
三峡水库光泽黄颡鱼年龄、生长与死亡特征研究
2022-01-20 08:09廖传松叶少文张堂林刘家寿
水生生物学报 2022年1期
廖传松 李 为 苑 晶 叶少文 张堂林 刘家寿
(中国科学院水生生物研究所, 淡水生态与生物技术国家重点实验室, 武汉430072)
鱼类的年龄、生长与死亡等种群特征研究是开展渔业资源评估与管理的基础[1], 这些种群特征受遗传基础和自然环境的共同影响[2,3]。生活史理论认为种群的生活史策略是成熟年龄、幼体存活率及繁殖力三个维度间权衡的结果[3]。尽管受到遗传变异和进化史的限制, 面对不同的环境条件, 生物体的能量分配会在生长、存活及繁殖间进行权衡[4,5]。即使是同种鱼类, 在不同环境条件下形成的种群特征也会有所差异[4,6], 食物可得性、生境特征、捕捞压力及种间竞争等都可能影响种群特征[7, 8]。
光泽黄颡鱼(Pelteobagrus nitidus), 隶属于鲇形目(Siluriformes), 鲿科(Bagridae)[9], 是一种中小型底层鱼类, 主要分布于长江流域的干支流和湖泊, 具有较高的营养价值。已有学者对长江中下游一些水体的光泽黄颡鱼种群特征进行了研究, 例如, 袁刚等[10]研究了洞庭湖光泽黄颡鱼种群的食性, 发现水生昆虫幼虫和甲壳类是其主要食物, 但存在明显的季节变化, 这与刘其根等[11]在淀山湖的研究结果相似; 耿龙等[12]对淀山湖光泽黄颡鱼种群的生长、繁殖和摄食特征进行了初步研究。这些研究结果表明光泽黄颡鱼的种群生物学特征在不同的环境中具有一定的可塑性。
三峡水库位于长江上游, 是世界上已建成最大的水库之一。分布在库区的黄颡鱼属鱼类共有4种[13],三峡水库蓄水前黄颡鱼属鱼类在群落中所占比例为7.2%[14]。三峡水库蓄水改变了原有河流生境, 黄颡鱼属鱼类所占比例较蓄水前有所升高[14—17], 已成为库首和库中的优势类群[18]。其中, 光泽黄颡鱼分布于整个库区, 其生物量在鱼类群落中占比1.00%—6.79%[16,17]。由于三峡水库蓄水后水深增大、流速变缓和底泥沉积等因素, 作为光泽黄颡鱼重要饵料的底栖动物资源受到明显影响[19,20]。已有研究关注了三峡水库光泽黄颡鱼的繁殖和摄食特征[21,22],但关于光泽黄颡鱼的年龄和生长等种群特征尚不明晰。
本研究选择三峡水库库首、库中和库尾3个不同江段为研究地点, 研究光泽黄颡鱼的年龄结构、生长方程、死亡系数和肥满度等种群特征及其在不同生境之间的变化, 旨在为三峡水库鱼类资源的科学管理和保护提供基础资料。
1 材料与方法
1.1 采样点设置
