
山西灵空山典型天然林空间结构特征及其对生产力的影响
2022-02-10 12:35刘铭波韩海荣程小琴蔡锰柯刘慧敏
西北林学院学报 2022年1期
刘铭波,韩海荣,程小琴,蔡锰柯,刘慧敏
(北京林业大学 生态与自然保护学院,北京 100083)
天然林在无人为干扰情况下形成的复杂林分结构能代表该地区植物的最优配置[1],是维持生态系统功能与稳定性的重要因子,不仅反映了个体间竞争状态、分布格局及物种多样性[2],还影响着林分更新、发展的方向[3]。通过研究森林生态系统林分空间结构,揭示影响生产力变化的主要空间结构因子,有助于探究林分空间结构潜在作用机制,为“近自然”理念下的生产实践提供理论依据和研究基础[4]。
目前已有大量研究探讨林分结构多样性[5]与生产力的相关关系,如吴兆飞等[6]选取物种与胸径的丰富度、香农指数、辛普森指数来反映物种与结构多样性,以此探究东北温带森林生产力驱动因子;谭凌照等[7]选取胸径香农指数、均匀度、变异系数、基尼系数等指标表征群落结构变异,从而探讨其与生产力的关系。然而这些研究多采用与距离无关的林分非空间结构指标,难以体现林木空间分布状态。近年来许多学者提出了更为系统地描述林分空间特征的方法[8]。其中4株相邻木法[9]应用最为广泛,不仅可操作性强[10],还能充分反映参照树与周围树木的关系,精确地描述林分空间特征。但现有研究多针对结构相对简单的人工林,且侧重于林分空间结构描述[11]、人工林空间结构优化模拟[12]和结构参数综合评价[13]等方面,空间结构对天然林生产力驱动机制的研究较少。本研究基于该方法,结合表征林分垂直结构多样性的林层指数(S)[14],综合探讨二者相关关系。
辽东栎(Quercuswutaishanica)是我国暖温带落叶阔叶林的主要优势种和建群种之一,常生于华北东部地区的阳坡和半阳坡,具有抗干旱、耐瘠薄的特性[15];油松(Pinustabuliformis)是我国暖温带森林特有的优势树种,主要生长于北方地区[16]。灵空山国家级自然保护区地处华北暖温带落叶阔叶林和温带草原的过渡地带,生态环境优良且受人为干扰较少,具有较高研究价值。现有关于辽东栎林和油松林研究多集中在生物多样性[17]、间伐对土壤酶活性的影响[18],以及土壤理化性质对林分生产力影响等方面[19],关于林分空间结构因子对天然林生产力的影响机制鲜见报道。因此,本研究选取该地典型辽东栎林、油松林为对象,通过分析天然林空间结构特征,探究空间结构对乔木层生产力变化的驱动机制,以期对灵空山地区天然林保护、管理提供科学依据。
1 材料与方法
1.1 研究区概况
