
纳米纤维素-铁螯合物矫治梨树缺铁黄化症的转录组分析
2022-01-26 10:00郭献平边艺伟王东升吴中营王合中连晓东
河南农业科学 2021年11期
郭献平,边艺伟,王东升,吴中营,王合中,连晓东,郭 鹏
(1. 河南省农业科学院园艺研究所,河南 郑州 450002;2. 河南农业大学植物保护学院,河南 郑州 450002;3. 河南农业大学园艺学院,河南 郑州 450002)
梨树在缺铁条件下易发生缺铁黄化病。发生缺铁黄化的原因较多,经常发生缺铁的土壤类型是碱性土壤,尤其是石灰性土壤。石灰性土壤pH 值一般在7.4~8.5,并具有高重碳酸盐[1]。石灰性土壤面积占世界陆地面积的30%以上[2]。我国的石灰性土壤分布很广,约占耕地面积的1/2[3]。西班牙西洋梨Blanquilla[4],美国华盛顿西洋梨Anjou[4],中国新疆库尔勒香梨[5],安徽省砀山酥梨[6]、鸭梨和黄金梨[6],湖北省湘南梨和棠梨[7],浙江省上虞市海涂地黄花梨[8],河北省泊头市鸭梨[9],陕西省蒲城县砀山酥梨[10],天津市秋白梨[11],四川省汉源县黄金梨[12]均有报道因缺铁导致的梨树黄化病。缺铁条件下,梨树叶片表现为缺铁黄化,同时梨树生长量、果实产量和品质均下降。据报道,库尔勒香梨黄化树单果质量比正常树降低29.3%,可溶性固形物含量下降15.2%,维生素C 含量降低41.4%,硬度和可滴定酸含量升高[13]。
叶面喷施铁肥是矫治缺铁黄化的有效途径之一[14]。铁肥的种类较多,常见的有无机铁肥、螯合铁肥与有机复合铁肥,如FeSO4·7H2O、Fe-EDTA(乙二胺四乙酸铁钠)、木质素磺酸铁等。无机铁肥FeSO4·7H2O 价格低,但其在空气中很快会被氧化成为三价铁沉淀物,导致喷施效果差;螯合铁肥效果好,但Fe-EDTA 在自然条件下EDTA 转化成乙二胺三乙酸,再经过环化,变成二酮哌嗪,成为持久性有机污染物并在环境中积累[15];有机复合铁木质素磺酸铁在环境中易降解,不存在环境污染的问题,但其含铁量低,性质不稳定。纳米材料由于具有独特的理化性质,如表面效应、小尺寸效应、量子尺寸效应、宏观量子尺寸效应以及介电限域效应[16],在生物医药、材料加工和能源环境等领域均有应用。纳米科技与农业科技的相互渗透,为农业科学发展提供了新的理论、方法和技术手段[17]。比如肥料方面,纳米几丁质促进N 代谢超过C 代谢,使冬小麦产量提高27.56%[18]。纳米二氧化钛利用TiO2的光化学效应,可增加种子的抗逆性[19]。从天然纤维素中提取的纳米纤维素(Cellulose nanocrystals,CNC),有资源丰富、可自然降解等优点,同时赋予它纳米粒子的特性,为其形成各种功能性复合材料提供了可能,具有广阔的应用前景[20]。如纳米纤维素与银螯合,CS-DCNC-AgNPs(壳聚糖-二醛纤维素纳米晶-银纳米粒)复合物具有良好的机械强度和疏水性,抗菌活性高,细胞毒性小,是一种有前途的、安全的抗菌药物[21]。本研究采用溶解级软木硫酸纸Temalfa 95A,通过水解法获得性能良好的纳米纤维素,将其与硫酸亚铁螯合形成新型可生物降解的铁螯合剂(纳米纤维素-铁螯合物,CNC-Fe);通过对杜梨缺铁黄化叶片喷施FeSO4和CNC-Fe 并进行相关生理指标和转录组测序分析,研究CNC-Fe 矫治梨树缺铁黄化症的效果及分子机制,为梨树抗黄化症机制等研究提供理论基础。
1 材料和方法
1.1 试验材料及处理