选取三峡水库秭归、万州和木洞3个江段作为研究地点, 这3个江段具有不同的生境特征和底栖动物资源。其中, 秭归(30°51′36.6′′ N, 110°59′51.9′′E)紧邻三峡大坝, 底栖动物密度为243 ind./m2; 万州(30°49′46.9′′ N, 108°25′03.8′′ E)位于库中, 距离大坝约300 km, 年平均流速为0.26 m/s, 底栖动物密度为184 ind./m2; 木洞(29°34′51.4′′ N, 106°51′01.2′′ E)位于库尾, 距离大坝约600 km, 年平均流速为1.28 m/s,底栖动物密度为8180 ind./m2[20,23](图 1)。
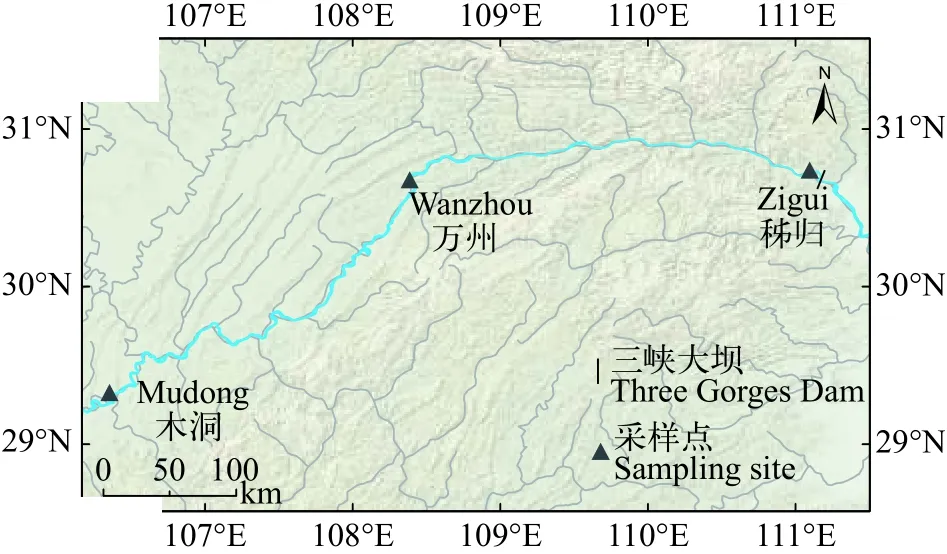
图 1 三峡水库采样点示意图Fig. 1 Map of sampling sites in the Three Gorges Reservoir
1.2 样品采集与处理
2015年1月至2016年10月, 按季度在3个江段采集光泽黄颡鱼样本, 秭归和万州的样本均采自刺网和地笼的渔获物, 木洞的样本采自刺网、地笼和拖网的渔获物, 在每个江段随机抽取鱼类样本, 在3个江段共采集鱼类2477尾。在新鲜状态下, 测量每尾样本的全长(TL, 精确到0.1 mm)、体长(SL)和体重(BW, 精确到0.1 g)。对部分样本进行解剖, 鉴定性别(雌、雄和性别不明), 剥离附着于内脏表面的脂肪并称重(脂肪重), 分离肝脏并称重(肝脏重), 然后称量去内脏后的空壳重。取出左右微耳石作为鱼类年龄的鉴定材料, 去除附带组织并用清水洗净,自然晾干后编号保存。
耳石的处理在实验室内进行, 采用中性树脂将耳石固定在载玻片上, 竖立放置, 先后采用400、1000和2000目的水砂纸打磨, 当打磨至耳石中心时,用抛光纸抛光。接着将耳石翻面, 用相同方法固定、打磨和抛光, 直至轮纹清晰, 保存在载玻片盒中[24]。在显微镜下鉴定年龄, 年龄的鉴定和计数参照殷名称[2]描述的方法, 并测量耳石轮径 (Otolith radius, OR)[2], 本研究共鉴定360尾鱼类的耳石样品。采集鱼类样本期间, 采用HOBO温度记录仪记录水下1 m处水温, 频率为每3h记录一次。
1.3 数据分析
Kruskal-Wallis检验和t检验分别用于比较不同江段、不同性别间光泽黄颡鱼的平均体长的差异。卡方检验(Chi-Square tests)用于年龄结构的空间比较。构建鱼类体长与耳石轮径的回归关系, 协方差分析(ANCOVA)用以检验鱼类体长与耳石轮径的关系在江段间的差异。
