
miR-4795-3p通过靶向EGFR抑制胃癌细胞的增殖和侵袭
2022-02-10 07:26兰国玉汤守元黄耿朱钟钟李新明罗海平姜进平
国际医药卫生导报 2022年1期
兰国玉 汤守元 黄耿 朱钟钟 李新明 罗海平 姜进平
1鄂东医疗集团黄石市中心医院湖北理工学院附属医院胃肠外科,黄石 435000;2鄂东医疗集团黄石市中心医院湖北理工学院附属医院泌尿外科,黄石 435000
胃癌是全球最常见的恶性肿瘤之一,是癌症相关死亡的重要原因[1]。胃癌缺乏早期诊断和治疗的标志物,许多患者确诊时已处于晚期,导致胃癌患者的5年总生存率很低[2]。探究新的分子靶点对胃癌的诊疗具有重要意义。微小RNA(miRNA)是一类非蛋白编码的单链小RNA,长度为19~25个核苷酸,可在转录后调控基因的表达[3]。miRNA通过影响细胞的代谢、分化、增殖、侵袭等生物学过程,在胃癌的发生、发展过程中具有重要功能[4]。miR-4795-3p长度为22个核苷酸,其在疾病中的作用尚未见报道。本研究于2020年6月至12月探究miR-4795-3p对胃癌细胞增殖和侵袭能力的影响及可能的调控机制。
材料与方法
1、细胞株与主要试剂
胃癌细胞株BGC823购于中国科学院典型培养物保藏委员会细胞库;RPMI-1640培养基购于美国Gibco公司;淋巴细胞增殖检测(MTS)试剂盒购于上海生工生物工程股份有限公司;阴性对照模拟物、miR-4795-3p模拟物、表皮生长因子受体(epidermal growth factor receptor,EGFR)野生型(EGFR-WT)重组质粒、EGFR突变型(EGFR-MT)重组质粒购于上海碧云天生物技术有限公司;双荧光素酶报告基因检测试剂盒购于美国Promega公司;Lipofectamine 3000转染试剂、实时定量聚合酶链反应(qRT-PCR)试剂盒购于美国Invitrogen公司;超敏ECL发光试剂盒购于美国Thermo公司;一抗和辣根过氧化物酶标志的二抗购于美国BD公司。
2、细胞培养和转染
BGC823采用含10%胎牛血清RPMI-1640培养基培养于37℃、5%CO2培养箱中,在6孔板接种对数生长期BGC823细胞,采用Lipofectamine 3000试剂转染阴性对照模拟物或miR-4795-3p模拟物,标记为阴性对照组和miR-4795-3p组,12 h后更换培养基。
3、qRT-PCR检测miR-4795-3p、EGFRmRNA表达情况
采用TRIzol法提取两组对数生长期细胞的总RNA,逆转录为cDNA后进行qRT-PCR反应,以U6为miR-4795-3p的内参基因,以GAPDH为EGFR的内参基因,根据2-ΔΔCt法计算miR-4795-3p、EGFR mRNA表达情况。EGFR正向引物:5’-AGGCACGAGTAACAAGCTCAC-3’,反 向 引 物5’-ATGAGGACATAACCAGCCACC-3’;U6正 向 引 物:5’-TGACACGCAAATTCGTGAAGCGTTC-3’,反 向 引 物5’-CCAGTCTCAGGGTCCGAGGTATTC-3’;miR-4795-3p正向引物:5’-CAGAAGTGGCTAATAATA-3’,反向引物5’-CTTCATCAGCGTCAACAG-3’;GAPDH正 向 引 物:5’-GGAGCGAGATCCCTCCAAAAT-3’,反 向 引 物5’-GGCTGTTGTCATACTTCTCATGG-3’。