采用改良后的硫酸水解法制备纳米纤维素[22]。将Rayonier Advanced Materials 公司赠送的溶解级软木硫化纸(Temalfa 95A)粉碎后,取20 g 纤维素粉加入500 mL 三颈烧瓶,在水浴锅中预热至45 ℃。按1∶100 的反应比例向烧瓶中缓缓倒入200 mL 预热至45 ℃的64%硫酸;开启搅拌器,在45 ℃的恒温条件下反应45 min 后,用4 倍体积预冷的去离子水(4 ℃)终止反应。然后在4 ℃条件下9 000 r/min 离心15 min。弃上清,加入适量去离子水振荡润洗沉淀,倒入透析袋中,用去离子水透析10~15 d,除去游离酸。最后在冰浴中超声处理25 min,超声破碎功能设置为35%输出功率,工作5 s,间歇5 s,即可得到纳米纤维素水悬浮液。
杜梨幼苗长出6 片真叶后,选择高度均一的幼苗移至水培槽培养杜梨黄化苗,水培槽中营养液为1×10-8mol/L EDTA-Fe 改良的Hogland 营养液,pH 值约6.5[23]。杜梨幼苗长至25 cm左右时移至人工气候箱,每盆3 株黄化杜梨苗,设定16 h 光照(26 ℃)、8 h黑夜(18 ℃)。选择SPAD值10~30的叶片做标记。
试验设计3 个处理,处理1(T1)喷施4 mmol/L FeSO4,处理2(T2)为纳米纤维素与FeSO4按照电荷比1∶3 000螯合后喷施,每个处理均添加0.15%吐温80,对照(CK)喷施去离子水,每个处理重复3 次。处理72 h 后测定相关生理指标,并取叶片转至液氮速冻,置于-80 ℃冷冻保存备用。
1.2 纳米纤维素粒径、Zeta电位和综合电荷密度的测定
利用动态光散射法,用马尔文纳米粒度仪(Nanosizer,ZS 90)测定纳米纤维素的流体动力学粒径和Zeta电位。使用雷磁电导率仪(DDS-307)通过电导滴定法测定纳米纤维素的磺酸基综合电荷密度[24]。
1.3 叶片活性铁、叶绿素相对含量(SPAD值)和净光合速率的测定
活性铁含量采用酸消解ICP 法测定[25‐26]。叶片在105 ℃杀青30 min,然后在70 ℃条件下烘干7 d至恒定质量后,称取干燥后叶片0.1 g,加入10 mL 1.0 mol/L 盐酸浸提,连续振荡24 h,过滤后用ICP 法测定浸提液中铁含量。叶绿素相对含量利用spad-502 便携式叶绿素测定仪测定。采用汉莎光合作用测定仪(CIRAS-3)测定净光合速率。
1.4 转录组测序
转录组测序所用材料为3 种处理72 h 后的叶片,3次生物学重复。RNA 提取采用总RNA 提取试剂盒,RNA 质量检测使用Qubit,保证RNA 质量满足建库使用。测序工作委托苏州金唯智生物科技有限公司基于Illumina HiSeqTM 2500 测序平台完成,采用Hisat2(V2.0.1)软件进行序列比对,Htseq 软件(V0.6.1)计算基因FPKM(Fragments per kilo bases per million reads)值[27]。铁蛋白家族基因表达量采用Z-score归一法处理。
1.5 差异表达基因筛选与功能注释
利用DESeq2(V1.6.3)进行差异表达基因分析,以差异基因表达变化2 倍以上,且Q-vaule≤0.05 进行筛选。将获得的差异基因进行GO(Gene ontology)和KEGG(Kyoto encyclopedia of genes and genomes)注释和富集分析。果胶甲酯酶(PME)和果胶甲酯酶抑制子(PMEI)结构域利用SMART(http://smart.embl‐heidelberg.de/)分析验证。
