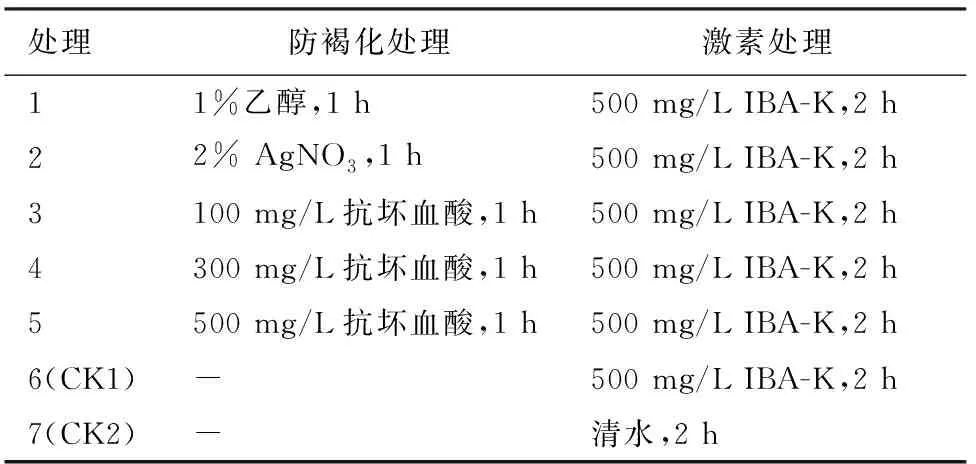
表1 防褐化处理试验设计Table 1 Anti-browning treatment experiment design
1.2.2 乙醇和IBA-K处理对插穗生根过程中氧化蛋白酶系统变化的影响 以清水浸泡处理插穗2 h后扦插为空白对照组,处理组采用1%乙醇1 h+500 mg·L-1IBA-K 2 h后扦插。定期随机取样,在扦插当天取样1次,扦插后每10 d取样1次,共计6次。
取样时将插穗基部韧皮部与木质部分离,剥离取2~3 cm韧皮部,放置于-80 ℃超低温冰箱保存备用。
测定处理组和对照组不同时期插穗韧皮部的氧化蛋白酶:过氧化物酶(POD)、多酚氧化酶(PPO)、吲哚乙酸氧化酶(IAAO)活性,过氧化物酶(POD)活性测定参照李合生[11]的方法;多酚氧化酶(PPO)活性测定参照郝再彬等[12]的方法;吲哚乙酸氧化酶(IAAO)活性测定参照高俊凤[13]的方法,3次重复。
1.2.3 乙醇和IBA-K处理对插穗生根过程内源激素变化的影响 测定处理组和对照组不同时期插穗韧皮部的内源激素:吲哚乙酸(IAA)、脱落酸(ABA)和玉米素核苷(ZR)含量,采用酶联免疫法(ELISA)测定,3次重复。
采用Excel 2016进行数据统计,采用SPSS 22.0进行方差分析,origin 8.5绘制图表。
2 结果与分析
2.1 防褐化处理对蒙古栎插穗生根的影响
蒙古栎嫩枝防褐化处理后再经IBA-K生根诱导处理可显著提高插穗的生根率,防褐化处理插穗的扦插生根效果明显优于CK2(表2)。蒙古栎插穗经1%乙醇和2% AgNO3处理后,生根率、最长根长和根系效果指数显著高于CK1和CK2,表明防褐化处理促进了不定根的发育伸长。其中,1%乙醇处理后的生根率、平均根长和根系效果指数最佳,分别为21.1%、8.5 cm和5.713。蒙古栎插穗经100 mg·L-1抗坏血酸处理后,生根数量最多,可见适宜浓度的抗坏血酸更有利于提高不定根生根的数量。而IBA-K处理的愈伤率达43.1%,显著高于其他处理,说明单独的IBA-K处理有利于愈伤组织的形成。可见,防褐化处理明显提高了蒙古栎嫩枝扦插的生根率和根系效果指数。
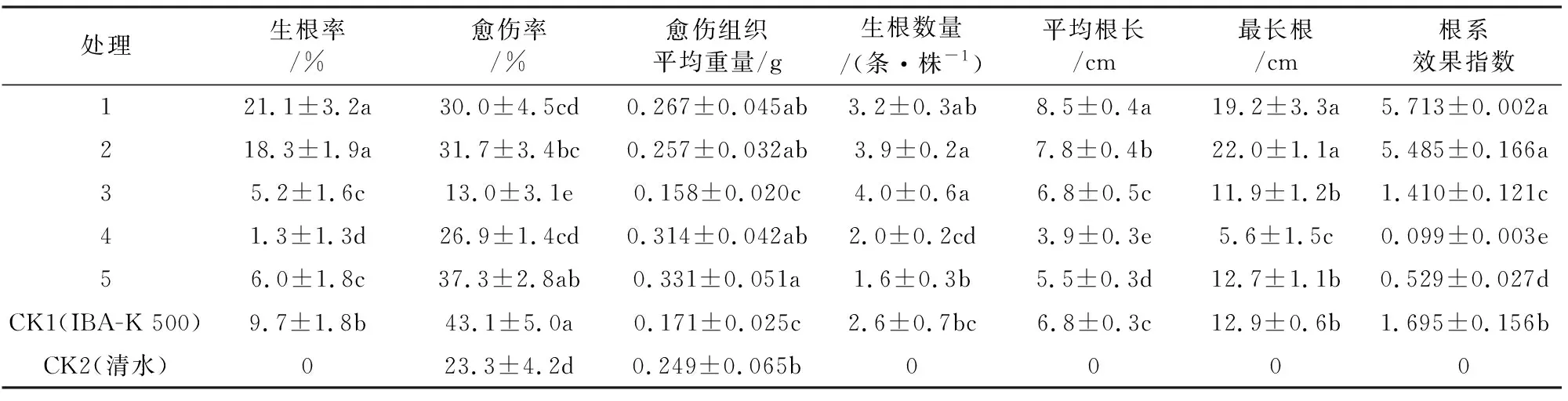
表2 防褐化处理对蒙古栎插穗生根性状的影响Table 2 Effects of preventing browning on rooting characters of Q.mongolica cuttings
2.3 蒙古栎插穗生根进程中氧化蛋白酶活性的变化
