
短枝型富士苹果枝条导管分子特征参数及其与树体生长的关系
2022-01-26 10:00付晓雅朱玉宁李中勇刘文田徐继忠
河南农业科学 2021年11期
付晓雅,沈 震,朱玉宁,李中勇,刘文田,徐继忠
(1. 河北农业大学园艺学院,河北 保定 071001;2. 河北省曲阳县农业农村局,河北 曲阳 073100)
矮化密植是世界苹果栽培发展的总趋势,应用矮化品种是实现密植栽培的重要途径之一[1‐2]。短枝型苹果以短枝结果为主,树体矮小,树形紧凑,短枝系数和萌芽率高,结果早,产量好[3‐4],比普通型苹果矮化20%或更多[5]。短枝型苹果多数由芽变而来,少数是经诱变、杂交选育等手段获得[6]。这类资源与乔砧结合,采用密植栽培可克服乔砧密植单产低、果实品质差、费工、技术复杂、推广难度大等问题;与矮化中间砧相结合,实行“双矮”密植栽培可实现早果、丰产[7‐8]。多年来,国内外在短枝型苹果品种选育、特性特征、变异机制以及遗传机制研究等方面都取得了较大进展[9]。
导管分子是植物木质部的重要组成细胞之一,其主要功能是输导水分和矿质元素,木质部水分输导功能主要受导管形态结构的影响。因此,研究导管分子形态结构对揭示树体致矮机制有重要意义。杨佩芳等[10‐11]对苹果短枝型品种研究结果表明,短枝型品种较普通型品种导管分子显著地窄而短,导管细胞密度、导管直径明显小于普通型品种。陈静等[12]对不同品种苹果枝条解剖发现,凡导管密度大、所占木质部百分比高的品种,生长势强于密度小、所占木质部百分比低的品种。郭学民等[13]研究表明,矮生桃树导管分子长度与直径均比乔生桃树小。王秀娟等[14]发现,矮生型梨砧木导管分子长度与直径显著小于普通型梨砧木。但是,前人研究多集中于导管分子特征参数或管腔大小某一方面,内容单一,不够系统,关于短枝型富士与普通型富士导水率差异更是鲜见报道。鉴于此,选择普通型富士和短枝型富士作为试材,对不同品种的树体大小、枝类组成、导管特征参数及最大导水率进行比较分析,从解剖方面研究其影响输水效率的机制,探索短枝型品种矮生机制,从而为植物生长发育规律研究提供解剖学证据。
1 材料和方法
1.1 试验材料
试验在河北省保定市曲阳县刘家马村绿阳果园进行。供试材料选用普通型品种富士冠军、昌红,短枝型品种礼泉短枝、天红2号,嫁接在SH40中间砧上,树龄为5 a,株行距为2 m×4 m,果园为常规管理,灌溉便利。选取树势健壮、长势一致苹果树为试材,每个品种设3 次重复。选取树体中上部外围1 年生枝,每棵树随机选取3~5 个枝条,带回实验室用于导管特征的观察、导水率的测定。
1.2 试验方法