1.6 实时荧光定量PCR(qRT-PCR)验证
对参与KEGG 通路的8 个差异表达基因(Differentially expressed genes,DEGs)表达模式通过实时荧光定量PCR 进一步分析。采用qRT-PCR 内参基因归一化2-ΔΔCt法进行转录组数据的验证,3 个生物学重复。使用在线软件Primer Premier 5设计引物,梨Actin基因作为内参,引物序列见表1。

表1 实时荧光定量PCR引物Tab.1 qRT-PCR primers
1.7 数据统计与分析
利用SPSS 22.0 进行统计分析,采用Excel 和Origin 8.5进行数据处理和作图。
2 结果与分析
2.1 纳米纤维素的表征
经马尔文纳米粒度仪测定,纳米纤维素的流体动力学粒径和Zeta 电位分别为(84.3±0.1)nm 和(-47.30±1.0)mV,说明纳米纤维素带负电荷(图1)。通过电导滴定法检测纳米纤维素水悬浮液的综合电荷密度,纳米纤维素综合电荷密度的电导率滴定曲线为典型的V 形滴定曲线,与文献[22‐24]中的结果一致,其综合表面带电荷量为(108.5±3.1)mmol/kg(图2)。
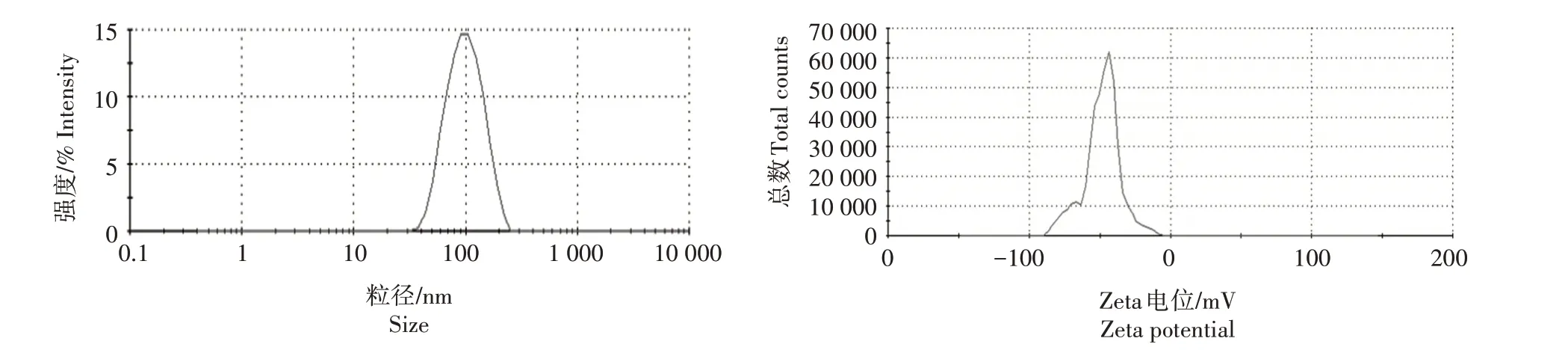
图1 纳米纤维素粒径和Zeta电位Fig.1 Particle size and Zeta potential of CNC
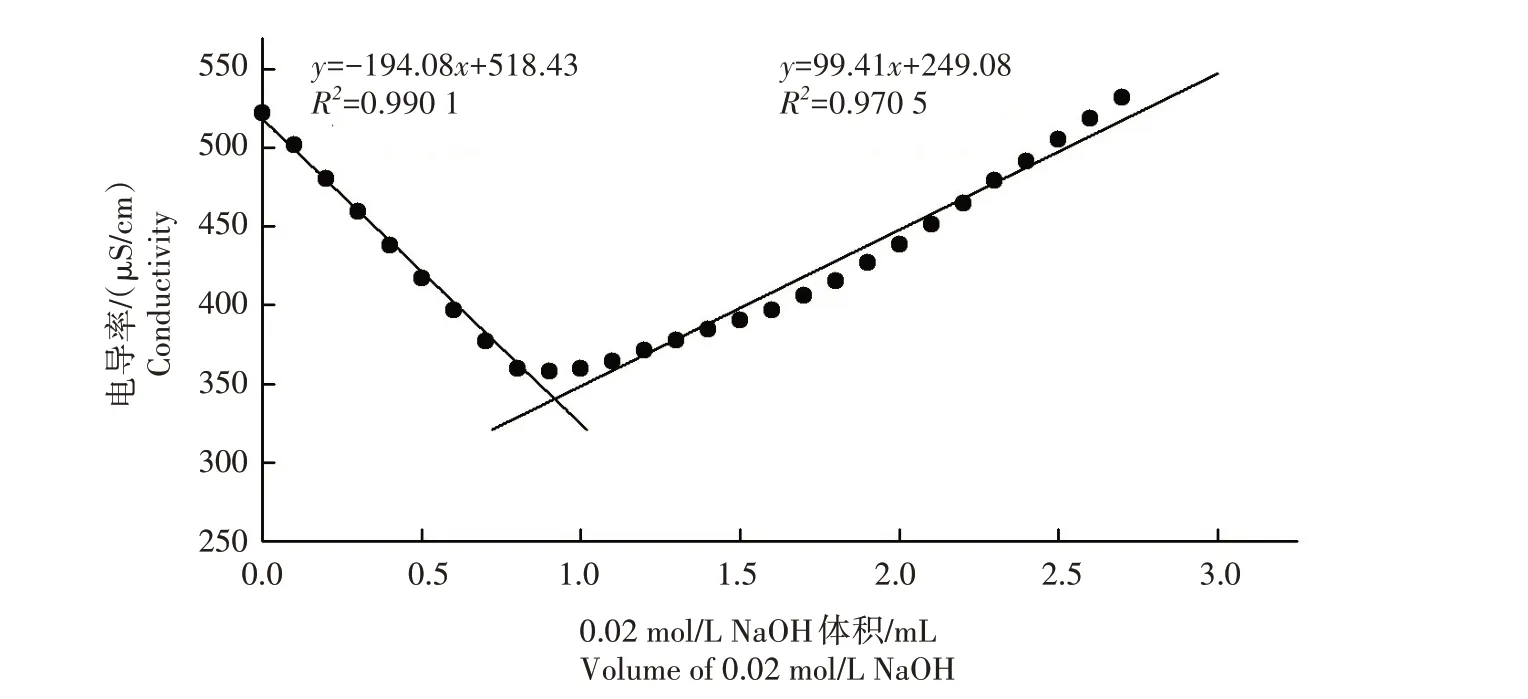
图2 纳米纤维素综合电荷密度的电导率滴定曲线Fig.2 Conductometric titration curves for the determination of integrated charge density of CNC
2.2 不同处理对杜梨叶片活性铁含量、SPAD值和净光合速率的影响
因FeSO4易被氧化成为三价铁沉淀物降低复绿效果[28],处理72 h 后,T1(喷施FeSO4)部分复绿,而T2(喷施CNC-Fe)基本复绿(图3)。“铁素黄化悖论”说明缺铁黄化叶片的全铁含量与叶绿素含量相关性极低,而叶片活性铁与叶绿素含量之间则呈现高度正相关[29‐30]。由表2 可知,T1 与T2 均提高叶片的活性铁含量,T1 和T2 叶片活性铁含量分别比CK 提高110.7%和235.8%,差异均显著。T2 叶片活性铁含量比T1显著提高59.4%。

图3 不同处理72 h后杜梨叶片表型Fig.3 Phenotypes of Pyrus betulifolia leaves after 72 h of different treatments
SPAD 值能够反映叶片黄化程度,因与叶绿素含量有较好的相关性,被广泛用于评价作物缺铁状况[31]。由表2 可知,T1 和T2 不同程度地提高杜梨叶片的SPAD 值,T1、T2 的SPAD 值比CK 分别提高26.1%和61.7%,差异均显著;且T2 比T1 的SPAD 值显著提高28.2%。T1、T2 净光合速率比CK 分别提高70.1%、98.5%,差异均显著。
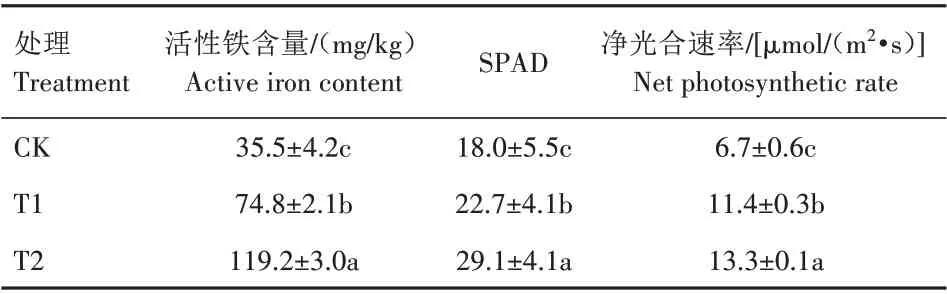
表2 不同处理72 h后杜梨叶片活性铁含量、SPAD值和净光合速率Tab.2 Active iron content,SPAD value and net photosynthetic rate of Pyrus betulifolia leaves after 72 h of different treatments