乙醇和外源激素的施加,促进了POD、PPO酶活性的增加,而使IAAO活性受到抑制。在插穗生根过程中POD活性呈上升趋势,且活性一直高于对照(图1)。生根初期0~10 d迅速增加,是因为外源激素的施加促进了POD活性上升,这有利于诱导基部皮部膨大形成内部愈伤组织;10~30 d保持相对平稳状态;30~40 d急剧增加,此时,插穗不定根大量形成并伸长,而POD活性明显增加有利于根部木质化所需的木质素的合成,有效促进根系大量形成和伸长。插穗生根进程中处理组的PPO活性始终大于对照(图2),说明外源激素处理促进了插穗PPO的活性。在0~10 d,PPO活性保持相对平稳状态;在20 d达到峰值,有利于插穗基部皮部膨大从而大量形成愈伤组织;在30~50 d,PPO活性又迅速上升,此时,不定根生成伸长,可见高活性PPO不仅促进愈伤组织的形成,还促进不定根大量生成和伸长生长。在插穗生根关键期中,PPO与POD维持高活性且呈现协同作用,从而促进不定根的发育伸长。
蒙古栎插穗IAAO的活性在生根进程各时期总体呈下降趋势,且始终小于对照(图3),说明外源激素处理有效降低了插穗IAAO的活性。在0~10 d活性迅速上升达到峰值,此时愈伤组织发育,这说明IAAO氧化插穗体内IAA,外源激素处理的插穗可以有效降低IAAO的活性,减少IAA的分解,有利于愈伤组织的发育;30 d后,处理组插穗IAAO活性维持在一个较低水平,促进了插穗不定根发育和伸长生长;而对照组一直处于较高水平,抑制了IAA的含量影响了不定根的发育生长。
在不定根生根关键期间,POD、PPO、IAAO 3种酶协同作用促进了不定根的生长,较高活性POD、PPO与维持在低活性的IAAO更有利于不定根在生根关键期的发育。
2.4 蒙古栎插穗生根进程中内源激素变化
内源激素比值变化能反映对蒙古栎插穗生根的调控机制。蒙古栎插穗在扦插初期IAA/ABA比值在处理组和对照组均开始下降(图4),因为插穗离开母体,受到切伤刺激,导致ABA的含量增加。在10~20 d愈伤组织大量形成期内,处理组比值上升接近峰值,对照组也大幅上升,较高的比值有利于蒙古栎愈伤组织发育和不定根原基的诱导。20 d后,处理组比值出现下降,此时不定根开始表达发育,消耗IAA,随后又迅速上升,此时是不定根大量生成时期,ABA降至初始水平,故比值上升;40 d后IAA/ABA值下降,此时不定根伸长生长。在蒙古栎插穗的生根关键期,处理组插穗的IAA/ABA值始终高于对照组,可见外源激素在一定程度上提高了插穗IAA/ABA的比值。IAA/ZR比值整体上呈现出“下降-上升-下降-上升”的趋势(图5)。扦插前期的0~20 d,处理组IAA/ZR比值先急剧降低而后上升,对照组相比于处理组变化较小,都在20 d达到峰值,有利于愈伤组织的大量生成及不定根原基的诱导。20~30 d不定根开始发生,IAA含量相对稳定,而ZR含量开始上升,高含量的ZR有利于不定根的发育,故IAA/ZR值下降。30 d后不定根开始大量生成,IAA含量和ZR含量的上升有利于不定根的伸长发育,处理组与对照组的IAA/ZR比值上升而后缓慢下降,恢复正常水平,但对照组变化幅度始终较处理组小。可见,较高的IAA/ABA值和较低的IAA/ZR值有利于蒙古栎嫩枝插穗生根。