研究区位于山西灵空山自然保护区,地处太岳山系中部(36°39′N,112°5′-112°6′E),属于典型暖温带大陆性季风气候,年均温8 ℃,无霜期125 d,年降水量600~650 mm,集中于7-9月。地势北高南低,平均海拔1 600 m,土壤类型以褐土为主[15]。该地地带性植被有暖温带针叶林、落叶阔叶林和针阔混交林,主要乔木树种有辽东栎、油松、白桦(Betulaplatyphylla)、核桃楸(Juglansmandshurica)、山杨(Populusdavidiana)等;主要灌木有胡枝子(Lespedezabicolor)、黄刺玫(Rosaxanthina)、土庄绣线菊(Spiraeapubescens)等;草本主要有细叶苔草(Carexrigescens)、糙苏(Phlomisumbrosa)、山罗花(Melampyrumroseum)等。
1.2 样地设置与调查
2019年7-8月在灵空山自然保护区采用典型抽样法选取立地条件基本相似,林相整齐,具有代表性的辽东栎林和油松林,设置样地大小为20 m×30 m,各9块,调查时将其划分为24个5 m×5 m的小样方,以便于每木检尺和树木准确定位。样地的调查因子主要包括树种名称、胸径(DBH≥2 cm)、树高、树木坐标及海拔、坡度等基本样地信息(表1)。同时,在样地内以4 cm为1个径级,每个径级选取1~3株作为标准木,在胸径(1.3 m)高处向东西和南北2个方向垂直钻取2根树芯(钻至髓心处),取出后及时放入吸管中保存,编号并记录相应信息,每块样地约20棵树,40根树芯。样本采集后带回实验室,固定于定制木槽中于通风处阴干,并依次使用200、400、600目砂纸打磨,直至年轮清晰。
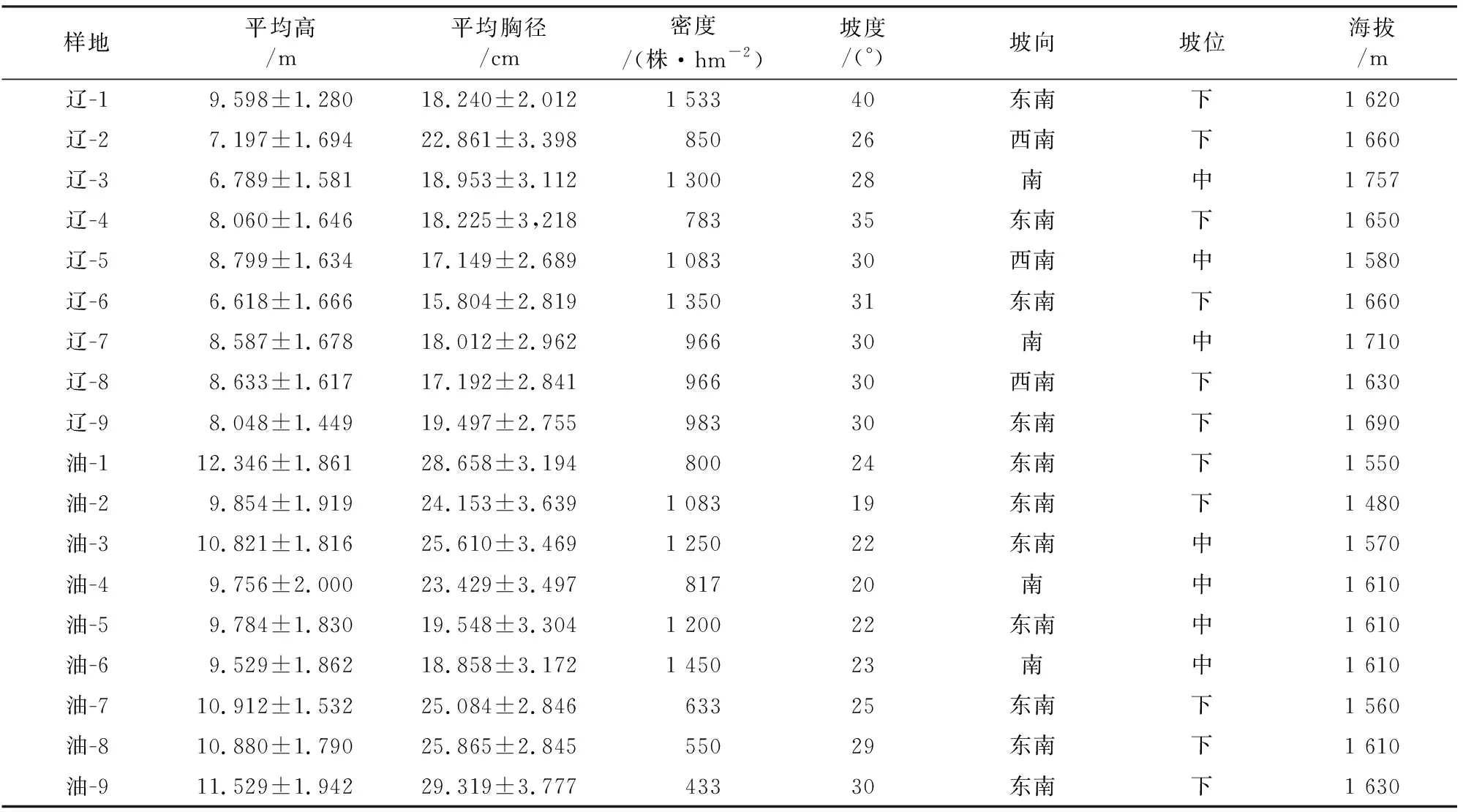
表1 样地基本信息Table 1 Basic information of sample plots
1.3 林分空间结构参数计算
基于相邻木关系的林分空间结构描述方法为群落结构的表达提供了科学依据。林分中任意一株单木(参照树)和其周围的4株最近相邻木构成基本结构单元[9]。结构参数主要有4个,W用于描述树木个体水平分布格局;M用于描述树种空间隔离程度;U用于描述参照树与相邻木胸径的相对大小状态,即相对优劣状态;C用于描述林木冠层拥挤程度。本研究增加了描述林分垂直结构多样性的林层指数(S)[14]。
1)角尺度(W)[9]指任意2株最近相邻木与参照树所夹小角α小于标准角α0(α0=72°)的个数占所考察的4个α角的比例:
(1)
(2)
W取值依次是0、0.25、0.50、0.75、1.00,分别对应绝对均匀、均匀、随机、不均匀、团状分布5种生物学意义。具体研究实践表明,分布格局也可划分为以下3种类型:当W<0.457时为均匀分布,当0.457≤W≤0.517时为随机分布,当W>0.517时,为聚集分布。