4、MTS法检测BGC823细胞增殖情况
调整两组BGC823细胞密度为6×103个/ml,以200μl/孔接种至96孔板,于第1、2、3、4天检测BGC823细胞的增殖。加入20μl/孔MTS试剂,于培养箱孵育4 h,酶标仪检测每孔于450 nm波长处吸光度(A450)值,绘制BGC823细胞生长曲线。
5、Transwell实验检测BGC823细胞侵袭能力
用不含血清的RPMI-1640培养基调整两组BGC823细胞密度为1×105个/ml,以200μl/孔接种于Transwell上室,在下室加入600μl/孔含血清的培养基,培养24 h后,甲醇固定并采用结晶紫染色,于倒置显微镜下计数细胞侵袭数。
6、生物信息学软件预测和双荧光素酶报告基因实验验证miR-4795-3p的靶基因
采用miRcode数据库预测miR-4795-3p的靶基因,EGFR可能是miR-4795-3p的靶基因。分别将EGFR-WT质粒或EGFR-MT质粒与阴性对照模拟物或miR-4795-3p模拟物共转染至BGC823细胞,48 h后采用双荧光素酶报告基因检测试剂盒检测每孔BGC823细胞荧光素酶的相对活性。
7、蛋白质免疫印迹试验(Western blot)检测靶基因蛋白的表达
用RIPA裂解液裂解BGC823细胞并提取总蛋白,采用SDS-PAGE凝胶电泳分离蛋白,硝酸纤维素膜转膜。室温封闭后加入一抗,4℃孵育过夜。加入二抗,室温孵育1 h,滴加ECL显影液,采用凝胶成像仪曝光,观察每组蛋白条带。
8、统计学分析
采用SPSS 21.0软件分析数据,符合正态分布的计量资料均以均数±标准差(±s)表示,两组间比较应用独立样本t检验,以P<0.05为差异有统计学意义。
结 果
1、各组BGC823细胞中miR-4795-3p的表达情况
阴性对照组和miR-4795-3p组BGC823细胞中miR-4795-3p的表达分别为(1.02±0.11)和(11.04±1.23),miR-4795-3p组是阴性对照组的10.82倍,差异有统计学意义(t=8.14,P<0.01)。
2、高表达miR-4795-3p对BGC823细胞增殖能力的影响
与阴性对照组相比,miR-4795-3p组BGC823细胞在第2、3、4天的A450值显著下降(均P<0.05),表明miR-4795-3p能够抑制胃癌BGC823细胞增殖能力(图1)。
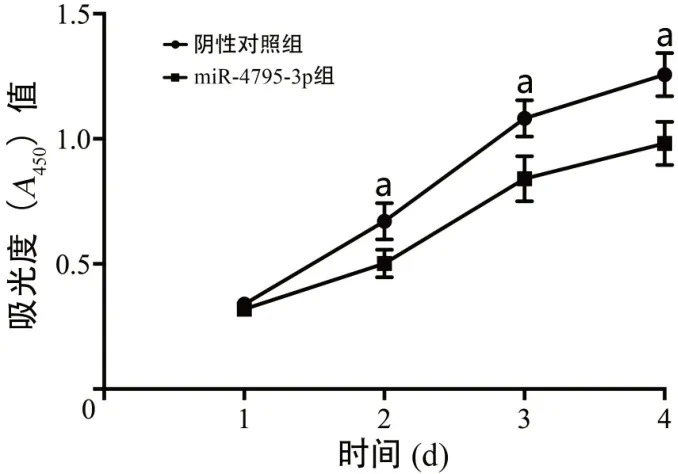
图1 高表达miR-4795-3p对BGC823细胞增殖能力的影响
3、高表达miR-4795-3p对BGC823细胞侵袭能力的影响
阴性对照组和miR-4795-3p组BGC823细胞细胞侵袭数分别为(79.76±6.27)个和(31.38±5.46)个,差异有统计学意 义(t=5.82,P<0.01),提示miR-4795-3p可抑制胃癌BGC823细胞的侵袭能力。
4、双荧光素酶报告基因实验验证miR-4795-3p的靶基因
双荧光素酶报告基因实验显示,阴性对照+EGFR-WT组和miR-4795-3p+EGFR-WT组BGC823细胞相对荧光素酶活性分别为(1.01±0.08)和(0.22±0.05),差异有统计学意义(t=8.58,P<0.01),提示miR-4795-3p可靶向结合EGFR。
