
北京植被净初级生产力对物候变化的响应
2022-02-07 01:04魏晓帅高永龙范雅倩张德怀李鑫豪刘新月徐铭泽查天山
农业工程学报 2022年18期
魏晓帅,高永龙,范雅倩,林 岭,毛 军,张德怀,李鑫豪,刘新月,徐铭泽,田 赟,刘 鹏,贾 昕,查天山
北京植被净初级生产力对物候变化的响应
魏晓帅1,2,高永龙3,范雅倩4,林 岭3,毛 军1,2,张德怀5,李鑫豪1,2,刘新月1,2,徐铭泽1,2,田 赟1,2,刘 鹏1,2,贾 昕1,2,查天山1,2※
(1. 北京林业大学水土保持学院,北京 100083;2. 北京林业大学水土保持国家林业局重点实验室,北京 100083;3. 北京市园林绿化规划和资源监测中心,北京 100013;4. 北京松山国家级自然保护区管理处,北京 102100;5. 密云区园林绿化局,北京 101599)
植被净初级生产力(Net Primary Productivity,NPP)对物候的响应是全球气候变化背景下的重要研究内容,气候变化对植物物候与NPP的影响仍需明了,物候的时空变异规律更需深入探讨。该研究基于2001—2020年MODIS归一化植被指数(Normalized Difference Vegetation Index,NDVI)数据提取北京植被物候信息,利用CASA(Carnegie-Ames-Stanford-Approach)模型模拟NPP,通过线性回归、趋势分析和结构方程模型等方法,阐明NPP与物候时空变化特征,探究气象因素和物候变化对NPP的影响。结果显示:1)2001—2020年超过70 %的区域出现生长季开始(Start of Growing Season,SOS)逐渐提前,平均每年提前0.57 d。超过90%的区域生长结束期(End of Growing Season,EOS)逐渐推迟,平均每年推迟0.51 d。2)SOS提前和生长季(Length of Growing Season,LOS)延长均对NPP增长产生显著影响(0.05)。SOS每提前1 d,NPP增长3.74 g/m2;LOS每延长1 d,NPP增长2.65 g/m2。秋季EOS推迟对NPP变化影响不显著。3)春季和秋季,气候通过改变物候(SOS、EOS)对NPP的间接影响大于直接影响,而夏季温度和降雨对NPP的直接影响更大。该研究表明春季物候变化是NPP年际变异的重要驱动因子,春季物候提前导致NPP年总量增加。研究结果是都市区植被生产力如何响应气候变化认识的重要补充。
遥感;净初级生产力;植被物候;气候变化;NDVI
0 引 言
碳循环是生物圈中基本的生物地球化学过程,其作为陆地生态系统的关键过程,已成为生态学及相关学科的研究热点[1]。气候变暖会造成植被物候的变化[2-3],植被物候受气候因素调节,影响生态系统功能和生物过程,改变相关的生理和生物地球化学过程,对陆地生态系统的碳循环有重要影响[4]。
净初级生产力(Net Primary Productivity,NPP)是碳循环的主要组成部分[5],是生态系统固定碳能力和判定生态系统对气候变化适应性的重要指标[6]。植物生长季开始(Start of Growing Season,SOS)、生长季结束(End of Growing Season,EOS)和生长季长度(Length of Growing Season,LOS)是典型物候期,已经被认为是影响生态系统功能等许多生物过程的关键指标[7]。物候对气候变化非常敏感,全球温度升高可能会导致物候提前[2,8],并且植物光合作用在不同季节对气候变化的响应不同[9]。春季和秋季物候的变化可以不同程度地改变生长季节长度,并影响陆地生物圈与大气间的碳循环。因此,阐明物候变化对生态系统生产力的影响及其对气候的反馈,探究物候变化、气象因素和生产力间的相互关系变得尤为重要。
传统物候观测存在观测成本高、时间序列短、观测方法不统一等问题。归一化植被指数(Normalized Difference Vegetation Index,NDVI)可以有效捕捉植被冠层的绿度信息,通过遥感监测数据反演物候信息使长时间大区域植被物候的研究成为了可能,已经广泛用于植被物候方面的研究[9-10]。季前降水可能增加春季物候开始期的水量需求从而间接影响植被春季生长季开始期的变化[11],也有研究认为生长季的提前是由全球气候变暖导致的[2-3,12],气候变化背景下,人们普遍认为会出现植被生长季开始时间提前和结束时间推迟,从而增加生长季的长度,因此初级生产力的增加是因为有更多的时间用于碳同化和生物量增长[13]。但也有研究发现,早春可能会增加蒸腾速率,导致夏季和秋季水分胁迫,降低生产力[14-15];并且生长季的延长可能会增加秋季生态系统的呼吸作用时间,导致碳收益被抵消[8,16],因此虽然秋季变暖倾向于延长生态系统的生长季节,但季末呼吸损失了早春的固碳量,升温和早春也可能通过间接或滞后效应影响生长季节后期的生态系统功能[17]。然而,物候和气候因素对NPP的相对重要性仍然没有得到有效的解答。