1.2.1 树体结构参数 于2019—2020 年连续2 a 对富士系品种进行树相调查,具体方法为秋季落叶后用钢卷尺测量树高、枝展、外围1 年生枝长。树高:测量中间砧嫁接口到生长点之间的距离;枝展:分别测量东西、南北最外围枝条之间距离;外围1年生枝长度:随机选取树冠外围1 年生枝条10 个,用卷尺进行测量。干周:用卷尺测量中间砧嫁接口上部10 cm 处的粗度;枝类组成调查方法:落叶后分别统计单株树体的短枝(<5.0 cm)、中枝(5.0~15.0 cm)、长枝(>15.0 cm)数量。
1.2.2 枝条解剖结构 新鲜材料用FAA 液固定,用于解剖结构观察。徒手切片,然后利用显微摄像系统照相,用图像处理软件进行木质部导管个数、木质部导管管腔面积统计,每棵树重复9次,导管密度为单位面积内导管个数,管腔直径为同面积圆的直径,导管总面积占木质部比例=管腔面积×导管密度×100%。
1.2.3 导管分子形态特征观察 供试材料为枝条中部茎段,去皮后,将木质部切成长×宽×高为1.0 cm×0.3 cm×0.3 cm 的小段作为样品。将样品放入青霉素小瓶中,加入10%铬酸和10%硝酸等量离析液,置于30 ℃培养箱中离析1~2 d,中间可多次更换离析液,材料软化后,用清水反复漂洗至酸性去除,离析后的材料放于50%乙醇中保存备用。观察时材料从50%乙醇中取出,1%番红染色,制作临时装片。用上海比目仪器有限公司产BM2030E 型生物显微镜观察,并采用ToupCam 摄像装置拍摄,显微图像测量分析软件Toup-View 测量。试验设3次重复,每个重复随机测量90 个导管分子的长度、宽度、端壁尾长、端壁倾斜角,取平均值,并统计导管分子中单尾导管数、无尾导管数、双尾导管数,计算每种导管分子占比。
1.2.4 导水率测定 参照靳欣[15]的方法测定导水率。选取长度大于50 cm的枝条,在水中剪取3 cm长的茎段,用0.45 μm 微孔过滤器抽滤的0.025 mol/L KCl 溶液作为测定溶液和冲洗液。先用0.175 MPa压力差推动KCl 溶液流过枝条,冲去茎段所有气穴栓塞后,将压力调为5 kPa,使KCl 溶液在低压状态下流过茎段,流出液接到一个收集瓶中,该瓶放置在与电脑连接天平上,将茎段的直径和长度值输入电脑LPFM 软件,通过LPFM 测定软件获取天平读数随时间的变化值并计算枝条的导水率。测定时每5 s 记录一个值,直至其相对稳定,即为枝条的最大导水率。
1.3 数据分析
利用Excel和SPSS 20.0软件进行数据处理和统计分析。
2 结果与分析
2.1 富士苹果树体结构参数比较