2)大小比数(U)[9]指胸径大于参照树的相邻木个数占所考察的4株相邻木的比例:
(3)
(4)
U取值同上,分别对应优势、亚优势、中庸、劣态、绝对劣态,明确定义了参照树在其结构单元中所处的生态位即竞争能力,生物意义明显。
3)混交度(M)[9]指参照树i的4株最近相邻木与参照树不属同种的个体所占的比例:
(5)
(6)
M取值同上,分别对应零度混交、弱度混交、中度混交、强度混交、极强度混交5种生物学意义,表示参照树在该结构单元的隔离程度。
4)密集度(C)[20]指林木冠层拥挤程度:
(7)
(8)
C取值同上,分别对应很稀疏、稀疏、中等密集、比较密集、十分密集,表示参照树与其4个最近相邻木的竞争拥挤程度,还反映了森林冠层是否连续覆盖林地。
5)林层指数(S)[14]描述林分垂直结构多样性,用公式表示为:
(9)
zi为参照树i的空间结构单元内林层数;sij为离散性变量,林层指数越接近1,表明林分在垂直方向上成层性越复杂。
1.4 乔木层生产力计算
通过Lintab6树木年轮分析仪测量年轮宽度,精确至0.01 mm。根据各树种异速生长模型计算历年乔木整株生物量,相邻2 a生物量之差即为该样地林分生产力。各树种异速生长模型及生产力计算公式如下。
油松生物量[21]:
V=2.905D0.549e0.097D
(10)
辽东栎生物量[21]:
V=0.187(D2H)0.833
(11)
山杨生物量[21]:
V=0.085D2.480
(12)
生产力:
N=Va-Va-1
(13)
式中:V为标准木乔木层生物量;D为标准木胸径;H为标准木树高;a为年份;N为该样地净生产力。
1.5 数据处理与分析
采用林分空间结构软件(Winkelmass 1.0)计算每株林木的空间结构参数W、U、M;采用Excel 2003计算结构参数C、S,并统计林木空间结构相对频率值;使用Canoco 5.0软件进行冗余分析并绘图,探讨林分空间结构参数对林分生产力的影响。
2 结果与分析
2.1 2种林分空间结构特征
由表2可知,2种天然林角尺度频率值均以随机分布轴(W=0.50)呈正态分布,说明2种林分类型林木多呈随机分布状态;各分布等级间大小比数频率值均无显著性差异(P>0.05),说明2种林分类型林木胸径大小分化程度较低;当分布等级为0和0.25时,2种林分混交度频率值显著大于其他分布等级(P<0.05),说明2种天然林均以零、弱度混交状态为主;分布等级为1时,2种天然林密集度频率值均显著大于其他等级(P<0.05),说明2种林分类型冠层分布十分密集。