由于生态系统的复杂性,物候和气候因素对净初级生产力的影响仍然存在着很大的不确定性,生长季延长对年际NPP的影响仍不明确。在生态系统中,NPP受到植被物候、降水及温度等环境因素的共同影响,气象和物候指标(SOS、EOS)在NPP年际变化中的贡献度并没有得到有效量化。在气候变化和城市热岛效应的影响下城市植被更容易受到环境变化的影响,对气候敏感性更高[18]。在气温上升和气候变化加剧的背景下,了解NPP与物候之间的关系有助于改善人们对城市区域植被碳汇的预测。北京作为中国首都,并且拥有丰富的森林资源,在气候变化和实现“碳中和”背景下已经受到了国际和社会各界的广泛关注[19]。为了解决以上存在的问题,本研究以北京为研究区域,探讨物候变化在NPP时空变化中的作用及其潜在驱动力,阐明NPP和物候(SOS、EOS和LOS)的时空变化特征,探究物候及NPP的相关性并且量化物候和气象因素对NPP的影响。
1 材料与方法
1.1 研究区域
研究区位于中国北京,其气候属于温带半湿润大陆性季风气候,具有明显的季节分异性,对于植被物候相关方面的研究有很强的代表性。范围39°26′N~41°03′N,115°25′E~117°29′E,位于华北平原北部,毗邻渤海湾。北京平原海拔20~60 m,山地海拔1000~2000 m。北京森林资源丰富,森林面积58.81万hm2(其中有林地面积58.73万hm2),森林覆盖率35.84%。年平均气温为9.7°C,年降水量为424.2 mm。降水的季节分布极不均匀,年降水量的80 %集中在夏季的6—8月。主要乔木类型为暖温带落叶阔叶林和暖温带针叶林。有栓皮栎()、油松()、侧柏()、栎类()和桦木科()等,山顶1 800~2 000 m以上生长杂草草甸。
为了排除土地利用和人为因素的干扰,本研究利用GlobeLand30数据集对北京2000年、2010年和2020年土地利用情况进行分析,由于耕地、水体、人造地表和裸地受人为干扰影响严重,所以只针对林地、草地、和灌木地进行研究,并选择从2000—2020年土地利用未变化的地区进行研究(图1)。
1.2 数据来源
1.2.1 MODIS NDVI 数据
本研究选取NASA 提供的MODIS MOD13A1数据集进行植被相关信息提取(https://ladsweb.modaps. eosdis.nasa.gov/search/),空间分辨率为500 m,采集时间范围为2001—2020年,时间分辨率为16 d。MOD13A1数据充分考虑了高覆盖度、低视角、云影、气溶胶等因素的影响,被广泛应用于不同尺度植被的动态监测和研究,为了剔除裸土和水体对NDVI的影响,研究只考虑全年NDVI均值大于0.1、最大NDVI大于0.15的栅格[19-20]。
1.2.2 气象和土地利用数据
气象数据来自中国气象数据网(http://dataxma.cn/),为了保证空间插值结果更加精确本研究选取了北京及其周边19个气象站点,获取了2001—2020年月平均气温、月降水量和日照时数,由于太阳辐射(Solar Radiation,SOL)监测站较少,为了获得更精确的地表辐射,采用纬度和日照时数的经验公式,得到最终的太阳辐射[21]。利用ArcMap软件采用克里金(Kriging)方法进行空间插值,分辨率为500 m。对插值结果进行投影变换,切割提取研究区域,形成栅格数据集。为了保证气象数据的可用性,本研究同时获取了中国青藏高原科学数据中心(Tibet Plateau Scientific Data Center,TPDC)2001—2020年空间分辨率为1 000 m的月气温、月降水量和辐射数据集。土地利用数据采用中国开发的30 m空间分辨率全球土地覆盖数据GlobeLand30数据集,经过重采样到500 m分辨率。
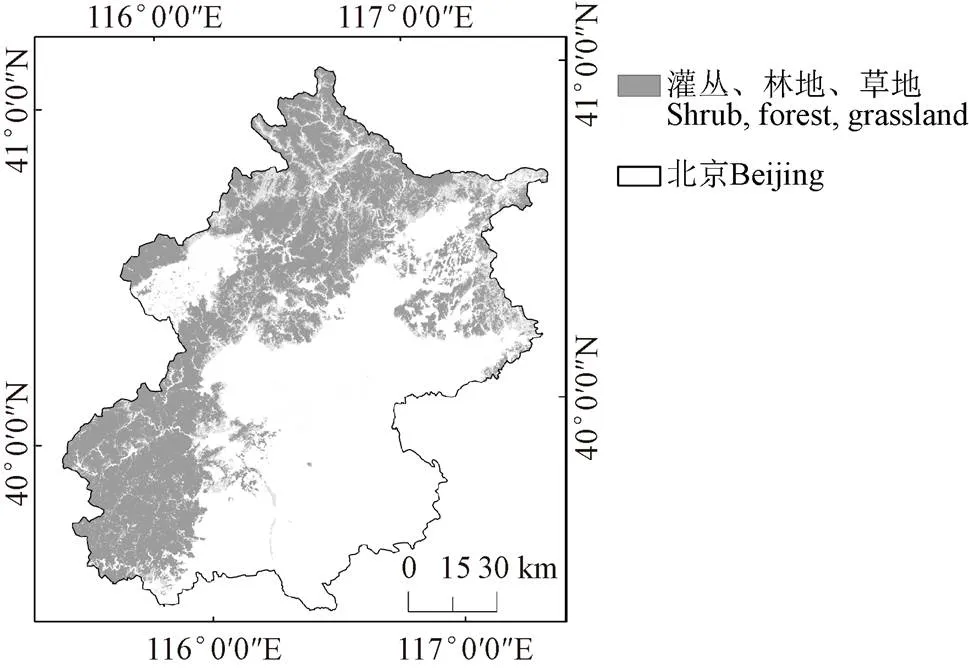
图1 研究区地理位置和土地利用图
1.3 研究方法
1.3.1 物候提取
MODIS NDVI数据集经过去噪声和最大合成处理后仍然存在一些噪声。这还包括在某些情况下存在的云污染。为了消除云、雪、冰污染造成的误差,进一步将Savitzky-Golay[22]滤波程序应用于每个年度NDVI周期,对NDVI时间序列进行平滑重构。动态阈值法对物候提取具有很大的灵活性和适用性[16]。本文通过Timesat软件,基于MODIS NDVI数据提取植被物候信息。根据前人关于北京地区物候观测数据和动态阈值设定,分别选用40%作为SOS阈值,45%作为EOS阈值,滑动窗口设置为5[23]。即NDVI曲线上升阶段,NDVI最大值与最小值差值的40%对应的时期定义为植被 SOS,NDVI曲线下降阶段,NDVI最大值与最小值差值的45%对应的时期定义为植被EOS。