2.1.1 不同类型树体大小 短枝型品种较普通型品种树体矮小,树形紧凑。由表1 可知,在2019—2020 年连续2 a 时间里,普通型品种富士冠军、昌红的树高显著高于短枝型品种礼泉短枝、天红2 号。2019年树高从大到小依次为富士冠军(336.33 cm)、昌红(331.00 cm)、天红2 号(267.00 cm)、礼泉短枝(263.00 cm),2020年树高从大到小依次为富士冠军(405.67 cm)、昌红(381.33 cm)、礼泉短枝(278.00 cm)、天红2 号(242.00 cm)。在连续2 a 中,普通型品种的干周也高于短枝型品种,2019 年,富士冠军、昌红的干周分别为20.96、21.98 cm,而礼泉短枝、天红2 号的干周为18.23、17.09 cm。2020 年,富士冠军、昌红的干周分别为27.37、28.23 cm,礼泉短枝、天红2号的干周分别为23.00、21.67 cm。在外围新梢长度方面,普通型品种高于短枝型,2019 年富士冠军长度最大,为44.60 cm,礼泉短枝长度最小,为14.30 cm。2020 年仍为富士冠军长度最大,为48.63 cm,天红2 号最小,长度为14.93 cm。2019—2020 年,普通品种的行内、行间枝展显著高于短枝型品种。
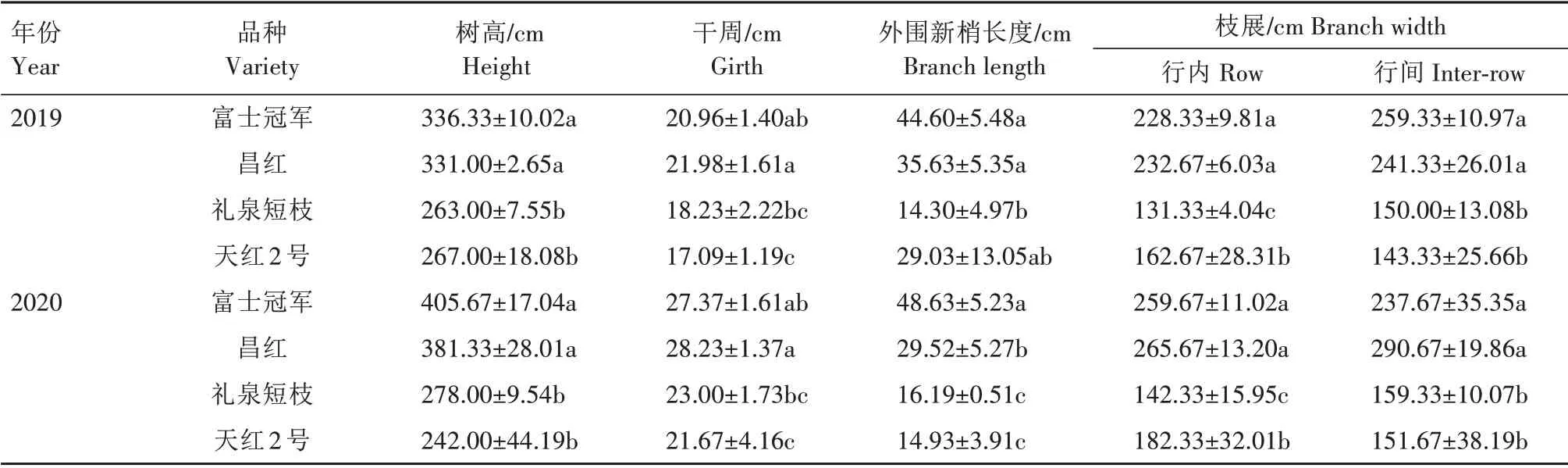
表1 普通型富士与短枝型富士树体大小Tab.1 Comparison of tree size between spur-type Fuji and standard-type Fuji
2.1.2 不同类型枝类组成 苹果以中短枝结果为主,短枝型品种与普通型品种相比有较多的短枝。由表2 可知,不同类型品种枝类组成有显著差异。短枝型品种的短枝比显著高于普通型品种,2019 年短枝比从大到小依次为天红2 号(81.37%)、礼泉短枝(80.44%)、富士冠军(53.74%)、昌红(51.02%),2020 年短枝比从大到小依次为天红2 号(81.10%)、礼泉短枝(79.23%)、昌红(67.11%)、富士冠军(57.89%)。普通型品种的长枝比显著高于短枝型品种,2019 年,普通型品种富士冠军的长枝比为31.26%,昌红为33.99%,短枝型品种礼泉短枝为12.22%,天红2 号为11.99%。2020 年,富士冠军长枝比为26.93%,昌红为18.70%,而礼泉短枝为10.64%,天红2号为7.57%。

表2 普通型富士与短枝型富士枝类组成Tab.2 Comparison of branch composition between spur-type Fuji and standard-type Fuji