构建光泽黄颡鱼体长与体重关系(BW=aSLb),采用t检验比较雌、雄个体的b值与3的差异, 判断鱼类的生长式型[25]。采用von Bertalanffy (VBGF)生长方程拟合其体长生长方程和体重生长方程其中,Lt和Wt分别表示t龄鱼的体长和体重,L∞和W∞分别表示渐进体长和渐进体重,K表示生长曲线的生长系数,t0为表示理论上Lt= 0时的年龄[26]。基于体长-年龄数据, 采用FiSAT软件(FAO-ICLARM Fish Stock Assessment Tools, Version 1.2.2)拟合L∞和K[27], 以样本中的最大体长作为模型中L∞估计值的初始值。t0采用Pauly[28]提出的公式计算, 总死亡系数(Z)采用Beverton和Holt[29]提出的公式计算, 自然死亡系数(M)与L∞、K与平均水温T相关, 采用Pauly[30]提出的公式计算。捕捞死亡系数(F)为总死亡系数(Z)与自然死亡系数之差, 种群开发率E由公式E=F/Z计算。参照詹炳义[1]介绍的方法构建光泽黄颡鱼的体长、体重的生长速度和生长加速度方程, 并评估其体重生长拐点年龄和拐点体重。
计算每个个体的肥满度 (Fullness index,Kr), 计算方法为Kr=100WE/(SL/10)3, 并计算肝体系数(Hepato-somatic index,HSI)和脂体系数(Lipid-somatic index,LSI), 分别表征为肝脏重和脂肪重与空壳重的比值。一般线性模型分别用于比较肥满度、肝体系数和脂体系数在不同江段之间的差异。本研究采用R 4.0.2分析数据和作图, 以P=0.05为显著性水平。
2 结果
2.1 体长分布与年龄结构
光泽黄颡鱼在库区的平均体长为(87.1±0.5) mm,在秭归、万州和木洞的平均体长分别为(80.1±0.9) mm(N=1044)、(90.6±0.9) mm (N=571)和(93.1±0.7) mm(N=862; Kruskal-Wallis test,H=138,P<0.01; 图 2),平均体重分别为(8.6±0.3)、(10.1±0.2)和(12.5±0.3) g(H=192.82,P<0.01)。所能鉴别性别的个体中, 雌、雄个体的平均体长无显著差异, 分别为(94.4±0.7) mm(N=1126)和(94.9±0.7) mm (N=995;t-test,t=-0.58,P=0.56), 性别不明个体的平均体长为(60.9±2.6) mm(N=356)。
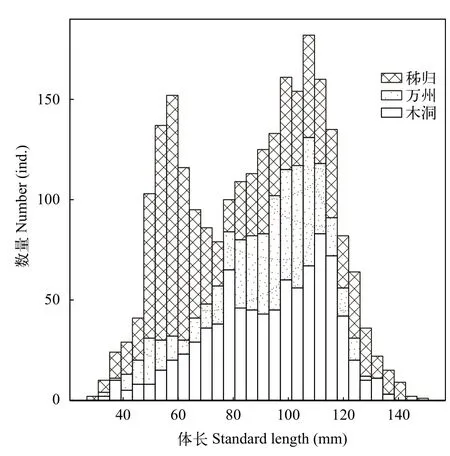
图 2 三峡水库不同江段泽黄颡鱼的体长分布Fig. 2 Standard length distributions of P. nitidus in different sections of the Three Gorges Reservoir
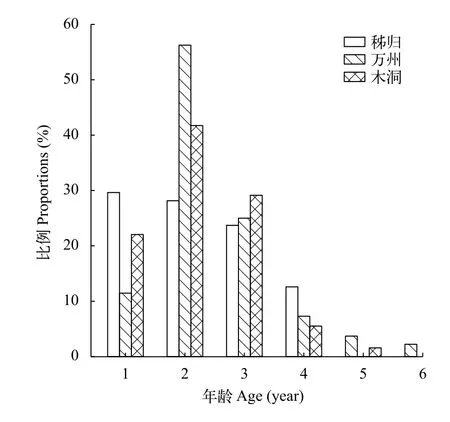