1.3.2 基于CASA模型的NPP估算
利用基于光能利用率理论的CASA(Carnegie-Ames-Stanford-Approach)模型逐像元计算北京2001—2020年每月NPP,该模型适用于区域尺度的遥感研究,在目前国际上计算NPP研究中被广泛应用[24-25]。该模型是由吸收的光合有效辐射(Absorbed Photosynthetically Active Radiation,APAR,MJ/m2)和光利用效率(g/MJ)确定,本研究最大光能利用效率设置为0.389 g/MJ,具体见文献[17]。为了验证该模型的模拟精度,本研究获取了2001—2018年基于EC-LUE(Eddy Covariance-Light Use Efficiency)模型,得到的全球地表卫星(Global Land Surface Satellite,GLASS)NPP(http://www.glass.umd.edu/Download.html)产品。该产品空间分辨率为500 m,时间分辨率为8 d,经累加计算得到月尺度NPP,并采用线性回归利用GLASS数据集NPP对CASA模型模拟NPP结果进行评价。
1.3.3 趋势分析
使用非参数Theil-Sen和Mann-Kendall趋势分析方法对NPP和物候指标(SOS、PPT和EOS)的年际趋势和显著性进行分析,这种趋势分析方法可以减少数据异常值的影响,被认为是一种可靠的非参数统计趋势计算方法[26-27],逐像元计算NPP、SOS、LOS和EOS的趋势坡度,即年际变化率,这种非参数方法不需要数据序列的正态性,广泛应用于气候数据和植被数据的趋势分析。
1.3.4 气候、物候和NPP相关性分析
通过线性回归分析和逐像元的相关性分析,探讨NPP与气象因子和物候因子的关系。为了更深入地了解物候因素、气象因素对NPP直接、间接影响和贡献程度,利用AMOS(Analysis of Moment Structure)软件,采用标准化的结构方程模型(Strcture Equation Modeling,SEM)对其进行分析。SEM是一种建立估计和检验因果关系的模型,也是基于变量的协方差矩阵来分析变量之间关系的一种统计方法[28],其通过标准化路径系数量化各解释变量的相对贡献。本研究中结构方程建立之初包括所有可能的关系,通过设定模型、模型检验和模型修正,逐步删除相关性较小的关系,直到所有关系都具有较高的相关性,并且模型拟合参数最优。以下指标满足范围则认为模型可行:卡方自由度比<3;>0.05(>0.05说明数据不拒绝模型结构);拟合优度指数(Goodness of Fit Index,GFI)和标准化拟合指数(Normalized Fitting Index,NFI)越接近于1模型结构越好。
2 结果与分析
2.1 净初级生产力和气象插值结果评估
分别在CASA模型拟合的年NPP和GLASS数据集年NPP中随机抽取100个像元,共获得1800(18×100)个样本,结果表现出较好的拟合效果(2=0.57,<0.001),并且研究区域月值NPP拟合中两组数据具有较好的拟合效果(2=0.94,0.001)(图2)。中国青藏高原科学数据中心(Tibet Plateau Scientific Data Center,TPDC)气象数据集与克里金(kriging)插值气象数据进行线性拟合,结果显示年平均温度(2=0.82,0.001)、年降雨量(2=0.73,0.001)和年总辐射(2=0.69,0.001)在两个数据集的拟合中效果良好(图3)。

注:图2a中数据点代表研究区的月NPP,图2b中数据点为随机抽取100个像元中每个像元的年NPP。

注:TPDC为中国青藏高原科学数据中心气象数据集。
2.2 植被物候与净初级生产力的时空变化
北京植被2001—2020年NPP、SOS、EOS和LOS的年平均空间分异特征分别如图4。年平均NPP为507.99 g/m2,数值范围在370~570 g/m2之间,80%以上的区域在490~570 g/m2之间,北京北部区域多年平均NPP大于北京南部区域(图4a)。SOS年平均为第109天,主要在第97~125天内(图4b);EOS年平均为第286天,在第276~304天内(图4c);LOS年平均为176 d,有超过90%的区域在159~193 d(图4d);
研究区超过99 %的像元NPP逐渐增大,趋势显著的像元频率为41.6%(图5a);超过74%的像元SOS逐渐提前,趋势显著的像元频率为33.4%(图5b);超过93%的区域EOS逐渐推迟,趋势显著的像元频率为23.2%(图 5c);超过99%的区域内LOS有延长,趋势显著的像元频率为55.9%(图5d)。NPP在2001—2020年间显著增大(0.05)(图6a),平均每年增长2.98 g/(m2·a);SOS显著提前(0.05)(图6b),平均每年提前0.57 d;EOS显著推迟(0.05)(图6c),平均每年推迟0.51 d;LOS在20年间显著延长(0.05)(图6d),平均每年延长1.05 d。