综上可知,喷施CNC-Fe比单独喷施FeSO4更能提高黄化叶片的活性铁和叶绿素含量以及光合能力,说明CNC-Fe矫治缺铁黄化症的能力更强。
2.3 测序数据质量分析
由表3 可知,利用Illumina HiSeqTM 2500 测序平台分别对T1、T2 和CK 叶片进行转录组测序。测序结果显示,叶片GC 含量均大于48%,Q20、Q30 值都高于91%,表明转录组测序的质量较高,可以进行下一步分析。经聚类图分析,表明每组重复性较好(图4)。
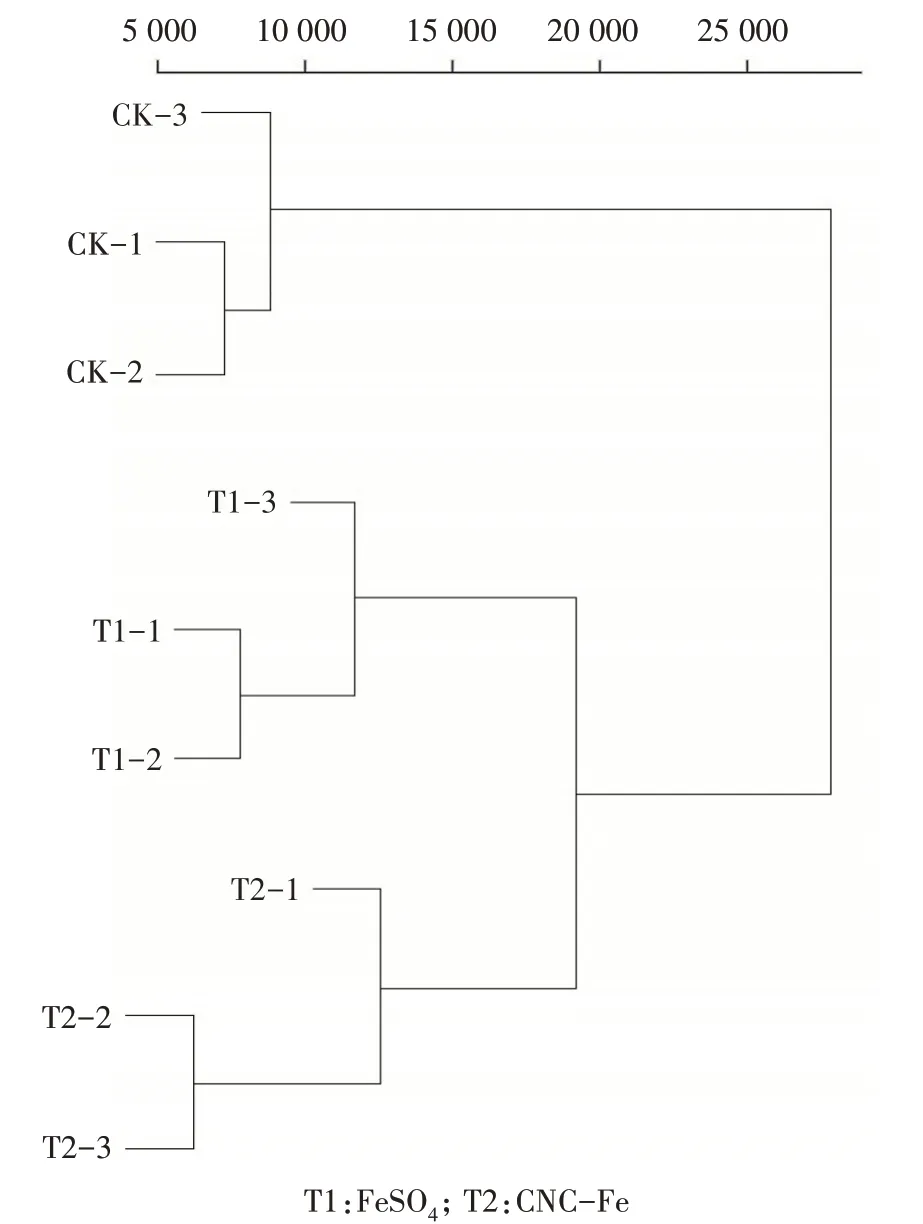
图4 不同处理杜梨叶片样品间基因表达量聚类Fig.4 Cluster diagram of gene expression among Pyrus betulifolia leaves samples of different treatments
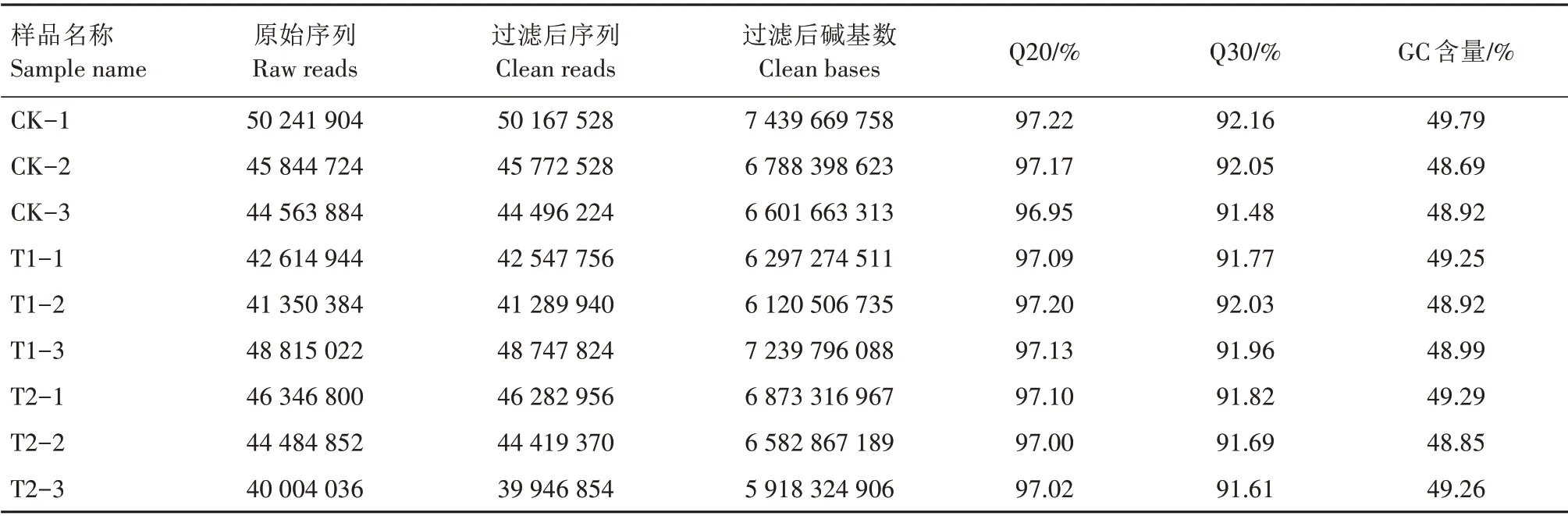
表3 转录组测序数据Tab.3 Data assembly for transcriptome sequencing
2.4 基因表达水平分析
由图5 可知,与CK 相比,T1、T2 分别有1 033、1 943 个差异基因。T1 上调基因527 个,下调基因506 个;T2 上调基因861 个,下调基因1 082 个。不同处理间相比,T1较T2有650个上调基因,329个下调基因。通过韦恩图分析,T1、T2 与CK 相比,分别特有DEGs 为586、1 496 个,说明T1 和T2 对矫治缺铁黄化症的调控方式存在较大差异。
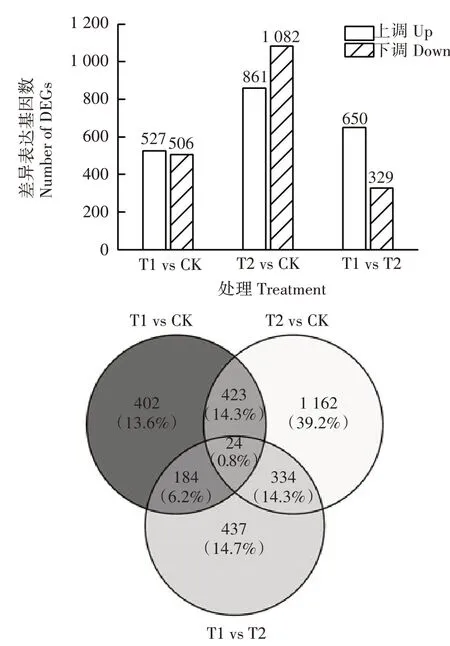
图5 不同处理杜梨叶片中上调和下调DEGs的数量与DEGs韦恩图Fig.5 Numbers of up-regulated and down-regulated genes and Venn diagram of DEGs in Pyrus betulifolia leaves under different treatments
2.5 差异表达基因GO功能富集分析
通过差异表达基因GO 功能富集分析,T2 比T1多26 个GO 项。由图6 可知,GO 功能富集分析提取了分子功能(Molecular function,红色部分)、细胞组分(Cellular component,蓝色部分)和生物过程(Biological process,绿色部分)前10 个GO 项。与CK 相比,T1 和T2 中共有GO 通路包括GO∶0046872金属离子固定(Metal ion binding)、GO∶0016491 氧化还原酶活性(Oxidoreductase activity)、GO:0016020膜(Membrane)、GO∶0009507 叶绿体(Chloroplast)、GO∶0055114 氧化还原过程(Oxidation‐reduction process)和 GO∶0009765 光 合 捕 光 过 程(Photosynthesis,light harvesting)。其中叶绿体和光合捕光过程的差异表达基因均上调,说明T1 和T2可能通过调节金属离子固定、氧化还原酶、膜、叶绿体和光合捕光系统等相关基因的活性,提高活性铁、叶绿素含量和光合能力。