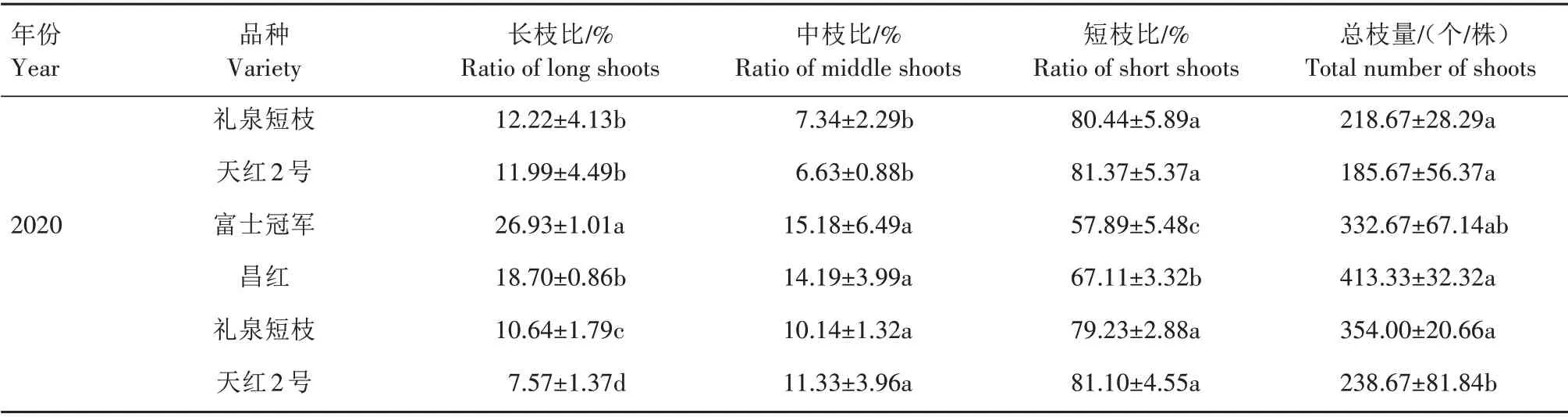
续表2 普通型富士与短枝型富士枝类组成Tab.2(Continued) Comparison of branch composition between spur-type Fuji and standard-type Fuji
2.2 富士苹果导管分子形态特征参数比较
2.2.1 管腔大小 通过徒手切片观察4个品种枝条横截面显微结构,可见管孔形状多为圆形或椭圆形,少数为多边形,管孔均呈分散型分布在射线内(图1)。由表3 可知,普通型品种导管密度高于短枝型品种。天红2 号密度最低,为365.25 个/mm2,昌红最高,为424.97 个/mm2。普通型品种的管腔面积、管腔直径高于短枝型品种,管腔面积依次为富士冠军>昌红>礼泉短枝>天红2 号,即652.19 μm2>604.83 μm2>532.32 μm2>509.42 μm2,管腔直径依次为富士冠军>昌红>礼泉短枝>天红2 号,即28.78 μm>27.69 μm>25.94 μm>25.34 μm。普通型品种富士冠军、昌红管腔总面积占木质部比例显著高于短枝型品种礼泉短枝、天红2 号,富士冠军为25.83%,昌红为25.56%,而礼泉短枝为20.05%,天红2 号为18.55%。以上结果表明,短枝型富士的导管密度、管腔面积、管腔直径、管腔总面积占木质部比例都要低于普通型富士。2.2.2 导管类型 在光学显微镜下可观察到,不同类型品种导管分子多为直径均匀的管状细胞,从导管分子侧壁次生增厚和木质化的方式看,普通型与短枝型品种导管分子均为孔纹导管。从穿孔板的式样可以看出,末端多为单穿孔(图2A—L、图3A—K、图3M),偶见双穿孔(图2M、图3L)。
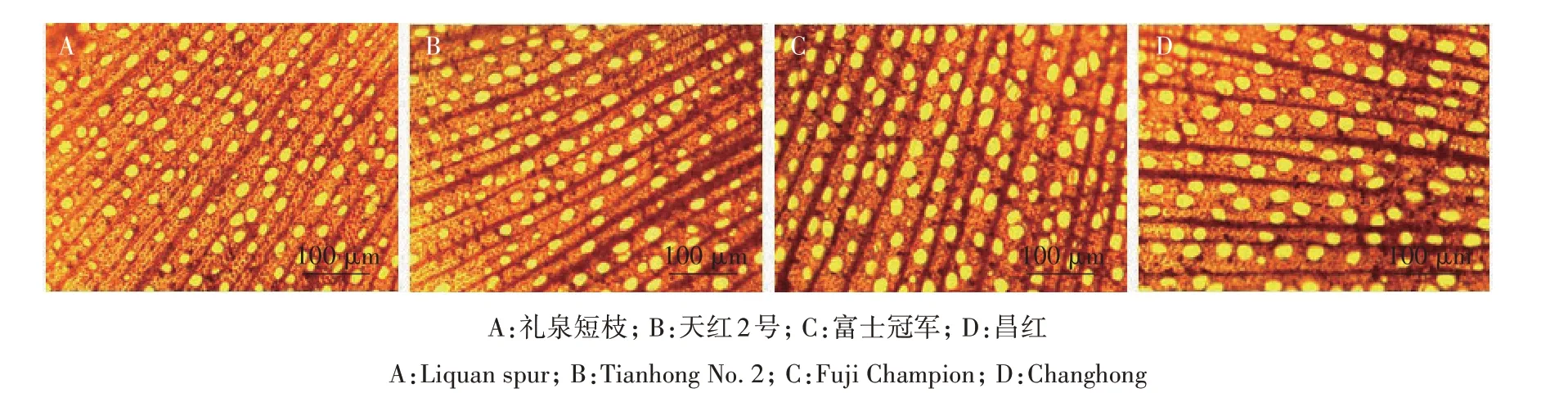
图1 普通型富士与短枝型富士1年生枝横截面Fig.1 Cross section of annual branches of spur-type Fuji and standard-type Fuji
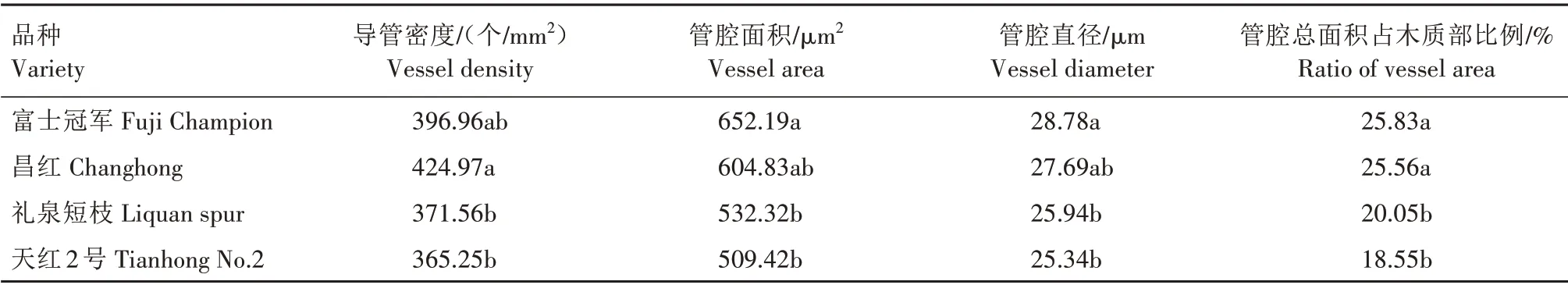
表3 普通型富士与短枝型富士枝条导管分子结构差异Tab.3 Difference of vessel element structure between spur-type Fuji and standard-type Fuji