a. 净初级生产力a. Net Primary Productivity(NPP)b. 生长季开始b. Start of growing season (SOS)c. 生长季结束c. End of growing season (EOS)d. 生长期长度d. Length of growing season (LOS)

a. NPPb. SOSc. EOSd. LOS

注:横坐标中0、5、10、15、20分别对应2000、2005、2010、2015、2020年。
2.3 物候因素与NPP的相关性
2001—2020年平均物候指标(SOS为第110天,EOS为第282天)为对照与每年物候指标相减,负值代表提前,正值代表推迟。SOS与NPP呈现显著负相关(2=0.61,0.05),SOS每提前一天,NPP增大3.74 g/(m2·a)(图 7a),但EOS与NPP关系不显著(2=0.16,=0.08)(图7b)。LOS与NPP显著正相关(2=0.45,<0.05),LOS每延长一天,NPP增长2.65 g/(m2·a)(图7c)。
在空间上(图8),79.77%的区域SOS与NPP年际变化呈现显著负相关(0.05),75.82%的区域LOS与NPP年际变化呈现显著正相关(0.05),但只有23.81%的区域EOS与NPP年际变化呈现显著正相关(0.05)。

注:负值代表提前,正值代表推迟。下同。

a. NPP-SOS b. NPP-EOSc. NPP-LOS
SOS与春季NPP为极显著负相关(2=0.61,0.001)(图9a),与夏季NPP没有显著的相关性(2=0.04,=0.42)(图9b),表明SOS提前并不会对夏季的NPP产生显著影响;EOS与秋季NPP为显著正相关(2=0.37,0.05)(图9d)。
在空间上(图10),94.37%的区域SOS与春季NPP年际变化呈现显著负相关(0.05)。SOS与夏季和秋季NPP在空间上分别有13.69%和6.76%的区域呈现显著负相关性,分别有64.5%和63.4%的区域呈现负相关但不显著。EOS与秋季NPP年际变化68.01%的区域呈现显著正相关(0.05)。

a. 春季NPP-SOSa. Spring NPP-SOSb. 夏季NPP-SOSb. Summer NPP-SOSc. 秋季NPP-SOSc. Autumn NPP-SOSd. 秋季NPP-EOSd. Autumn NPP-EOS
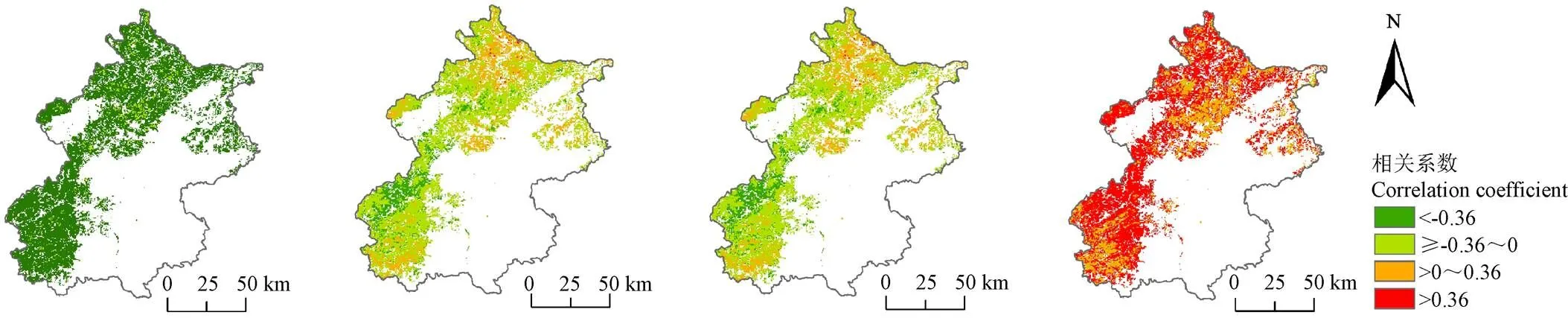
a. 春季NPP-SOSa. Spring NPP-SOSb. 夏季NPP-SOSb. Summer NPP-SOSc. 秋季NPP-SOSc. Autumn NPP-SOSd. 秋季NPP-EOSd. Autumn NPP-EOS
2.4 物候和气象因素对NPP的耦合影响
在年尺度上(图11a)SEM拟合参数卡方自由度比=1,=0.466,GFI=0.991,NFI=0.986;季节尺度上(图11b)SEM拟合参数卡方自由度比=0.94,=0.526,GFI=0.856,NFI=0.773。年总降雨量和年平均温度对SOS影响显著(0.05),SOS对年NPP影响显著(0.05)。年总降雨量对EOS影响极显著(0.001),但EOS对年NPP影响不显著(>0.05)。年总降雨量和年均温通过物候对NPP的间接影响,大于对NPP的直接影响。年均温和年降雨量通过SOS对NPP的间接影响远大于通过EOS对NPP的间接影响,说明增温和降雨主要作用于SOS提前,进而导致年NPP增加。春季平均温度通过SOS对NPP的间接影响(0.46)大于春季和夏季平均温度对NPP的直接影响(-0.37)。虽然春季总降雨量、夏季总降雨量和秋季总降雨量均对EOS产生显著影响,但是EOS对NPP影响不显著。
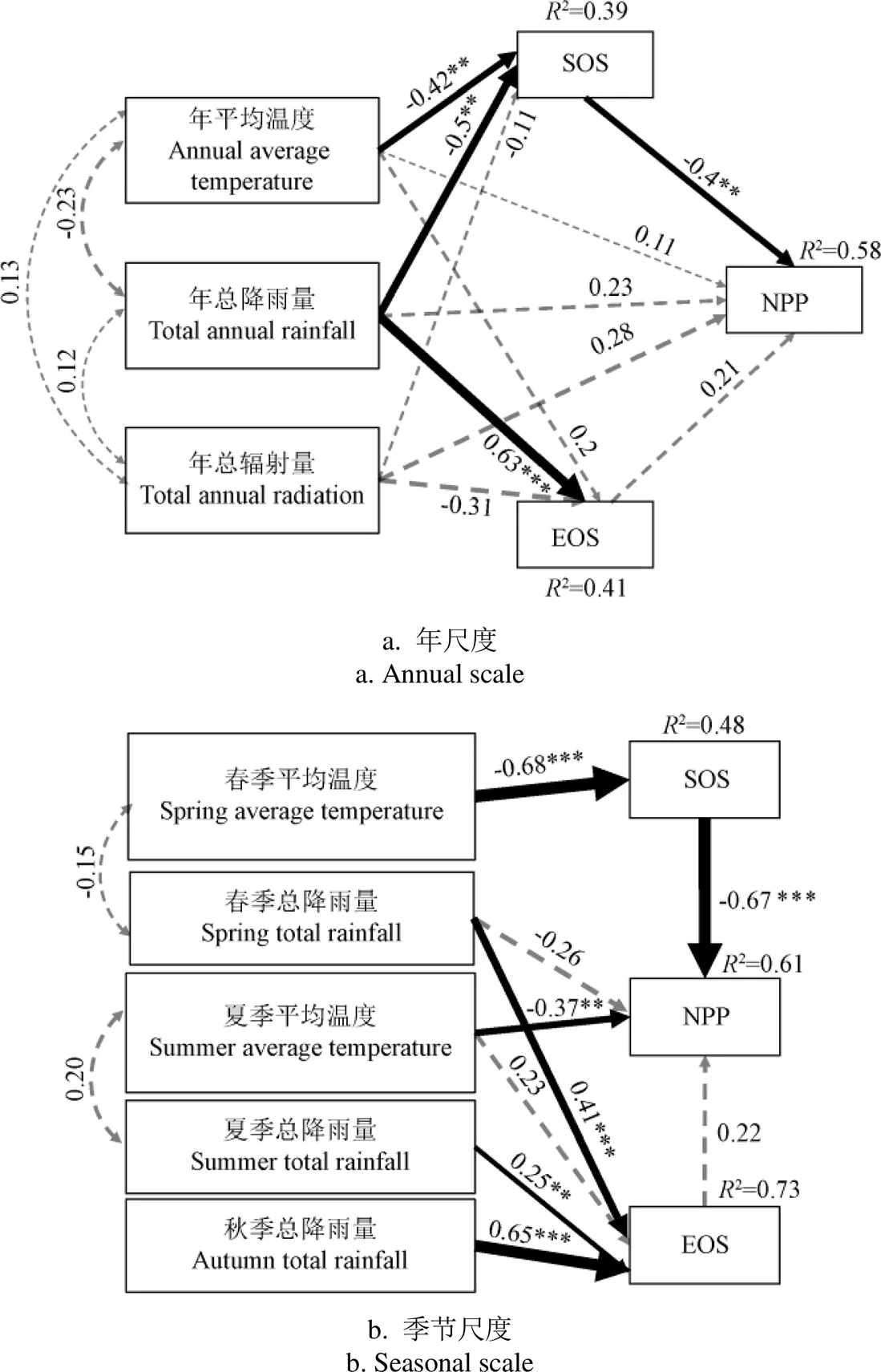
注:线上的数值为标准化通径系数,箭头粗细表示标准化系数的大小,实线表示关系显著(P <0.05),虚线表示不显著,星号***和**显著性水平分别为 0.001和0.05。R2表示有关变量的共同解释度。
3 讨 论
3.1 物候和NPP的相关性分析