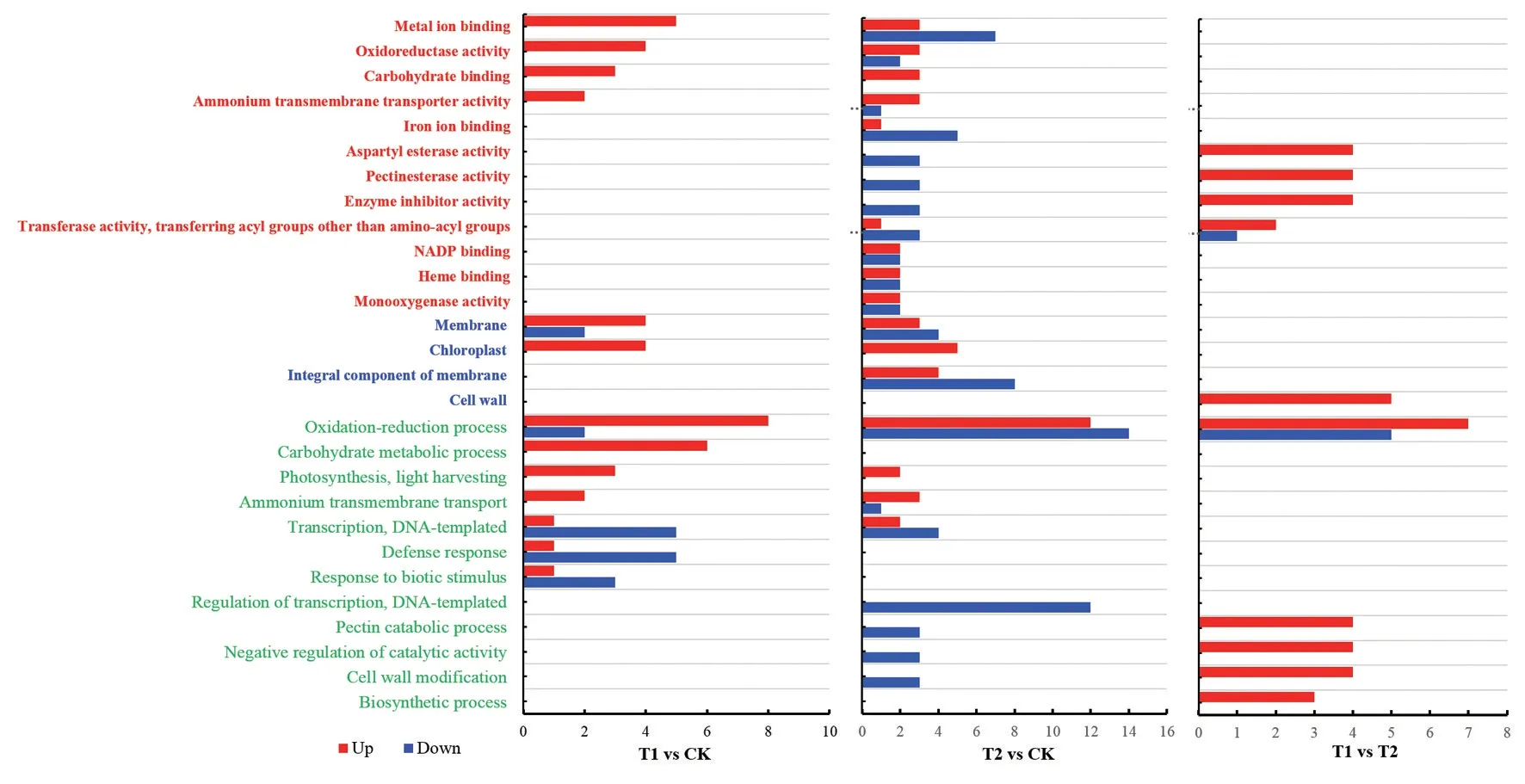
图6 不同处理杜梨叶片差异基因的GO富集分析Fig.6 GO enriched analysis of differentially expressed genes in Pyrus betulifolia leaves under different treatments
与CK 相比,T2 特有的GO 通路有GO∶0005506铁离子固定(Iron ion binding)、GO∶0030599 果胶甲酯酶活性(Pectinesterase activity)、GO∶0045490 果胶分解代谢过程(Pectin catabolic process)和GO∶0042545 细胞壁修饰(Cell wall modification)。在T1 vs T2 中,GO 富集的主要通路除了包含果胶甲酯酶活性、果胶分解代谢过程、细胞壁修饰,还包括GO:0005618细胞壁(Cell wall),经分析这些GO项差异基因为PME4(Pectin methyl‐esterase 4)、LOC103943418(Probable pectinesterase/pectinesterase inhibitor 34)、LOC103962785(Probable pectinesterase/pectinesterase inhibitor 51)、LOC103948035(Probable pectinesterase/pectinesterase inhibitor 34),均为果胶甲酯酶,CK、T1 和T2 的PME4表达量分别为81.57、55.27、8.75,LOC103943418表达量为47.77、34.44、10.30,LOC103962785表达量为0.31、0.50、0.01,LOC103948035表达量为111.23、123.79、43.41,PME4和LOC103943418表达量由高到低依次均为CK、T1 和T2,LOC103962785和LOC103948035表达量由高到低依次均为T1、CK 和T2,喷施CNC-Fe螯合物的叶片72 h 后果胶甲酯酶表达量均低于喷施FeSO4和CK。
2.6 差异表达基因KEGG富集分析
KEGG 富集分析结果(图7)显示,与CK 相比,T1 处理中仅有55 个差异表达基因注释到4 个通路中;而T2 处理中有712 个差异基因注释到18 条通路,说明T2能够调动更多的通路促进叶片复绿。其中光合作用-天线蛋白(Photosynthesis‐antenna proteins)和光合作用(Photosynthesis)2个通路,在T1和T2 处理中都呈显著富集。T1 中光合作用-天线蛋白通路共有8 个差异表达基因,全部为上调;T2中注释到17 个差异表达基因,也全部为上调。T1中光合作用通路注释到9 个差异表达基因(8 个上调,1个下调);在T2中光合作用通路注释到41个差异表达基因(40 个上调,1 个下调)。此外,与CK 相比,T1 中植物病原互作(Plant‐pathogen interaction)和脂肪酸伸长(Fatty acid elongation)也为富集通路;T2 中代谢途径(Metabolic pathways)、次生代谢产物生物合成(Biosynthesis of secondary metabolites)、碳代谢(Carbon metabolism)和光合组织的碳固定(Carbon fixation in photosynthetic organisms)等呈现显著富集。