图 3 三峡水库不同江段光泽黄颡鱼的年龄结构Fig. 3 Age structures of P. nitidus in different sections of the Three Gorges Reservoir
光泽黄颡鱼的年龄结构由1—6龄组成, 以2龄为优势年龄组, 占42.04% (图 3)。秭归种群的最大年龄为6龄, 以1龄、2龄和3龄为优势年龄组, 分别占29.63%、28.15%和23.70%; 万州种群最大年龄为4龄, 以2龄和3龄占优势, 分别占56.25%和25.00%; 木洞种群最大年龄为5龄, 以1龄、2龄和3龄为优势年龄组, 分别占22.05%、41.73%和29.13%, 年龄结构在江段间存在显著差异(χ2=29.65,P<0.01)。
2.2 体长-体重关系与生长、死亡参数
光泽黄颡鱼的体长-体重关系为:
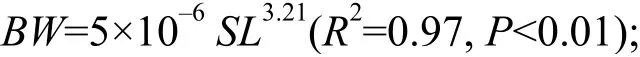
光泽黄颡鱼的体长-体重关系的b值与3差异显著(t-test,P<0.01), 属正异速生长, 且木洞的b值(3.25)显著高于秭归(3.18)和万州的值(3.19; Kruskal-Wallis test,H=8.60,P<0.01)。光泽黄颡鱼体长、体重的VBGF生长方程如下:
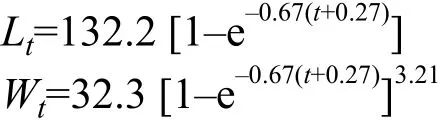
体长生长速度和生长加速度如下:
体长生长速度: dlt/dt=88.6e-0.67(t+0.27)
体长生长加速度: d2lt/dt2= -59.3e-0.67(t+0.27)
体重生长速度和生长加速度如下:
体重生长速度: dWt/dt=69.5 e-0.67(t+0.27)[1-e-0.67(t+0.27)]2.21
体重生长加速度: d2Wt/dt=46.6e-0.67(t+0.27)[1-e-0.67(t+0.27)]1.21(3.21 e-0.67(t+0.27)-1)
如图 4所示, 根据以上计算得到光泽黄颡鱼的体重生长拐点年龄为1.5龄, 拐点体重为8.8 g, 总死亡系数(Z)为1.39/a, 自然死亡系数(M)为0.78/a, 捕捞死亡系数(F)为0.61, 据此得到种群开发率(E)为0.44。
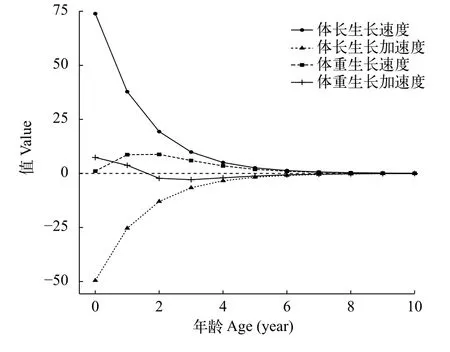
图 4 三峡水库光泽黄颡鱼生长速度、生长加速度与年龄关系Fig. 4 Relationships between growth speed, growth acceleration and ages of P. nitidus in the Three Gorges Reservoir
2.3 耳石轮径与体长关系
光泽黄颡鱼体长(SL)与耳石轮径(OR)关系为:
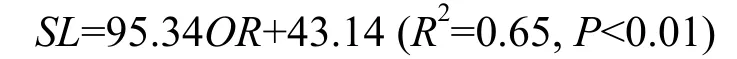