根据具尾情况分为两端具尾(图2A、图2C—E、图2I、图2K—M、图3B—E、图3H—I、图3M)、一端具尾(一端无尾)(图2F—H、图3A、图3F、图3J—L)及两端无尾(图2B、图2J、图3G)3 种类型。由表4可知,普通型与短枝型品种导管分子具尾情况无明显差异,都以两端具尾类型为主,其次为一端具尾,两端无尾类型最少。在两端具尾导管类型中,普通型品种富士冠军、昌红所占比例分别为81.78%和82.89%,短枝型品种礼泉短枝和天红2 号比例分别为73.76%和86.04%;在一端具尾类型中,富士冠军与昌红所占比例分别为17.41%和16.23%,礼泉短枝和天红2号分别为22.05%和13.96%;在两端无尾类型中,普通型品种富士冠军、昌红所占比例分别为0.81%和0.88%,短枝型品种礼泉短枝和天红2号所占比例分别为4.19%和0。从导管分子尾的长短来看,2种类型品种尾长无明显差异。
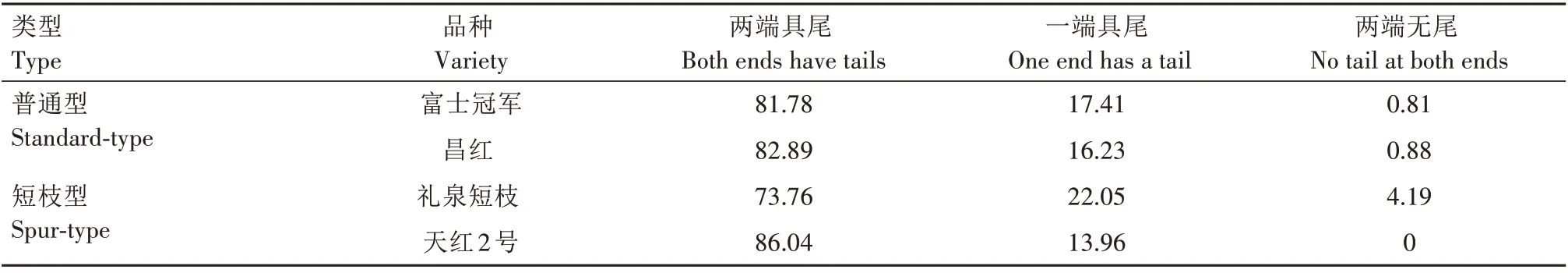
表4 短枝型富士与普通型富士导管分子具尾分布比例Tab.4 Proportion of end wall with tail of vessel element between spur-type Fuji and standard-type Fuji%
根据导管分子两端端壁斜度,可以分为两端倾斜(图2A、图2C—F、图2H—K、图2M、图3A、图3C—J、图3L—M)、一端倾斜(图2G、图2L、图3K)、两端水平(图2B、图3B)3 种类型(斜度小于15°视为水平)。由表5可知,普通型与短枝型品种导管都以两端倾斜类型为主,其次为一端倾斜,两端水平类型最少。在两端倾斜分布比例中,普通型品种富士冠军、昌红分别为94.28%和96.43%,短枝型品种礼泉短枝和天红2号分别为92.07%和94.83%;在一端倾斜类型中,富士冠军和昌红分别为4.29%和3.57%,礼泉短枝和天红2 号分别为6.10% 和4.02%;在两端水平分布比例中,普通型品种富士冠军、昌红分别为1.43%和0,短枝型品种礼泉短枝和天红2号分别为1.83%和0.71%。
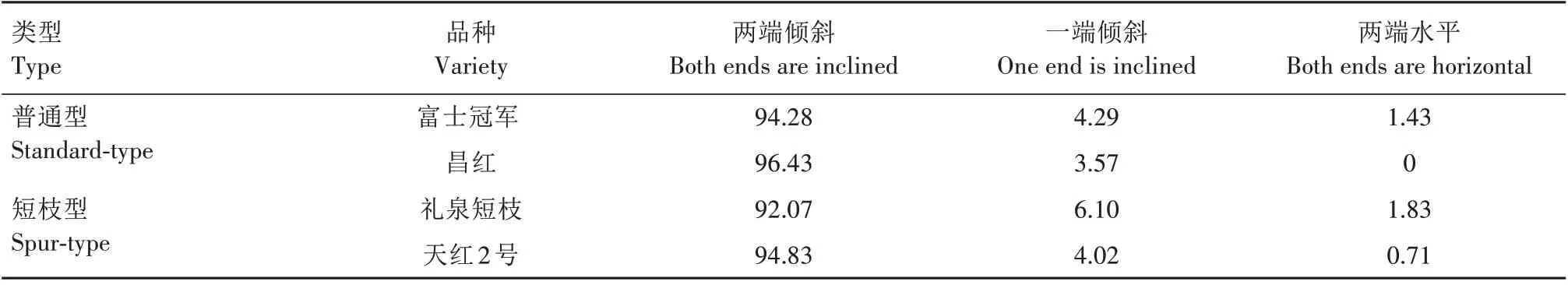
表5 短枝型富士与普通型富士导管分子端壁倾斜角分布比例Tab.5 Proportion of end wall inclination angle of vessel element between spur-type Fuji and standard-type Fuji%
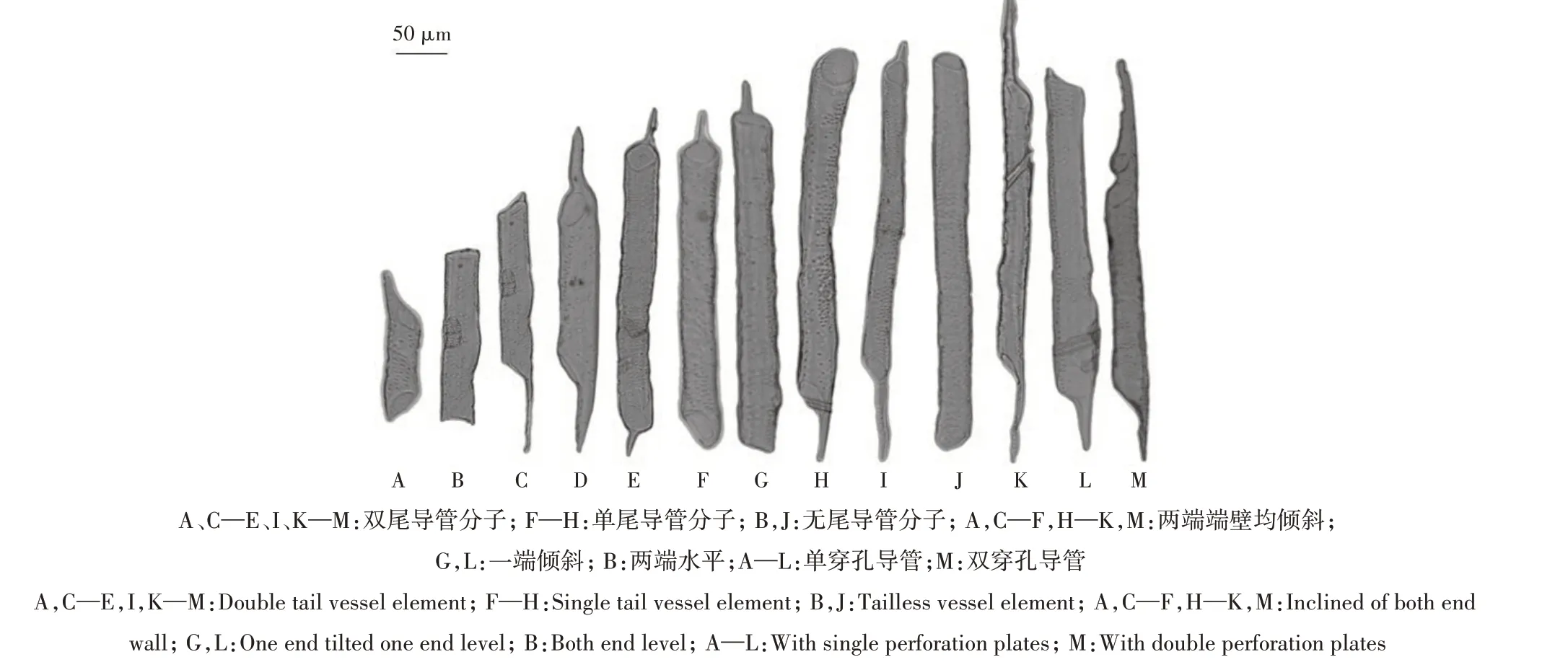