研究区超过99%的区域生长季长度逐渐延长,在其他地区物候的相关研究中,生长季长度有出现缩短[29]、延长[30],但都表现为SOS提前。SOS趋势显著的像元频率高于EOS,前人研究也发现SOS受到冷积温和热积温的双重影响变异性强于EOS[31]。NPP年际变化的原因受到植物自身生理特性和环境因素等的影响,其中气候变化和物候因素是NPP逐年增长的重要原因[32]。有研究发现生长季的延长并不一定会增加生态系统的碳吸收,但会因为水分胁迫等因素对植被固碳产生影响[16]。早春可能会增加蒸腾速率,导致夏季土壤水分减少,进而导致生产力降低[33],但也有研究发现,生产力的抑制作用发生在秋季,而不是夏季[34],其可能的原因是由于早春水分耗散,增强了秋季水分胁迫,或者由于秋季变暖延长生态系统的生长季长度,由此造成季末呼吸损失了早春的固碳量[7]。在本研究中,SOS、LOS与NPP相关性更强(0.05),并且逐像元分析结果也发现,北京植被超过75%的区域SOS、LOS与NPP显著相关(图7、8),因此推测年际NPP增长的主要物候原因在于SOS的提前。SOS与春季NPP极显著负相关(图9a),EOS与秋季NPP之间显著正相关(图9d),说明NPP会因为春季物候提前和秋季物候延迟均会增长,而不会由于水分可用性的减小而降低,可能是由于北京属于温带季风气候区,明显的特点是夏季高温多雨,因此夏季水分胁迫对NPP的作用较小,春季NPP增长消耗的水分不会对夏季和秋季NPP产生影响。因此基于北京充沛的夏季降雨,SOS提前和EOS推迟会增加植被固碳的时间,导致年际NPP增长。
3.2 物候和气候因素对NPP的耦合影响
降水与NPP呈显著正相关,是由于降水增加了土壤水分,满足了植被生长和生产力提高的水分需求,尤其是春末和夏季[11]。温度通过多个途径影响物候和植被生长[34],春季气温对NPP具有直接的正相关关系。春季气温升高,使种子的有效积温提前进而导致叶片萌发和展开提前[22],植被在全年固碳的时间增长,因此,春季温度也会通过改变物候因素间接对NPP产生影响,在本研究中,春季温度通过物候对NPP的间接影响远远大于直接影响(图11b)。秋季降雨对EOS有显著影响,EOS对NPP的影响不显著,但秋季降雨对NPP的直接影响远小于通过物候的间接影响(图11b),因此相对于气候因素对NPP的直接影响来说,其通过秋季物候对NPP的间接影响更大。综上,在春季和秋季,气候通过物候(SOS、EOS)对NPP的间接影响大于直接影响。这在之前的研究中也有相似的发现,气候通过物候对生态系统碳循环的间接影响会比直接影响更大[35]。夏季气温升高导致植物蒸发加速缺水,会直接影响植被生长[33],本研究发现夏季温度对NPP具有显著负相关影响(图 11b),并且物候因素对其的影响很小(图11a),因此夏季气象因素对NPP的直接影响大于通过物候的间接影响。气候变暖对NPP的影响受到物候和环境因素的共同调控,其通过改变光合作用和生物过程直接和间接的影响生态系统碳平衡[36],在不同的环境中,早春生长可能对随后的每个季节的生态系统生产力产生混合影响[14]。春季物候通过直接和间接调节季节碳动态的地位不容忽视,气候对年际NPP的影响有很大一部分是通过物候间接调控的[37],对于NPP的模拟研究中物候对其产生的影响应得到重视。
4 结 论
北京植被生长季开始(Start of Growing Season,SOS)在2001—2020年间显著提前,平均每年提前0.57 d;生长季结束(End of Growing Season,EOS)显著推迟,平均每年推迟0.51 d。北京植被净初级生产力(Net Primary Productivity,NPP)在2001—2020年逐年增长,多年平均值在空间上北部区域大于南部区域。对于北京地区而言,生长期提前会增加植被碳固存的时间,导致NPP年总量增加,而不会产生负向影响。春季物候在调节NPP年际变化中占据重要地位,并且NPP年总量由物候引起的增长主要原因在于SOS提前。本研究为研究全球变暖条件下植被物候变化对NPP的影响提供了重要信息,有助于进一步认识植被NPP对气候变化的响应。
[1] Wang K, Wang X H, Piao S L, et al. Unusual characteristics of the carbon cycle during the 2015-2016 El Nino[J]. Global Change Biology, 2021, 27 (16): 3798-3809.
[2] Wang L Q, Hans J D B, Chen L X, et al. Urban warming increases the temperature sensitivity of spring vegetation phenology at 292 cities across China[J]. Science of the Total Environment, 2022, 834: 155154.
[3] Li M H, Wang X Y, Chen J C. Assessment of grassland ecosystem services and analysis on its driving factors: A case study in Hulunbuir Grassland[J]. Frontiers in Ecology Evolution, 2022, 10: 841943.