不同江段种群的体长与耳石轮径关系分别为:
秭归:SL=98.86OR+40.21 (R2=0.68,P<0.01);
万州:SL=85.78OR+46.56 (R2=0.63,P<0.01);
木洞:SL=98.95OR+43.41 (R2=0.67,P<0.01)。
光泽黄颡鱼的体长与耳石轮径关系的斜率在江段间无显著差异(ANCOVA,F=1.66,P=0.19; 图 5)。按照获得的所有个体的体长与耳石轮径关系进行体长退算(表 1), 实测的平均体长与退算体长间无显著差异(t-test,t=-0.71,P=0.51) 。
2.4 肥满度、肝体系数和脂体系数
光泽黄颡鱼的肥满度、肝体系数和脂体系数在库区的平均值分别为1.063±0.027、0.026±0.001和0.007±0.001, 均呈现显著的空间差异(P<0.01), 且具有相似的变化趋势, 除肝体系数在木洞和万州的值无显著差异外, 其余均是在木洞的值显著高于在秭归和万州的值(表 2)。光泽黄颡鱼的肥满度与肝体系数、脂体系数均呈显著的线性关系(图 6; 肝体系数:HSI=0.028Kr-0.00332,R2=0.16,P<0.01; 脂体系数:LSI=0.0325Kr-0.0277,R2=0.22,P<0.01)。
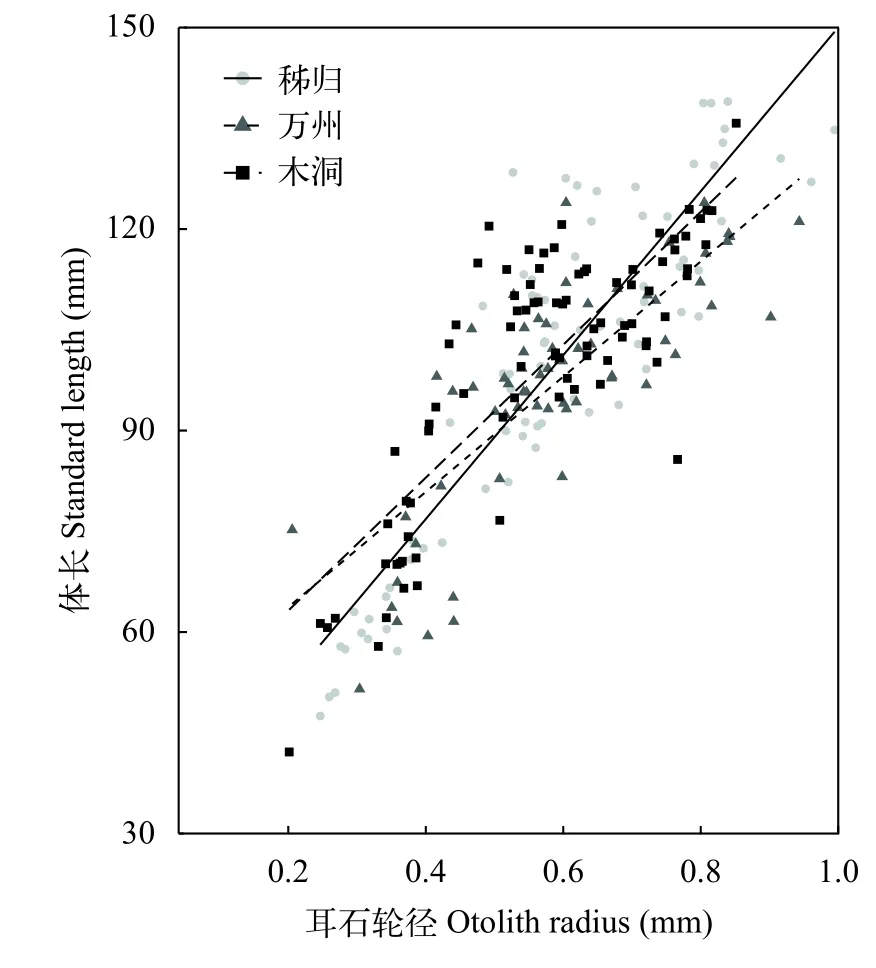
图 5 三峡水库光泽黄颡鱼体长与耳石轮径的关系Fig. 5 Relationship between standard length and otolith radius of P. nitidus in the Three Gorges Reservoir
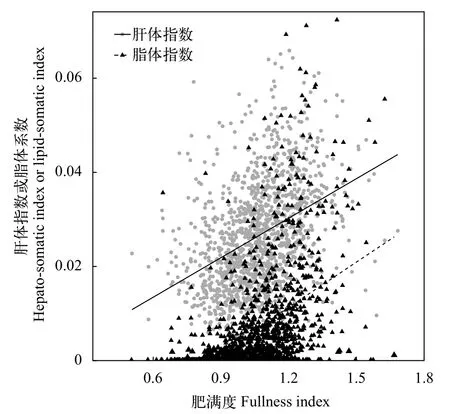
图 6 三峡水库光泽黄颡鱼肥满度与肝体系数、脂体系数间的关系Fig. 6 Relationship of the fullness index with the hepato-somatic index and lipid-somatic index of P. nitidus in Three Gorges Reservoir
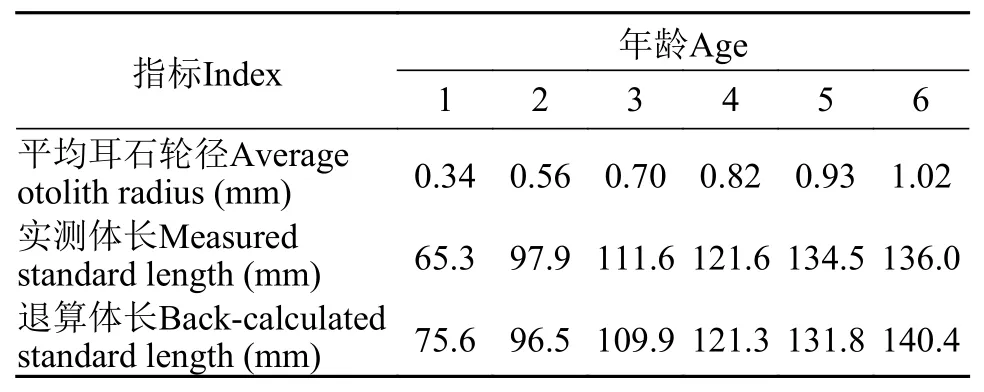
表 1 三峡水库光泽黄颡鱼的体长退算Tab. 1 Back calculation of the standard length of P. nitidus in the Three Gorges Reservoir
3 讨论
3.1 个体大小与年龄结构