图2 普通型富士1年生枝导管分子形态Fig.2 Characteristic of vessel element in annual stem of standard-type Fuji

图3 短枝型富士1年生枝导管分子形态Fig.3 Characteristic of vessel element in annual stem of spur-type Fuji
2.2.3 导管分子长度与直径
2.2.3.1 导管分子长度与直径大小 由表6 可知,短枝型品种的导管分子长度与直径较普通型品种更为短小、狭窄。普通型富士导管分子长度为148.55~525.89 μm,短枝型富士为 105.34~523.69 μm。普通型富士冠军导管分子长度平均为333.20 μm,昌红为335.00 μm,短枝型富士礼泉短枝导管分子长度平均为319.38 μm,天红2 号为316.75 μm,短枝型富士导管分子长度显著小于普通型富士。普通型富士导管分子直径为22.68~54.07 μm,短枝型富士为21.87~50.03 μm,富士冠军导管分子平均直径为37.85 μm,昌红为37.89 μm,礼泉短枝为33.18 μm,天红2号为34.21 μm,短枝型富士导管分子平均直径显著小于普通型富士。
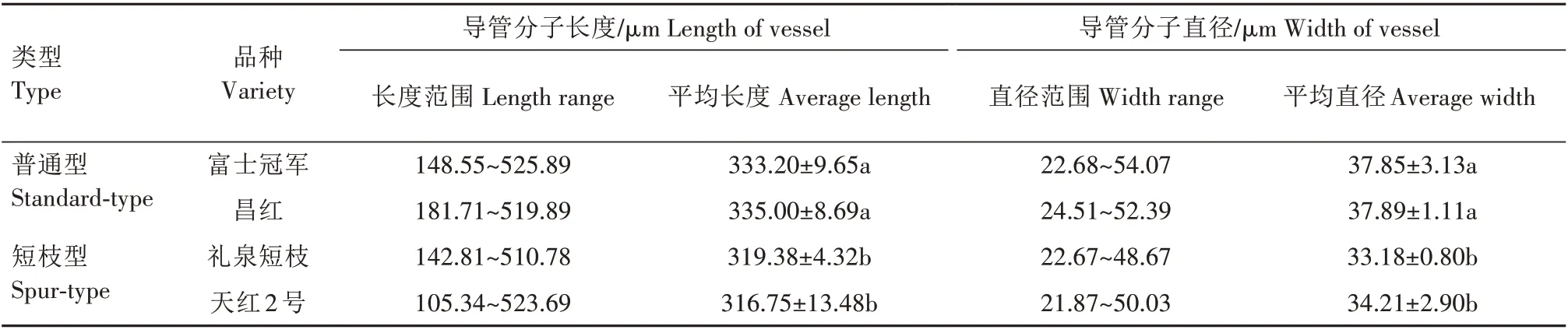
表6 短枝型富士与普通型富士导管分子长度和直径Tab.6 Comparison of length and width of vessel element between spur-type Fuji and standard-type Fuji
2.2.3.2 导管分子长度与直径分布频率 由表7 可知,各品种导管分子长度主要分布于301~450 μm,在0~150 μm 范围所占比例最小。导管分子直径主要分布于30.1~45.0 μm,在大于45.0 μm 范围所占比例最小。其中富士冠军与昌红导管分子长度在301~450 μm 所占比例分别为55.69%和60.99%,礼泉短枝与天红2 号分别为55.17%和60.22%。在大于450 μm 范围中富士冠军与昌红分别为8.13%和7.17%,礼泉短枝与天红2 号分别为3.45% 和3.79%,短枝型品种比普通型品种低3.38~4.68 个百分点;富士冠军与昌红30.1~45.0 μm 导管直径分别为69.51%和81.61%,礼泉短枝与天红2 号分别为72.03%和69.08%。在大于45.0 μm 范围中富士冠军为12.20%,昌红为9.42%,礼泉短枝和天红2 号分别为1.92%和6.87%,短枝型品种比普通型品种低2.55~10.28个百分点。