3 结论与讨论
在生根过程中,防褐化处理有效提高了蒙古栎插穗的扦插生根率。1% 乙醇1 h浸泡结合500 mg·L-1IBA-K激素处理,通过提高POD与PPO的活性,降低IAAO的活性,增加IAA/ZR和IAA/ABA的比值,提升蒙古栎嫩枝扦插成活率。林木扦插生根诱导过程受到生理、形态、生化等多方面因素的影响,而植物生长调节剂和插穗处理方式是关键的制约因素[6]。对于难生根的栎类树种影响插穗生根的因素有很多[14-17],其中植物生长调节剂处理是促进难生根树种插穗生根的重要技术方法,不仅有利于根原始体的诱导,而且能够促进不定根的发育和生长[18]。同时可以增加组织再生能力,提高酶活性诱导根源基的发生,从而有利于植物扦插繁殖形成不定根[19],但不同种类对植物插穗生根效果的影响不同。栎树体内含有酚类等阻碍生根的物质,属扦插极难生根树种,其中主要抑制扦插生根的物质为单宁[8],而蒙古栎枝条富含单宁,单宁中含有很多的酚羟基,对光很敏感,且极易被氧化成黑色的氧化物,容易导致插条基部褐化,从而导致插穗死亡,阻碍插穗生根,所以对插穗的预处理方式尤为重要。本研究发现,采用1%乙醇和2% AgNO3进行浸泡处理后再用IBA-K浸蘸的生根效果要显著好于单独使用IBA-K处理的效果,通过1% 乙醇处理1 h和500 mg·L-1IBA-K处理2 h的防褐化处理可以使生根率达到21.1%。
在蒙古栎嫩枝插穗生根进程中,外源激素IBA-K的处理能够一定程度上提升插穗内POD、PPO的活性,降低IAAO的活性,均有利于蒙古栎嫩枝扦插生根。植物扦插不定根的发育及伸长与过氧化物酶(POD)、多酚氧化酶(PPO)和吲哚乙酸氧化酶(IAAO)相关氧化酶活性存在密切关系[20-22]。POD普遍存在于植物体内,既参与生物体的多种生理生化过程,又与光合作用、呼吸作用都有密切关联,有利于根原基的诱导[23]。PPO不仅促进细胞分裂、分化以及根原基形成和发育[22],而且参与愈伤组织的形成,对不定根发育起到重要的作用[24-25]。POD与PPO相辅相成,协同作用促进不定根根系的形成,而IAAO是分解IAA的专一性酶,过高的IAAO含量会抑制不定根根系的形成。本试验结果表明,在10 d内的愈伤组织诱导初期,POD活性呈上升趋势;在不定根伸长期,POD活性再次大幅度上升。POD活性在扦插生根关键期都出现了高峰[26-27],其作用的产物是促进不定根形成的辅助因子,参与根的诱导和表达,POD活性变化与马振华等[28]的研究结果相符。在植物生根过程中,酚类物质对不定根的形成和发育起着很重要的作用。