分布等级为0时,辽东栎林角尺度频率值显著大于油松林(P<0.05),说明呈均匀分布状态的辽东栎更多;相同分布等级下,2种林分间大小比数频率值无显著性差异(P>0.05),说明2种林分类型的林木胸径大小分化程度相近;分布等级为0.25时,辽东栎林混交度频率值显著大于油松林(P<0.05),说明弱度混交状态辽东栎更多;分布等级为0.50和0.75时,辽东栎林密集度频率值显著大于油松林(P<0.05),说明在林冠重叠程度较高时辽东栎林木分布更多。
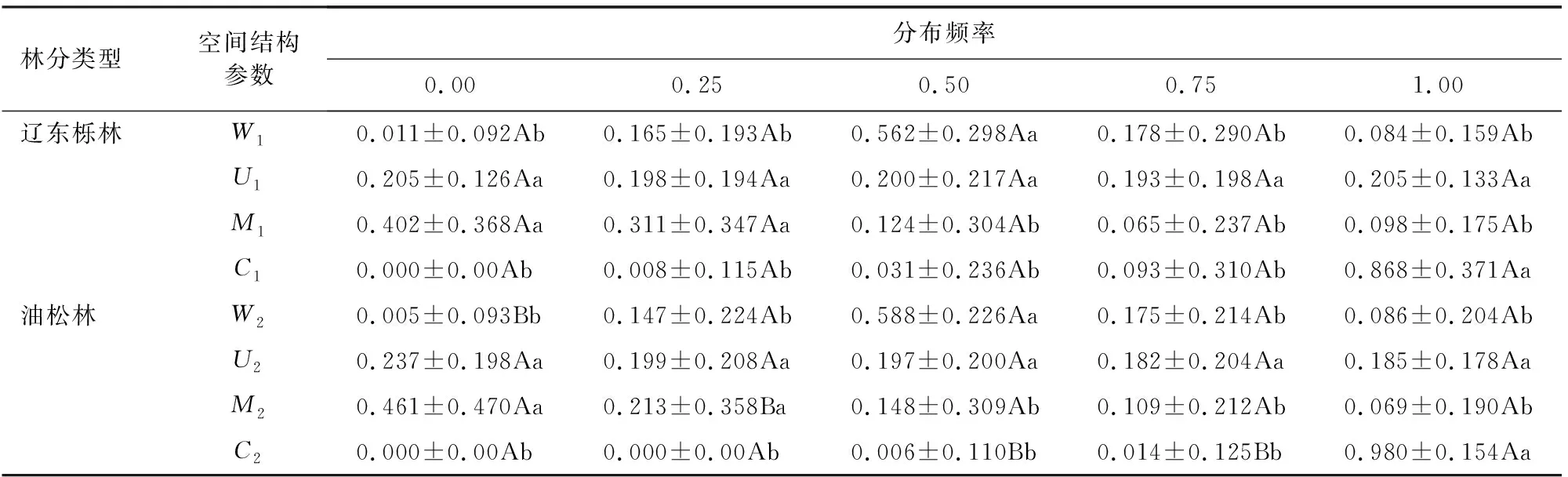
表2 各分布等级下辽东栎、油松林空间结构参数分布频率Table 2 Distribution frequency of spatial structure parameters of Q.wutaishanica and P.tabuliformis forests in different distribution levels
综合各分布等级来看(表3),2种林分整体水平格局均呈现聚集分布(W>0.517);辽东栎林大小比数为0.50~0.75,说明辽东栎林整体处轻微劣势状态,油松林大小比数为0.25~0.50,说明油松林整体处于中庸状态;2种天然林混交度均为0.25~0.50,林分均呈现中度混交状态,树种多样性较低;2种天然林密集度均大于0.95,林冠层整体处于十分拥挤状态;林层指数相差不大,且油松林垂直结构略优于辽东栎林。

表3 辽东栎、油松林空间结构参数特征Table 3 Average value of spatial structure parameters of Q.wutaishanica and P.tabuliformis forests
2.2 2种林分生产力特征
由表4可知,辽东栎林蓄积量为101.117~215.906 t·hm-2,林分生产力为1.845~5.808 t·hm-2·a-1;油松林蓄积量为57.430~135.540 t·hm-2,林分生产力为1.670~5.126 t·hm-2·a-1。辽东栎林蓄积量、生产力水平显著大于油松林(P<0.05),说明辽东栎林有机物积累能力更强,乔木层资源更丰富。
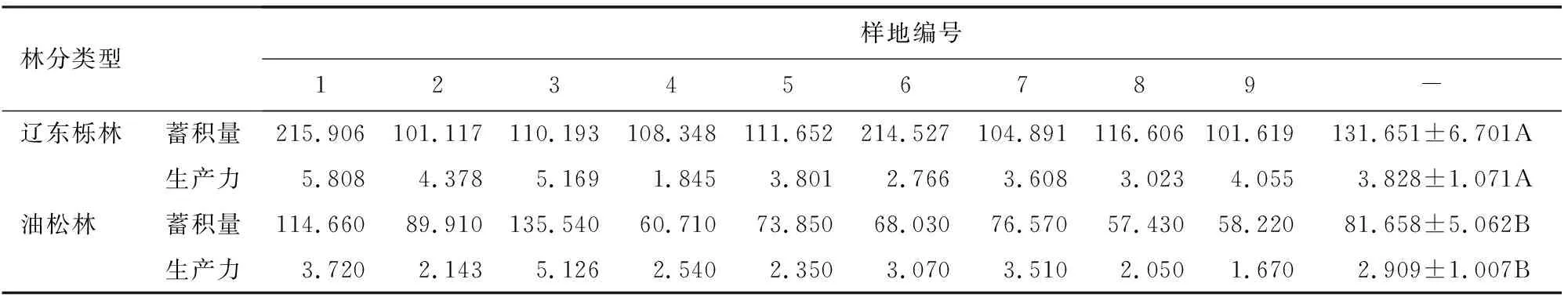
表4 辽东栎、油松林蓄积量、生产力特征Table 4 Stock volume and productivity characteristics of Q.wutaishanica and P.tabuliformis forests
2.3 林分空间结构与生产力的关系