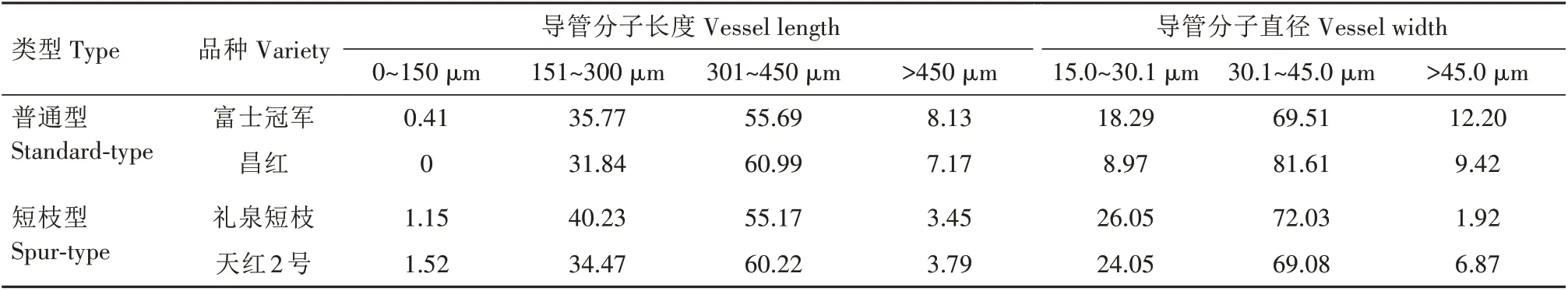
表7 短枝型富士与普通型富士导管分子长度和直径分布比例Tab.7 Distribution of length and width of vessel element between spur-type Fuji and standard-type Fuji%
2.2.4 导管分子端尾长度 如图4 所示,普通型富士冠军导管分子端尾长度显著高于短枝型礼泉短枝、天红2 号,为57.13 μm;而昌红导管分子端尾长度低于短枝型富士,为42.81 μm。统计分析表明,2种类型富士导管分子端尾长度变化无规律。
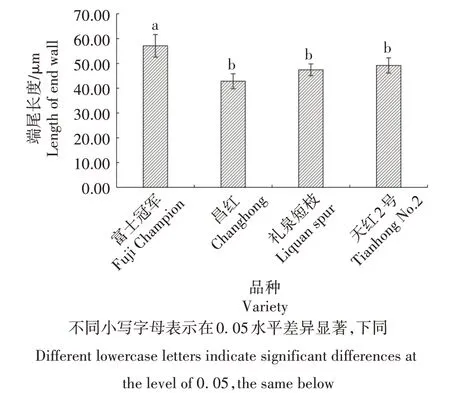
图4 不同类型富士导管分子端尾长度比较Fig.4 The end wall length of vessel element in different types of Fuji
2.2.5 导管分子端壁倾斜角 如图5 所示,短枝型品种的端壁倾斜角大于普通型品种,短枝型品种天红2 号的端壁倾斜角为48.78°,礼泉短枝为46.78°,普通型品种富士冠军的端壁倾斜角度为44.81°,昌红为44.51°。统计分析表明,短枝型品种导管分子端壁倾斜角较普通型品种更为倾斜。
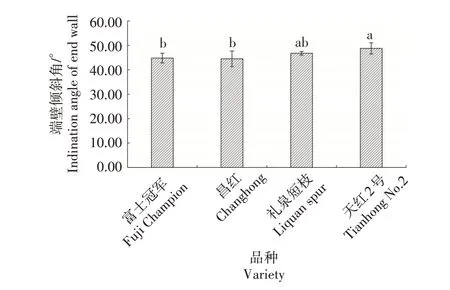
图5 不同类型富士导管分子端壁倾斜角比较Fig.5 The end wall inclination angle of vessel element in different types of Fuji
2.3 富士苹果最大导水率比较
如图6 所示,富士冠军的最大导水率最高,为5.65×10-5kg/(m·s·MPa),其次为昌红,最大导水率为5.31×10-5kg/(m·s·MPa)。礼泉短枝、天红2 号最大导水率分别为3.80×10-5、3.43×10-5kg/(m·s·MPa)。说明普通型品种最大导水率高于短枝型品种,结合2 种类型品种导管形态特征,表明水分长距离运输的效率和安全性与树木木质部导管的结构有直接关系。
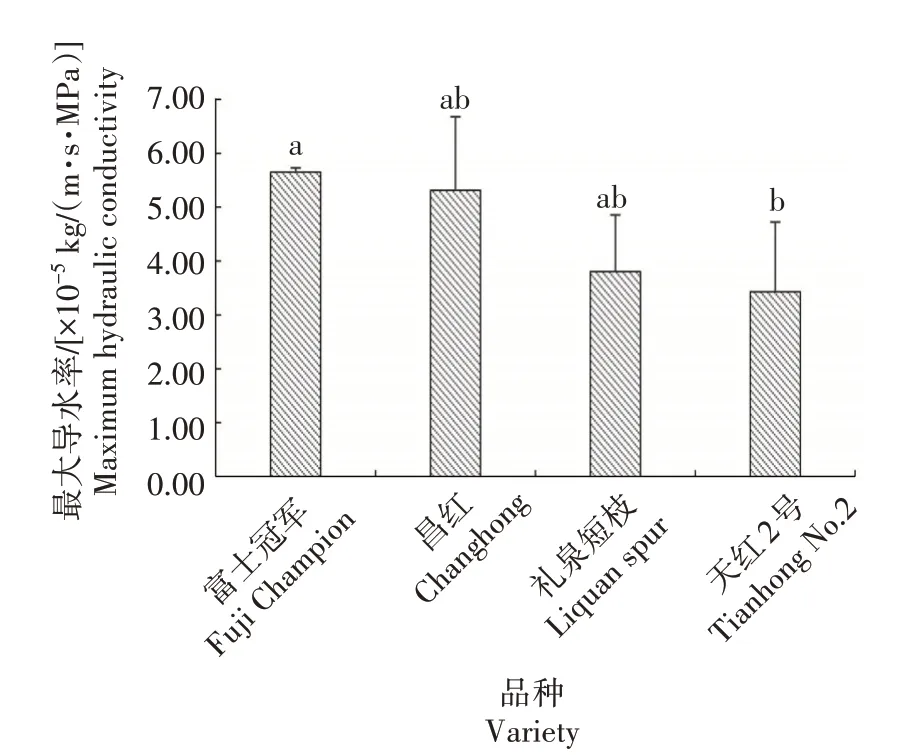
图6 不同类型富士最大导水率比较Fig.6 Maximum hydraulic conductivity in different types of Fuji
3 结论与讨论