[4] Frank D, Reichstein M, Bahn M, et al. Effects of climate extremes on the terrestrial carbon cycle: Concepts processes and potential future impacts[J]. Global Change Biology, 2015, 21: 2861-2880.
[5] 尹小君,祝宏辉,Gao Gerry,等. 气候变化和人类活动对天山北坡净初级生产力变化的影响[J]. 农业工程学报,2020,36(20):195-202.
Yin Xiaojun, Zhu Honghui, Gao Jerry, et al. Effects of climate change and human activities on net primary productivity in the Northern Slope of Tianshan, Xinjiang, China[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2020, 36(20): 195-202. (in Chinese with English abstract)
[6] 徐勇,郑志威,戴强玉,等. 顾及时滞效应的西南地区植被 NPP 变化归因分析[J]. 农业工程学报,2022,38(9):297-305.
Xu Yong, Zheng Zhiwei, Dai Qiangyu, et al. Attribution analysis of vegetation NPP variation in Southwest China considering time-lag effects[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2022, 38(9): 297-305. (in Chinese with English abstract)
[7] Richardson A D, Anderson R S, Arain M A, et al. Terrestrial biosphere models need better representation of vegetation phenology: results from the North American Carbon Program Site Synthesis[J]. Global Biogeochemical Cycles, 2012, 18 (2): 566-584.
[8] Liu Q, Piao S, Janssens I A, et al. Extension of the growing season increases vegetation expo‐sure to frost[J]. Nature Communications, 2018, 21(6), 2366-2378.
[9] Piao S L, Liu Q, Chen A, et al. Plant phenology and global climate change: Current progresses and challenges[J]. Global Change Biology, 2019, 25 (6): 1922-1940.
[10] Piao S, Fang J, Zhou L, et al. Variations in satellite-derived phenology in China's temperate yegetation[J]. Global Change Biology, 2006, 12: 672-685.
[11] Yang J, Zhang X C, Luo Z H, et al. Nonlinear variations of net primary productivity and its relationship with climate and vegetation phenology[J]. China. Forests, 2017, 8 (10): 361.
[12] Shen M, Piao S, Cong N, et al. Precipitation impacts on vegetation spring phenology on the Tibetan Plateau[J]. Global Change Biology, 2015, 21 (10): 3647-3656.