T1 与T2 相比,木栓质和蜡的生物合成(Cutin,suberine and wax biosynthesis)、脂肪酸生物合成(Fatty acid biosynthesis)、脂肪酸代谢(Fatty acid metabolism)、甘油磷脂代谢(Glycerophospholipid metabolism)、硫代谢(Sulfur metabolism)、Ras 信号通路(Ras signaling pathway)等共11 个通路呈现显著富集。
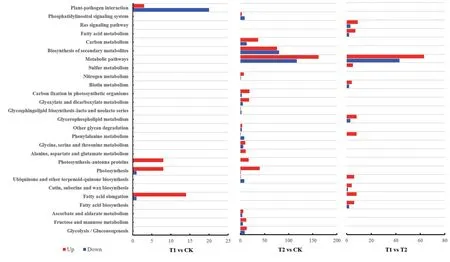
图7 不同处理杜梨叶片差异基因的KEGG显著富集分析Fig.7 KEGG analysis of differentially expressed genes in Pyrus betulifolia leaves under different treatments
2.7 铁蛋白(Ferritin)家族基因表达趋势
梨基因组中铁蛋白家族共有4 个基因,分别为Fer1(LOC103930157)、Fer2(LOC103958648)、Fer3(LOC103965102)和Fer4(LOC103937157)。通过转录组中铁蛋白家族基因热图分析(图8)可知,T2的4种铁蛋白基因表达量均高于T1,除CKFer2表达量高于T1 和T2 外,T1 和T2Fer1、Fer3、Fer4表达量均高于CK,说明T2叶片铁离子贮存能力强于T1。
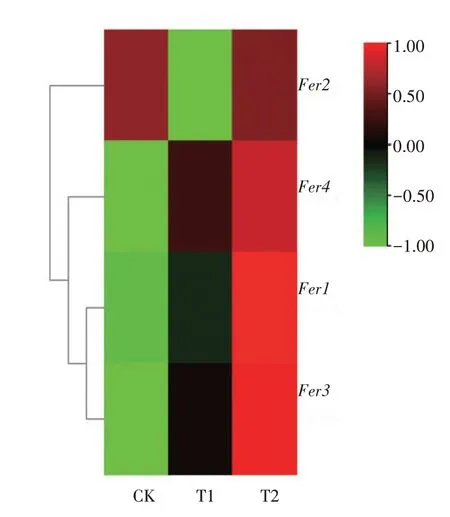
图8 不同处理杜梨叶片铁蛋白家族基因热图Fig.8 Heat map of ferritin family gene in Pyrus betulifolia leaves under different treatments
2.8 光合作用相关的差异表达基因
与CK 相比,T1 和T2 共有的通路是光合作用-天线蛋白,有58 个基因上调表达,其中56 个基因表现出T2中表达量最高。在光合作用-天线蛋白通路中(图9),编码光合作用-天线蛋白LHCⅠ基因家族有5 类基因,与CK 相比,T1 和T2 有3 类5 个基因表达上调,分别为Lhca1(编码捕光复合体Ⅰ叶绿素a/b-1 结合蛋白)、Lhca2(编码捕光复合体Ⅰ叶绿素a/b-2 结合蛋白)、Lhca4(编码捕光复合体Ⅰ叶绿素a/b-4 结合蛋白);编码LHCⅡ蛋白基因家族有7 类基因,与CK 相比,T1、T2 有6 类13 个基因呈现上调表达,分别为Lhcb1(编码捕光复合体Ⅱ叶绿素a/b-1 结合蛋白)、Lhcb2(编码捕光复合体Ⅱ叶绿素a/b-2 结合蛋白)、Lhcb3、Lhcb4、Lhcb5、Lhcb6。在以上所有上调表达基因中,T2 表达量均高于T1,说明CNC-Fe能更加有效地向反应中心输送能量。
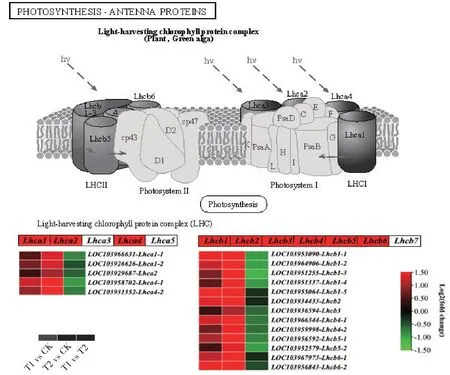
图9 参与光合作用-天线蛋白通路的DEGs表达情况Fig.9 Summary of DEGs in the photosynthesis-antenna proteins pathway