在愈伤组织形成期PPO活性较高,说明此阶段PPO催化形成了大量“IAA-酚酸复合物”,从而促进愈伤组织的形成[27]。在愈伤组织膨大形成与不定根伸长的时期,处理组较对照组PPO活性大幅度增加,有利于根原基诱导和促进不定根伸长生长。处理组插穗IAAO活性明显比对照组低,主要是外源IBA进入植物组织后,迅速转化为IAA,抑制了IAAO活性升高,从而有利于不定根生成,40 d后不定根大量生成,IAA合成减弱,导致IAAO活性缓慢上升,但活性仍处于较低水平。
插穗生根过程中内源激素的含量变化表现出的规律性,与插穗形态变化相对应;外源激素IBA-K在一定程度上提高了IAA/ABA、IAA/ZR的值。扦插生根期间内源激素之间存在相互协同作用,较高的IAA/ABA值和较低的IAA/ZR值有利于蒙古栎嫩枝插穗不定根发育和伸长。多项研究表明,IAA对不定根的发生有着明显促进作用,不仅可以调节增强插穗基部细胞内相关酶的活性,加快呼吸速率,促进细胞分裂和分化,还可以促进mRNA的合成,影响差异蛋白质的表达,提高插穗能量代谢、胁迫抗性和信号传递等各方面的能力[29-31]。ABA对插穗生根具有明显的抑制作用,高浓度的ABA含量会抑制插穗生根[32]。有关于细胞分裂素ZR对扦插生根的作用研究各异,有些研究认为高浓度的ZR可以促进不定根的发生[33];也有一些研究认为,较低浓度细胞分裂素促进插穗生根[17]。在植物扦插生根的过程中,多种内源激素共同作用、相互制约[34]。有研究发现,IAA/ABA值越高越有利于不定根发育[18]。而高水平的IAA/ABA和IAA/ZR值有利于插穗根原基的诱导[35]。IAA/ABA值与生根呈正相关,通过外源处理保持较高的IAA/ABA值是不定根发育和生长的主要条件[32-33]。IAA/ZR值有着重要的调节功能,促进了愈伤组织的形成、不定根的产生和伸长生长[34,36]。本研究发现,IAA/ABA值在处理组始终高于对照组,且在生根关键期均出现峰值,而IAA/ZR值较高时有利于不定根原基的诱导,同时生根进程中IAA/ZR值较低时有利于不定根的发育和生长,具体IAA/ZR值如何调整更有利于插穗生根,还有待于进一步的研究。
猜你喜欢
作物研究(2022年1期)2022-11-27
热带作物学报(2022年7期)2022-08-06
作物学报(2022年10期)2022-07-21
舰船科学技术(2022年11期)2022-07-15
散文诗(2021年22期)2022-01-12
现代农业研究(2021年10期)2021-11-02
中国蔬菜(2021年5期)2021-06-15
意林(绘英语)(2018年2期)2018-06-26
读写算·小学低年级(2017年7期)2017-08-11
山东青年(2016年1期)2016-02-28