耳石是用以鉴定鱼类年龄的重要且精确度较高的材料之一, 已在许多黄颡鱼属鱼类年龄与生长研究中用作鉴定材料[31—33]。本研究以微耳石作为年龄鉴定材料, 耳石轮径与体长显著正相关, 表明微耳石可作为其年龄鉴定的有效材料[34]。本研究共鉴定出6个年龄组, 优势年龄组为2龄, 比洞庭湖(3龄)、赣江的光泽黄颡鱼种群的年龄结构(3龄)更丰富, 最大体长(146.7 mm)也超过洞庭湖(约130 mm)和赣江的种群(约125 mm)[34,35], 表明三峡水库光泽黄颡鱼种群具有更完整的种群结构。此外, 由于缺乏三峡水库蓄水前有关光泽黄颡鱼的生物学数据,无法进行生物学特征的直接比较。本研究中光泽黄颡鱼的平均体重较1976年的估算值[14]降低, 这可能与蓄水前后捕捞方式及环境变化有关[36]。总体而言, 光泽黄颡鱼作为一种底层性小型鱼类, 适应环境能力较强, 其种群结构受蓄水的影响较小。

表 2 三峡水库不同江段光泽黄颡鱼肥满度、肝体系数和脂体系数(平均值±标准误)Tab. 2 The fullness index, hepato-somatic index, and lipidsomatic index of P. nitidus in different sections of the Three Gorges Reservoir (mean±SE)
3.2 生长与死亡特征
鱼类种群的生长和死亡等特征是科学评估和管理鱼类资源的关键[2]。体长和体重关系中的 b值可以用来判断鱼类是否处于等速生长。b值具有物种特异性, 能反映鱼类在不同环境的生长情况和饵料状况等[37]。三峡水库光泽黄颡鱼的b值为3.21, 表现为正异速生长, 这与嘉陵江下游、淀山湖的光泽黄颡鱼种群的生长式型不尽相同[12,38], 嘉陵江下游草街电站坝上水域种群表现为正异速生长(b=3.30),坝下水域种群则为等速生长(b=2.90); 淀山湖种群则表现为负异速生长(雌: b=2.53; 雄: b=2.56)。这种差异可能是栖息地环境(如饵料可得性、流态等)和遗传多样性等方面的不同所导致[39]。此外,我们发现三峡水库库尾光泽黄颡鱼种群的b值显著高于库首和库中种群, 这与肥满度的空间变化趋势一致, 这可能主要源于不同江段之间生境及相应的饵料基础的差异。
在渔业资源利用过程中, 为保持鱼类资源的可持续性, 宜选择在鱼类快速生长至缓慢生长的节点,即生长速度最大时(生长拐点)加以利用, 这是指导渔业资源管理的关键参数之一[1]。本研究评估了光泽黄颡鱼的体重生长拐点(1.47龄)和拐点体重(8.79 g)。在渔获物中, 1龄的光泽黄颡鱼个体数占比达到所有个体数21.05%, 对应的平均体重(3.46 g)也低于拐点体重, 表明全面禁渔措施实施前的渔业捕捞会对光泽黄颡鱼的补充群体造成一定的不利影响。