光合作用通路中编码PSⅡ系统相关的Psb基因家族有27 类基因(图10),与CK 相比,T1 和T2 有7 类14 个基因呈现上调表达,分别为PsbO(编码光系统Ⅱ-1 放氧增强蛋白)、PsbP(编码光系统Ⅱ-2放氧增强蛋白)、PsbQ(编码光系统Ⅱ-3 放氧增强蛋白)、PsbR(编码光系统Ⅱ10kDa 蛋白)、PsbW(编码光系统ⅡPsbW 蛋白)、PsbY(编码光系统ⅡPsbY蛋白)、Psb27(编码光系统ⅡPsb27 蛋白)。编码PSⅠ系统相关的Psa基因家族有16类基因,这些亚基蛋白主要参与PSⅠ反应中心的组成,与CK 相比,T1、T2 有8 类15 个基因呈现上调表达,分别为PsaD(编码光系统Ⅰ-Ⅱ亚基)、PsaE(编码光系统Ⅰ-Ⅳ亚基)、PsaF(编码光系统Ⅰ-Ⅲ亚基)、PsaG(编码光系统Ⅰ-Ⅴ亚基)、PsaH(编码光系统Ⅰ-Ⅵ亚基)、PsaK(编码光系统Ⅰ-Ⅹ亚基)、PsaN(编码光系统Ⅰ-PsaN 亚基)、PsaO(编码光系统Ⅰ-PsaO 亚基);编码细胞色素b6/f复合物和光合电子传输过程的相关蛋白Pet基因家族有12类基因,有3类6个基因上调表达,分别为PetC(编码细胞色素b6-f 复合铁硫亚基)、PetE(编码质体蓝蛋白)、PetF(编码铁氧还蛋白),PetH(编码铁氧还蛋白-NADP+还原酶)下调表达。编码F 型ATP 酶相关蛋白有8 类基因,其中有3 类5 个基因上调表达,分别为gamma(F 型H+转运ATPase-gamma 亚基)、delta(F 型H+转运ATPasedelta亚基)、b(F型H+转运ATPase-b 亚基)。在以上40 个上调表达基因中,有38 个基因T2 表达量高于T1,说明CNC-Fe能更有效促进杜梨的光反应效率。
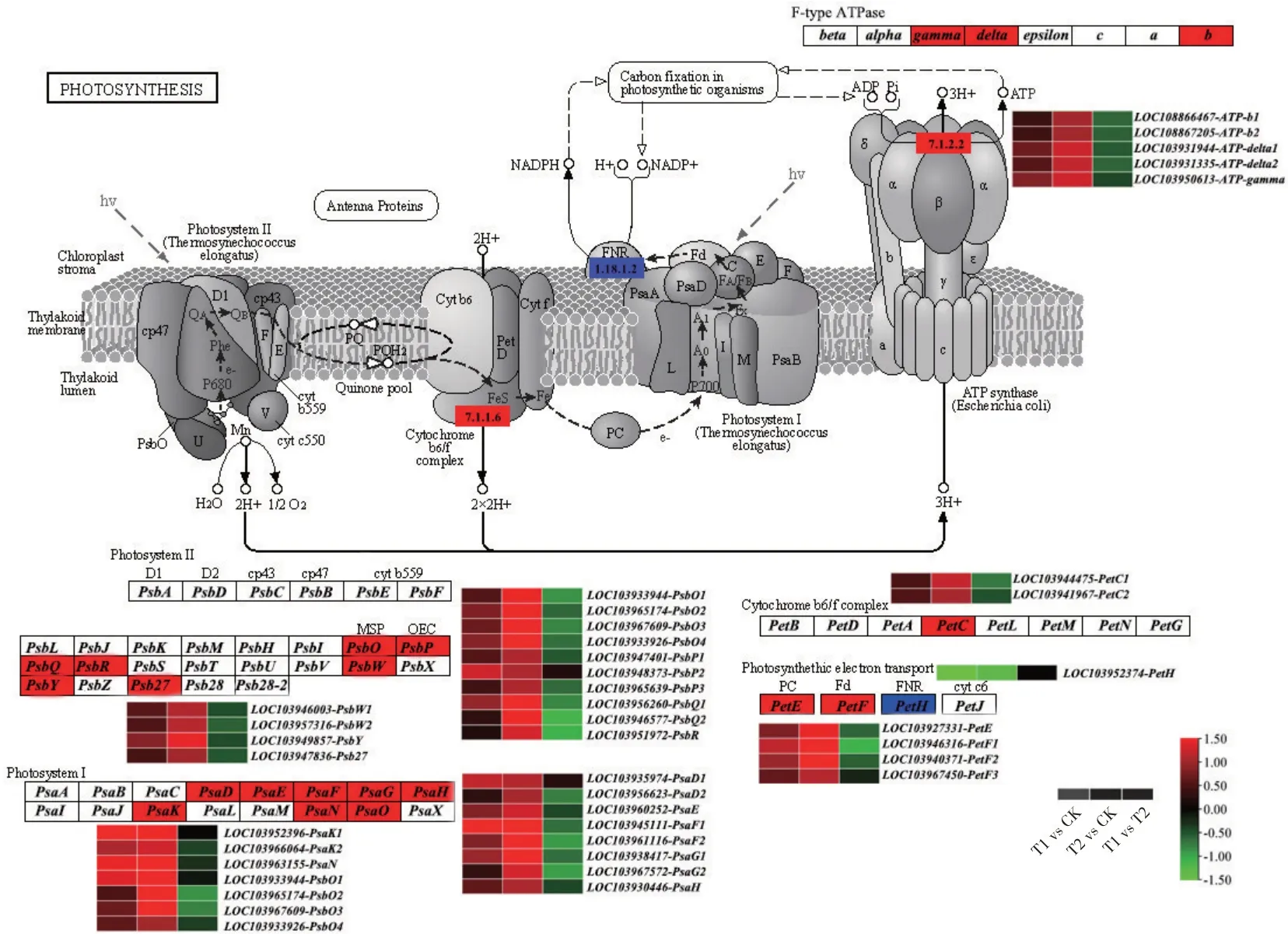
图10 参与光合作用通路的DEGs表达情况Fig.10 Summary of DEGs in the photosynthesis pathway
2.9 相关差异表达基因的定量验证
对参与光合作用中PSⅡ、PSⅠ、细胞色素b6/f复合物、光合电子传输的相关8 个DEGs 进行qRTPCR 分析,结果显示,与RNA-seq 分析的表达模式类似(图11)。参与PS Ⅱ的4 个DEGs 分别为LOC103946003、LOC103967609、LOC103948373、LOC103949857, 参 与 PS Ⅰ 的 DEGs 为LOC103945111,参与细胞色素b6/f 复合物的DEGs为LOC103944475,参与光合电子传输的2 个DEGs分别为LOC103927331、LOC103946316。
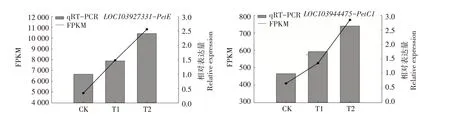
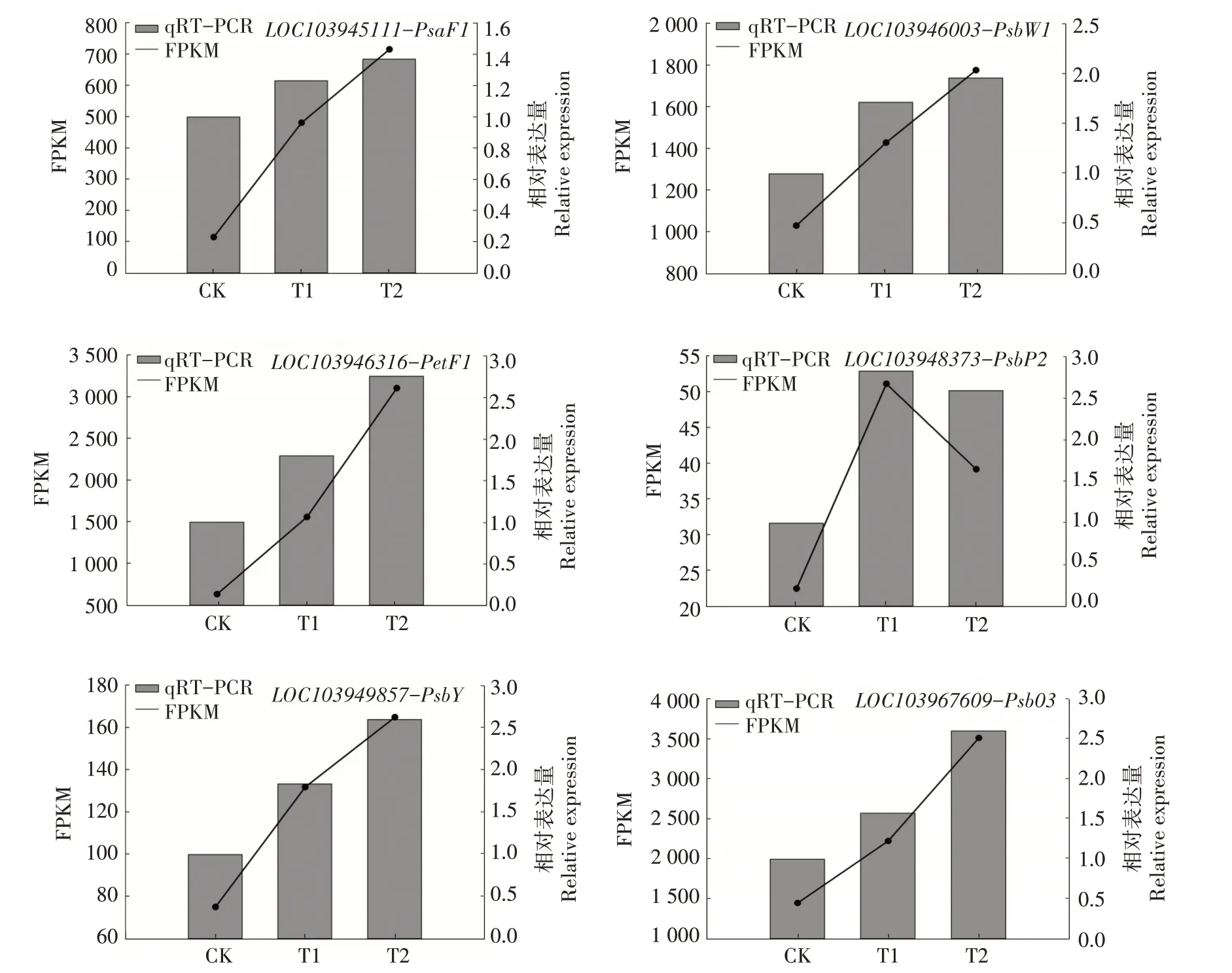
图11 候选基因的实时荧光定量PCR验证Fig.11 Quantitative real-time PCR validation of candidate genes
3 结论与讨论
3.1 喷施CNC-Fe比FeSO4能调动更多的基因和代谢通路,促进杜梨黄化叶片复绿
铁是植物中生长的必需微量元素,参与了多种代谢途径,比如叶绿素生物合成、光合作用、呼吸作用,另外还作为辅助因子参与电子传递[32]。处理杜梨72 h 后,通过对其叶片进行转录组测序与分析,解释了铁素代谢、光合作用等相关基因转录表达情况。与CK相比,T2比T1上调表达基因329个,下调表达基因650个;T2比T1 GO富集多26项,KEGG富集多14 个代谢通路。说明CNC-Fe 能够刺激更多基因和代谢通路促进杜梨黄化叶片复绿。