对2种林分类型各9个样地生产力大小进行冗余分析,以林分生产力、蓄积量为响应变量,以5个空间结构参数作为解释变量,探究其相关关系。由图1可知,第1轴和第2轴共解释了生产力变异程度的76.19%,能很好地展现辽东栎林生产力与林分空间结构因子之间的关系。辽东栎林蓄积量(V1)和C1、M1、S1、U1呈正相关,与W1呈负相关。林分生产力(N1)与U1呈正相关,与W1、C1、M1、S1呈负相关。其中U1与第1轴呈极显著正相关(P<0.01),解释了林分生产力变异的50.7%,说明U1是辽东栎林林分生产力变化的主要驱动因子。
由图2可知,第1轴和第2轴共解释生产力变异程度的82.66%,能很好地展现油松林生产力与林分空间结构因子之间的关系。蓄积量(V2)和生产力(N2)均与U2、C2、M2呈正相关,与W2、S2呈负相关。其中W2与第1轴呈显著负相关(P<0.05),解释了生产力与蓄积量变异的57.1%,说明W2是油松林林分生产力变化的主要驱动因子。
3 结论与讨论
3.1 结论
研究发现,山西灵空山自然保护区辽东栎林、油松林总体呈聚集分布,树种多样性较低且冠层分布十分密集,林木大小分化不明显,垂直结构较为简单。影响辽东栎林生产力变化的主要空间结构因子是大小比数,油松林生产力主要空间结构驱动因子则是角尺度。因此,该保护区经营管理时,辽东栎林应首先关注林木间胸径大小差异程度,油松林则应侧重其水平分布格局。林分空间结构能有效解释林分生产力变化,必要时可以依据空间结构参数对林分进行优化,从而更好地发挥天然林的生态价值。
3.2 讨论
3.2.1 辽东栎、油松林林分空间结构特征 研究结果表明,2种林分林木均呈现聚集分布格局。从单木角度来看,随机分布的结构单元数显著多于其他类型(P<0.05),并且在绝对均匀和团状分布时角尺度相对频率值几乎为0,符合林木自疏后呈随机分布状态这一生态学规律,与郝江勃等[22]在黄土高原地区的研究结果一致。本研究区内辽东栎林处于轻微劣势状态,而油松林处于中庸状态,分布等级越低,参照树的竞争压力越小,即辽东栎林竞争更激烈,结构单元多处于劣势。这可能是由于油松林大径级林木更多,其结构单元更具竞争优势[23]。2种林分零、弱度混交结构单元均占据较大比例,非同种个体抢占空余生态位机会偏小,群落演替尚处于较低阶段。本研究区内林分密度较大,资源和空间有限,制约了林下植被的更新,这与伊力塔等[15]、马映栋等[24]观点一致。2种林分密集度相近,且林冠重叠度均较高,这种冠层结构造成林下环境时空异质性较差,致使光照条件等资源利用受限[25]。2种林分垂直结构也较为简单,林层指数越小,其群落结构稳定性越差。这可能是由于该区域林分混交度低,导致树高差异不明显。