鱼类的开发系数也是渔业管理中的一个重要参数, 鱼类的合理开发强度宜为开发强度低于0.5,低于这个强度属于轻度开发, 高于这个强度则是处于过度开发状态, 会对鱼类种群的可持续产生负面影响[40]。据此, 三峡水库光泽黄颡鱼种群(E=0.44)尚处于轻度开发状态, 这与对淀山湖光泽黄颡鱼的研究结果相似[41]。此外, 对三峡水库蛇鮈(Saurogobio dabryi)等18种小型鱼类的种群特征研究[42]发现,这些小型鱼类种群的平均开发率为0.44, 处于轻度开发状态。这可能由于光泽黄颡鱼等鱼类个体较小, 各类渔具选择性捕捞造成的死亡率较低, 这有利于蓄水后光泽黄颡鱼种群的稳定[16,18]。
3.3 营养状况
肥满度是衡量鱼体能量储备、营养状况和环境条件的重要指标[43]。本研究发现三峡水库光泽黄颡鱼的肥满度呈现明显的空间变化, 库尾优于库中和库首, 这与三峡水库另一种杂食性鱼类—蛇鮈(Saurogobio dabryi)肥满度的空间变化规律一致, 反映了库尾种群营养状况更优[6]。食物组成能对鱼类的肥满度产生明显影响[44,45]。光泽黄颡鱼的营养状况空间变化与建坝后底栖动物生物量在库尾最高的空间规律相一致[20], 表明饵料的可得性可能是影响其肥满度的重要因素。库尾底栖动物生物量高, 其在库尾光泽黄颡鱼的食物中所占比例也明显高于库首和库中[22]。三峡水库中以底栖动物为主要食物的黄颡鱼种群的肥满度优于以虾类为主要食物的种群, 可能是由于摄食底栖动物相对于摄食虾类的净能量收益更高。Paukert和Rogers[46]也发现过类似现象, 科罗拉多河的偏翼亚口鱼(Catostomus latipinnis)的肥满度与底栖动物丰度正相关。然而,Perera等[33]却发现库首的黄颡鱼种群肥满度高于库尾种群, 环境的变化可能并不是这种差异的唯一原因, 与不同鱼类之间摄食习性等方面的差异可能也有关[43]。鉴于此, 本研究采用了多种指数来表征鱼类营养状况。
肝脏是鱼类的重要能量储存器官, 脂肪是重要的能量储存形式[47,48]。有研究认为肥满度表征的是存储于肌肉中的能量, 能代表整个鱼体的能量储备, 而肝体系数则更多地表征以脂类形式储存于肝脏中的能量[49]。在本研究中, 光泽黄颡鱼的肥满度与肝体系数、脂体系数均呈显著的正相关, 且肝体系数与脂体系数的空间变化趋势也与肥满度一致,表明肝体系数和脂体系数亦能反映出鱼类的能量储备和营养状况, 三峡水库蓄水后底栖动物资源的空间差异可能也是导致肝体指数与脂体系数空间差异的重要因素。
猜你喜欢
中国水利(2020年23期)2020-12-24
中华养生保健(2020年7期)2020-11-16
中国水利(2020年14期)2020-08-02
科学导报(2020年27期)2020-05-13
老友(2018年7期)2018-08-08
科学生活(2017年9期)2017-10-16
浙江海洋大学学报(自然科学版)(2012年6期)2012-10-21