3.2 CNC-Fe比FeSO4能改善铁素代谢相关基因表达,提高杜梨叶片的活性铁含量
植物铁蛋白是存在于植物细胞质体中的一种专门储存铁的蛋白质,约80%的铁存在于质体内的铁蛋白中[33]。植物铁蛋白由24 个亚单位组成,在其结构的空腔中可储存4 500 个铁原子[34]。本研究T1和T2 处理杜梨叶片活性铁含量比CK 分别提高110.7%和235.8%,差异均显著;T2 叶片活性铁含量比T1 显著提高59.4%。T2 的4 种铁蛋白基因表达量均高于T1,4 种铁蛋白基因的增幅各不相同。贾兵等[35]在外源喷施0.2%FeSO4处理缺铁黄化砀山酥梨叶片研究中,处理8 d 和12 d 后Fer1、Fer2、Fer3、Fer4相对表达量和活性铁含量显著高于清水处理黄化叶片。在砀山酥梨轻度、中度和重度缺铁条件下4种铁蛋白基因表达量分析中,与正常叶片相比,PbFer1、PbFer3和PbFer4的表达量均呈现随着缺铁程度的增加表达量逐渐降低,而PbFer2在轻度缺铁条件下的表达量高于正常叶片,中度和重度条件下逐渐降低[36]。本研究结果表明,Fer1、Fer3基因表达量更能体现叶片贮存铁的能力。
果胶是植物细胞壁的主要成分之一,并且是阳离子的主要结合位点。果胶甲酯酶普遍存在于高等植物的不同组织器官,如根、茎、叶、果实等,其对细胞壁组成和降解、根尖延伸、抗病等方面具有重要作用[37‐38]。缺铁条件下,植物根系果胶甲酯酶活性增强,使果胶产生较多的游离羧基即阳离子吸附位点,增加与铁的结合能力[39]。植物果胶甲酯酶抑制子与果胶甲酯酶结合形成1∶1的可逆复合物抑制其活性[40]。本研究转录组分析结果表明,T2 处理的叶片果胶甲酯酶表达量均低于T1 和CK,T1 处理PME4的表达量为T2 的6.3 倍,CKPME4的表达量为T2 处理的9.3 倍,说明CNC-Fe 可能通过抑制果胶甲酯酶表达量而降低果胶甲酯酶活性,减少质外体对铁离子的固定,使更多的铁通过铁素相关代谢转运至质体内,最终提高叶片活性铁含量使叶片复绿。
3.3 CNC-Fe比FeSO4更能提高叶绿素含量和光反应效率,增强杜梨叶片的光合能力
本研究中,T1 和T2 处理杜梨叶片SPAD 值分别比CK 显著提高26.1%和61.7%,净光合速率显著提高70.1%和98.5%。转录组分析光合作用-天线蛋白和光合作用2 个通路中,T1 和T2 处理共有58 个基因上调,其中T2处理有56个基因表达量高于T1。以上说明,喷施CNC-Fe比FeSO4更能提高叶绿素含量和光反应效率,增强杜梨叶片的光合能力。
综上可知,喷施CNC-Fe 能调动更多的基因和代谢通路,使其矫治梨树缺铁黄化症的能力强于FeSO4。
猜你喜欢
中国果业信息(2021年10期)2021-12-07
纺织科技进展(2021年4期)2021-07-22
纺织科技进展(2021年3期)2021-06-09
纺织科技进展(2021年3期)2021-06-09
疯狂英语·读写版(2020年11期)2020-12-21
安徽农学通报(2017年18期)2017-10-18
农业知识·百姓新生活(2014年5期)2015-01-20
热带农业科学(2014年6期)2014-08-07
食品与健康(2014年4期)2014-06-06
意林(2009年16期)2009-05-14