[13] Buermann W, Forkel M, O’Sullivan M, et al. Widespread seasonal compensation effects of spring warming on northern plant productivity[J]. Nature, 2018, 562: 110-114.
[14] Yu Z, Lu C, Cao P, et al. Earlier leaf-flushing suppressed ecosystem productivity by draining soil water in the Mongolian Plateau[J]. Agricultural and Forest Meteorology, 2018, 250: 1-8.
[15] Zhou X, Geng X, Yin G, et al. Legacy effect of spring phenology on vegetation growth in temperate China[J]. Agricultural and Forest Meteorology, 2020, 281: 107845.
[16] Richardson A D, Black T A, Ciais P, et al. Influence of spring and autumn phenological transitions on forest ecosystem productivity[J]. Philosophical Transactions of the Royal Society B-biological Sciences, 2010, 365: 3227-3246.
[17] 朱文泉,潘耀忠,张锦水. 中国陆地植被净初级生产力遥感估算[J]. 植物生态学报,2007,31(3):413-424.
Zhu Wenquan, Pan Yaozhong, Zhang Jinshui. Estimation of net primary productivity of chinese terrestrial vegetation based on remote sensing[J]. Journal of Plant Ecology, 2007, 31(3): 413-424. (in Chinese with English abstract)
[18] 李虹,冯仲科,唐秀美,等. 区位因素对绿地降低热岛效应的影响[J]. 农业工程学报,2016,32(2):316-322.
Li Hong, Feng Zhongke, Tang Xiumei, et al. Effect of green space location factors on reducing urban heat island effect[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2016, 32(2): 316-322. (in Chinese with English abstract)
[19] 方精云. 碳中和的生态学透视[J]. 植物生态学报,2021,45: 1173-1176.
Fang Jinyun. Ecological perspective of carbon neutralization[J]. Chinese Journal of Plant Ecology, 2021, 45: 1173-1176. (in Chinese with English abstract)
[20] 李艳,张成才,罗蔚然,等. 基于改进最大值法合成 NDVI 的夏玉米物候期遥感监测[J]. 农业工程学报,2019,35(14): 159-165.
Li Yan, Zhang Chengcai, Luo Weiran, et al. Summer maize phenology monitoring based on normalized difference vegetation index reconstructed with improved maximum value composite[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2019, 35(14): 159-165. (in Chinese with English abstract)
[21] 李贵才. 基于MODIS数据和光能利用率模型的中国陆地净初级生产力估算研究[D]. 北京:中国科学院研究生院,2004.
Li Guicai. Estimation of Chinese Terrestiral Net Pirmary Production Using LUE Model and MODIS data[D]. Beijing: The Graduate School of the Chinese Academy of Sciences. 2004. (in Chinese with English abstract)
[22] Chen J, Jonsson P, Tamura M, et al. A simple method for reconstructing a high-quality NDVI time-series data set based on the Savitzky-Golay filter[J]. Remote Sensing of Environment, 2004, 91 (3): 332-344.
[23] 徐珂. 基于 MODIS 影像的北京地区生长季及其影响因子研究[D]. 北京:北京林业大学,2012: 1-55.
Xu Ke. Study on Growing Season and Its Influencing Factors in Beijing based on MODIS Images[D]. Beijing: Beijing Forestry University, 2012: 1-55. (in Chinese with English abstract)
[24] 刘真真,张喜旺,陈云生,等. 基于 CASA 模型的区域冬小麦生物量遥感估算[J]. 农业工程学报,2017,33(4):225-233.
Liu Zhenzhen, Zhang Xiwang, Chen Yunsheng, et al. Remote sensing estimation of biomass in winter wheat based on CASA model at region scale[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2017, 33(4): 225-233. (in Chinese with English abstract)
[25] Potter C S, Randerson J T, Field C B, et al. Terrestrial ecosystem production: A process model based on global satellite and surface data[J]. Global Biogeochemical Cycles, 1993, 7(4): 811-841.
[26] Theil H. A rank-invariant method of linear and polynomial regression analysis[J]. Springer Netherlands, 1950, 53: 386-392.
[27] Sen PK. Estimates of the regression coefficient based on Kendall’s Tau[J]. Journal of the American Statistical Association, 1968, 63(324): 1379-1389.
[28] 乔英,马英杰,辛明亮. 基于改进S-W与结构方程模型的干旱区枣园蒸散特征分析[J]. 农业机械学报,2021,52(8):307-317.
Qiao Ying, Ma Yingjie, Xin Mingliang. Analysis of evapotranspiration characteristics of ziziphus jujuba Mill. Orchards in arid areas based on improved S-W and structural equation model[J]. Transactions of the Chinese Society for Agricultural Machinery, 2021, 52(8): 307-317. (in Chinese with English abstract)
[29] Bao G, Chen J, Chopping M, et al. Dynamics of net primary productivity on the Mongolian Plateau: Joint regulations of phenology and drought[J]. International Journal of Applied Earth Observation and Geoinformation, 2019, 81: 85-97.
[30] Wang X F, Xiao J F, Li X, et al. No trends in spring and autumn phenology during the global warming hiatus[J]. Nature Communications, 2019, 10: 2389.
[31] Wu L Z, Ma X F, Dou X, et al. Impacts of climate change on vegetation phenology and net primary productivity in arid Central Asia[J]. Science of the Total Environment, 2021, 796: 149055.
[32] Duveneck M J, Thompson J R. Climate change imposesphenological trade-offs on forest netprimary productivity[J]. Journal of Geophysical Research: Biogeosciences, 2017, 122: 2298-2313.