导管是植物木质部输导水分和无机盐的管状结构,其输导作用和它的结构特征是紧密联系的,导管分子的长短直接决定运输阻力的大小以及转运次数的多少。导管分子直径越大,运送营养物质的量越大,速度越快[16]。本研究中,短枝型富士的导管分子长度与直径显著小于普通型品种,这与杨佩芳等[10]发现短枝型品种较普通型品种短而狭窄相一致。短枝型品种具有小导管性,这种结构在一定程度上限制了水分和无机盐类的上运,使短枝型品种树体矮小,生长势弱[17]。本研究发现,短枝型富士导管端壁倾斜角大于普通型品种,而端壁倾斜角的大小影响了管腔端壁的接触面积,端壁倾斜角度大,导管之间接触的面积大,降低了植物运输水分的效率[18‐19]。王浩等[20]研究结果表明,抗寒葡萄导管分子两端斜度小于非抗寒品种,这种导管特性可以提高输水效率,在冬季和早春保证植株对水分的需要。
本试验通过对1 年生枝解剖结构研究可知,短枝型品种的导管密度、管腔直径、管腔面积、管腔总面积占木质部比例明显小于普通型品种。王辉[21]在比较苹果乔化砧木与矮化砧木时,得出乔化砧木八棱海棠枝条导管分子管腔直径、管腔面积、导管总面积占木质部比例较矮化砧木高的结果,与本研究结果一致。管腔大小和导管密度在树体地上部水分输导中起着重要作用,管腔总面积占木质部比例高,说明木质部输导能力强,有助于单位时间内输送更多的水分[22]。
同时本研究中最大导水率的结果显示,普通型品种最大导水率大于短枝型品种,最大导水率是导管分子除去栓塞后导管分子的导水率,根据流体力学中Hagen-Poiseuille 方程,导水率与导管半径的4次方呈正相关[23],管腔直径越大,水分运输速率越高,所以宽的导管可以有效提高水分运输速率。王林等[24]通过对刺槐根、茎研究结果表明,刺槐根部导管分子直径大于茎部,从而使得根部的导水能力远大于茎部。
综上所述,正是由于短枝型品种拥有短而窄的导管、较低的导管密度与管腔面积,使得短枝型品种输水效率与运输养分的能力都要弱于普通型品种,地上部生长受到了抑制,造成短枝型富士在连续2 a 内的树高、干周、外围新梢长度、枝展都要小于普通型品种。
猜你喜欢
影像科学与光化学(2022年4期)2022-07-19
今日农业(2021年19期)2022-01-12
今日农业(2021年17期)2021-11-26
现代仪器与医疗(2021年4期)2021-11-05
东南大学学报(医学版)(2021年3期)2021-08-20
今日农业(2021年7期)2021-07-28
医学前沿(2021年7期)2021-07-27
煤气与热力(2021年5期)2021-07-22
医学理论与实践(2021年3期)2021-02-23
健康必读(上旬刊)(2020年8期)2020-08-13