5、高表达miR-4795-3p对EGFRmRNA表达水平的影响
阴性对照组和miR-4795-3p组BGC823细胞中EGFR mRNA的表达分别为(1.00±0.03)和(0.35±0.04),证实miR-4795-3p可直接下调EGFR mRNA的表达(t=14.08,P<0.01)。
6、高表达miR-4795-3p对EGFR蛋白的表达
Western blot结果表明,与阴性对照组相比,miR-4795-3p组BGC823细胞EGFR蛋白减少,EGFR/MAPK信号通路蛋白如p-MEK、p-ERK表达降低(图2)。

图2 高表达miR-4795-3p对EGFR蛋白表达的影响
讨 论
研究表明,miRNA由具有发夹结构的miRNA前体通过Dicer酶加工合成,其表达具有明显的细胞特异性[5]。miRNA通过表现为癌基因或抑癌基因作用,其上调或下调与胃癌的发生和发展密切相关[6]。miRNA如miR-665[5]、miR-129-5p[7]等通过负性调控癌基因的表达,抑制胃癌的生长和转移。miRNA如miR-301a-3p[8]、miR-223-5p[9]等通过下调抑癌基因的表达,促进胃癌的生长和转移。miR-4795-3p在胃癌细胞中的作用并不清楚。本研究表明,高表达miR-4795-3p可显著抑制胃癌BGC823细胞的增殖和侵袭能力,miR-4795-3p在胃癌细胞中发挥抑癌基因作用。
miRNA与靶基因mRNA的3’非翻译区互补配对,诱导靶基因mRNA的降解或抑制蛋白合成,进而在转录后水平负性调控靶基因的表达[6]。本研究应用生物信息学软件miRcode数据库预测显示,miR-4795-3p和EGFR mRNA存在互补配对的反应元件。EGFR蛋白是一种糖蛋白,其在多种实体瘤中过表达,可促进肿瘤细胞的增殖、侵袭及血管生成[10]。高表达的EGFR与胃癌的淋巴结转移、临床分期呈正相关,是食胃癌患者的独立预后因素,EGFR参与胃癌的发生、发展过程[11]。双荧光素酶报告基因实验显示,miR-4795-3p可互补配对结合EGFR mRNA。本研究进一步证实,转染miR-4795-3p后细胞中EGFR表达下降,表明miR-4795-3p可靶向负调控EGFR的表达,EGFR是miR-4795-3p的靶基因。EGFR/MAPK信号通路的激活可促进胃癌细胞的增殖和侵袭[12]。本研究显示,miR-4795-3p下调EGFR表达后,BGC823细胞中EGFR/MAPK信号通路蛋白如p-MEK、p-ERK表达降低,EGFR/MAPK信号通路被抑制。
综上所述,miR-4795-3p能够通过靶向负调控EGFR表达,干扰EGFR/MAPK信号通路,抑制胃癌细胞的增殖和侵袭能力,本研究为miRNA在胃癌的靶向治疗研究提供了理论基础。
利益冲突 所有作者均声明不存在利益冲突
猜你喜欢
安徽农业科学(2022年9期)2022-05-17
保健与生活(2020年6期)2020-03-20
江苏农业科学(2019年23期)2019-03-03
恋爱婚姻家庭·养生版(2019年1期)2019-01-25
中国卫生产业(2018年12期)2018-05-14
中国中医药信息杂志(2018年11期)2018-01-05
三联生活周刊(2017年1期)2017-01-11
医学信息(2016年29期)2016-11-28