[33] Xu H J, Wang X P, Yang T B. Trend shifts in satellite-derived vegetation growth in central eurasia, 1982-2013[J]. Science of the Total Environment, 2017, 579: 1658-1674.
[34] Shi P, Chen Z, Reddy G V P, et al. Timing of cherry tree blooming: Contrasting effects of rising winter low temperatures and early spring temperatures[J]. Agricultural and Forest Meteorology, 2017, 240: 78-89.
[35] Shao J, Zhou X, Luo Y, et al. Biotic and climatic controls on interannual variability in carbon fluxes across terrestrial ecosystems[J]. Agricultural and Forest Meteorology, 2015, 205: 11-22.
[36] Niu S, Fu Z, Luo Y, et al. Interannual variability of ecosystem carbon exchange: From observation to prediction[J]. Global Change Biology, 2017, 26: 1225-1237.
[37] Wu C, Hou X, Peng D, et al.Land surface phenology of china’s temperate ecosystems over 1999-2013: Spatial-temporal patterns, interaction effects, covariation with climate and implications for productivity[J]. Agricultural and Forest Meteorology, 2016, 216: 177-187.
Responses of the net primary productivity of vegetation to phenological changes in Beijing of China
Wei Xiaoshuai1,2, Gao Yonglong3, Fan Yaqian4, Lin Ling3, Mao Jun1,2, Zhang Dehuai5, Li Xinhao1,2, Liu Xinyue1,2, Xu Mingze1,2, Tian Yun1,2, Liu Peng1,2Jia Xin1,2, Zha Tianshan1,2※
(1.,,100083,;2.,,100083,;3.,100013,;4.,102100,; 5.,101599,)
Net Primary Productivity (NPP) of vegetation is considered one of the main indicators for the carbon fixation capacity of ecosystems in the carbon cycle, particularly for the adaptability of ecosystems to climate change. Among them, the typical phenological factors are the key components of the ecosystem functions in many biological processes, including the Start of the Growing Season (SOS), End of the Growing Season (EOS), and Length of the Growing Season (LOS). However, it is still lacking in the relative importance of phenological and climatic factors to the NPP. The contribution of phenological factors (SOS, EOS, and LOS) to the interannual change of NPP has not been well quantified, due to the complex ecosystem. Therefore, this study aims to extract the phenological information of vegetation using a Cardiovascular-Ames-Stanford Approach (CASA) model, in order to examine the characteristics of spatial and temporal changes of NPP. The Normalized Vegetation Index (NDVI) was used from the MODIS data in Beijing from 2001 to 2020. The interaction between meteorological factors, phenological changes, and NPP was then explored using linear regression, trend analysis, and structural equation modeling. The results show that the SOS was gradually advanced by 0.57 each year on average from 2001 to 2020 over more than 70% of the regions, whereas, the EOS was gradually postponed by an average of 0.51 days per year over more than 90% of the regions. The NPP vegetation increased significantly from 2001 to 2020 (< 0.05), where the growth rate was greater in the last 10 years than that in the first 10 years. The average annual NPP value was greater in the northern region than that in the southern. There was a strong correlation between the phenological factors (SOS, and LOS) and NPP (<0.05). The pixel-by-pixel analysis also found that the SOS, LOS, and NPP were significantly correlated in the areas with more than 75% vegetation coverage. The NPP was also significantly affected by the advance of SOS and extension of LOS (<0.05). The NPP increased by 3.74 g/m2for every single day advance of SOS, while by 2.65 g/m2for every single day extension of LOS. There was no significant effect of the EOS delay in autumn on the NPP. A coupling effect of phenology and climatic factors varied with the season. There was a greater indirect effect of climate through changing phenology (SOS and EOS) on the NPP in spring and autumn, compared with the direct. By contrast, the NPP was more directly affected by climatic factors, temperature, and rainfall in summer. In conclusion, the spring phenological change was an important driving factor for the interannual variation in the NPP. Furthermore, the annual NPP increased to the phenological advance. The findings can also provide an important supplement to determine the vegetation productivity response to the climate change in urban areas.
remote sensing; net primary productivity; vegetation phenology; climate change; NDVI
10.11975/j.issn.1002-6819.2022.18.018
S7
A
1002-6819(2022)-18-0167-09
魏晓帅,高永龙,范雅倩,等. 北京植被净初级生产力对物候变化的响应[J]. 农业工程学报,2022,38(18):167-175.doi:10.11975/j.issn.1002-6819.2022.18.018 http://www.tcsae.org
Wei Xiaoshuai, Gao Yonglong, Fan Yaqian, et al. Responses of the net primary productivity of vegetation to phenological changes in Beijing of China[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2022, 38(18): 167-175. (in Chinese with English abstract) doi:10.11975/j.issn.1002-6819.2022.18.018 http://www.tcsae.org
2022-06-08
2022-09-12
国家重点研发计划项目(2020YFA0608100)
魏晓帅,博士生,研究方向为生态学遥感。Email:weixiaoshuai@bjfu.edu.cn
查天山,教授,博士生导师。研究方向为干旱半干旱生态系统碳水循环、植物生理生态。Email:tianshanzha@bjfu.edu.cn
猜你喜欢
农业工程学报(2022年12期)2022-09-09
热带作物学报(2022年6期)2022-07-08
海洋通报(2021年5期)2021-12-21
空间科学学报(2021年2期)2021-07-21
西南农业学报(2017年5期)2017-06-23
红土地(2016年3期)2017-01-15
中国学术期刊文摘(2016年1期)2016-02-13
中国学术期刊文摘(2016年8期)2016-02-13
环球市场信息导报(2015年15期)2015-12-03
应用海洋学学报(2015年4期)2015-11-24