3.2.2 辽东栎、油松林空间结构特征对生产力的影响 对辽东栎林分析发现,大小比数与林分生产力呈显著正相关(P<0.05),即大小比数越大,处于劣势的结构单元数量越多,林分生产力越高。辽东栎林中小径级树木较多,易处于劣势地位,竞争压力较大,但是处于发育早期的小径级林木生长速度较快,更易积累林分有机物质[26]。仇建习等[27]探究天目山近自然毛竹林空间结构与胸径关系所得结论与本研究相似。郭梦昭等[28]也得出了生产力与径级结构多样性呈负相关的结论。此外,密集度、混交度、角尺度和林层指数均与辽东栎林生产力呈负相关关系。密集度越大,林分趋于郁闭,穿透雨量低从而阻碍降雨再分配过程,使有机物积累缓慢。随混交度的增加,整体群落结构趋于完善,林下灌草同乔木层竞争环境要素,致使可利用资源和空间减少[29],种间竞争越加激烈。这与巫志龙等[30]认为中度混交降低了林分稳定性及M.Viletal[31]对木材生产力的研究结果一致。
对油松林分析发现,角尺度与林分生产力呈显著负相关(P<0.05)。林分角尺度越大,林木水平格局越聚集,加剧了有限空间内乔木层的资源竞争[32],造成林木胸径增长缓慢。此外,林木聚集分布会导致林分透光条件变差,抑制灌草层更新,整体群落结构变差。这与陈昊泓等[33]对湖南栎类天然次生林的研究结论一致。油松林生产力与密集度呈正相关,与辽东栎林结果相反。综合2种林分生物与区域特性考虑可能是由于:1)油松作为喜阳针叶林树种,同密集度水平的林下光照条件优于辽东栎林。2)研究区油松林林下枯落物层较辽东栎林薄,物质循环过程存在差异。3)所测研究区各样点油松林生产力均<6 t·hm-2·a-1,低于山西省平均水平(7.4 t·hm-2·a-1)[34],可能会对研究结果产生影响。
林分生产力大小不仅受光照、水分、土壤养分等环境条件影响,也会因林分空间结构所处微环境的改变而影响树木生长状态和有机物积累。山西灵空山辽东栎林、油松林生产力主要驱动因子不同,这归结于2种林分的树种生物学特性迥异与研究区域地理环境特殊性,从而造成空间结构与生产力响应机制不同。研究林分空间结构不能局限于描述林分状态,更要侧重于调整结构、提高林分生产力及生态系统服务功能等问题上[35]。当被调查林分结构并非理论上最优值时,可以通过改善小型结构单元来实现空间结构优化[36]。
猜你喜欢
辽宁省博物馆馆刊(2020年0期)2020-08-13
防护林科技(2020年6期)2020-08-12
诗林(2019年6期)2019-11-14
东坡赤壁诗词(2019年3期)2019-07-05
绿色科技(2019年6期)2019-04-12
绿色科技(2019年6期)2019-04-12
诗潮(2018年10期)2018-10-17
环球人文地理·评论版(2016年8期)2017-01-19
天津农业科学(2016年12期)2017-01-11
文艺生活·中旬刊(2016年11期)2016-12-13
