
三峡库区短颌鲚年龄和生长特性的研究
2022-01-20 08:09杨丽亚吕红健白云鹏苏胜齐姚维志
水生生物学报 2022年1期
杨丽亚 吕红健 付 梅 白云鹏 苏胜齐 姚维志
(西南大学水产学院, 西南大学渔业资源环境研究中心, 重庆 400716)
三峡库区是指受长江三峡工程淹没的地区, 位于长江上游, 由湖北省和重庆市所辖区县构成, 其中重庆市所辖区县占大部分, 主要包括巫山、巫溪、云阳、奉节、万州等区(县)。综合历史调查数据显示, 三峡大坝截流之前, 库区所在长江流域的渔业资源量就已呈现波动或缓慢下降趋势[1]。在三峡大坝截流之后, 三峡库区水位涨落节律与自然枯洪规律的逆反, 以及库区水位的上升, 造成了三峡库区鱼类生境的破碎[2,3]。加之外来水生生物入侵、人为污染和电鱼滥捕等因素, 使得目前三峡库区渔业资源面临着严重威胁[4,5]。相关研究表明三峡库区蓄水以来, 随着库区水体加深和水流减缓,形成了一个生态位严重空缺的类湖泊生态系统, 其营养物质远没有得到充分利用, 这为外来鱼类的进入和种群暴发提供了有利的环境条件[6,7]。2012年,巴家文和陈大庆[5]对三峡库区外来入侵鱼类的一项调查研究显示, 库区及长江上游外来入侵鱼类呈增长态势, 目前已发现外来入侵鱼类23种, 其中有9种是从我国其他水系侵入三峡库区, 剩下14种分别来自美洲、欧洲和非洲, 且部分种类处于种群数量的暴发阶段。
短颌鲚(Coilia nasus), 隶属鲱形目(Clupeiformes)、鳀科(Engraulidae)、鲚属(Coilia), 《中国动物志》将短颌鲚(曾被命名为Coilia brachygnathus)和湖鲚(曾被命名为Coilia nasus taihuensis)归为刀鲚(又称长颌鲚), 并认为它们均属淡水定居型刀鲚[8]。长期以来, 关于短颌鲚的分类地位一直存在争议。1908年Kreyenberg和Pappenheim[9]根据洞庭湖相关标本确立短颌鲚为一个物种, 至20世纪末期, 袁传宓等[10]、成庆泰和郑葆珊[11]、东海水产研究所等[12]的研究仍将短颌鲚认定为区别于刀鲚的一个物种, 他们认为短颌鲚与刀鲚的显著差别在于上颌骨长短, 短颌鲚上颌骨短, 向后不超过鳃盖骨, 而刀鲚上颌骨长, 向后伸达胸鳍基部,这一观点当时被国内大多数学者所认可。20世纪初短颌鲚分类出现新的观点, 学者们对刀鲚、短颌鲚及湖鲚的mtDNA Cytb序列变异、D-loop序列变异及线粒体COⅠ基因条形码序列变异进行研究后发现刀鲚、短颌鲚和湖鲚三者间的遗传距离均未超过0.02[13—15], 证实了短颌鲚及湖鲚均未到达种水平, 从而得出结论短颌鲚与湖鲚均为刀鲚的淡水生态型。
据历史文献资料记载, 短颌鲚主要分布在长江及淮河流域的中、下游, 及与长江流域相通的淡水湖泊, 例如鄱阳湖和洞庭湖[16,17], 截至2018年底未见短颌鲚在三峡库区分布的研究或调查报道。查阅长江上游渔业资源调查资料、《四川鱼类志》,均显示2015年前, 短颌鲚在长江上游无迹可寻。在三峡库区蓄水后, 库区渔民在捕捞的渔获物中不断发现疑似刀鱼的新种类, 2015年该种类数量在库区明显增长, 2017—2018年该种类单船日捕捞量可达近100 kg(西南大学渔业资源环境研究中心未发表数据)。2019年Yang等[18]通过线粒体DNA全序列检测方法, 首次确认并报道了三峡库区中疑似刀鱼的新种类为短颌鲚, 据西南大学渔业资源环境研究中心2018—2020年调查显示, 目前短颌鲚在三峡库区的扩散范围至少下游已至重庆巫山段, 上游已达重庆长寿段。历史研究表明, 短颌鲚具有适应力强、性成熟早和繁殖率高的生物学特性, 其以江河中浮游动物、小鱼虾和鱼苗为食, 又可作为大型肉食性鱼类的饵料[19,20], 在分布水域的食物链中占有重要地位。就目前三峡库区短颌鲚的研究情况来看, 还停留于报道该物种出现于库区阶段, 短颌鲚出现于三峡库区对水域鱼类群落结构及生态环境已产生或将产生怎样的冲击, 目前相关研究仍为空白, 开展这些研究需建立在掌握三峡库区短颌鲚生物学特性的基础之上。因此, 本研究以三峡库区的短颌鲚为研究对象, 分析目前三峡库区现有短颌鲚种群的年龄和生长特性, 以期为全面了解三峡库区现有短颌鲚种群的结构和动态特征提供基础数据, 为探索短颌鲚对其他关联水生生物的影响提供理论依据, 并为我国三峡库区鱼类种群动态监测、生物多样性保护和资源种群的科学管理提供理论和技术支撑。
1 材料与方法
1.1 实验材料
2019年7—12月分别在云阳(澎溪河)、奉节(草堂河、梅溪河)和巫山(大宁河)使用三层刺网和地笼采集短颌鲚样本, 共采集459尾, 将所采集样本带回实验室统一分析, 样本采集信息见表 1。各采样点地理位置如图 1所示。此外, 本研究选用短颌鲚中轴部侧线下方第3至第4行鳞片作为低龄短颌鲚样本(年龄≤3)的年龄鉴定材料, 选用上述鳞片及第5至第10号脊椎骨同时作为高龄短颌鲚样本(年龄>3)的年龄鉴定材料。

表 1 三峡库区短颌鲚的采集信息Tab. 1 Basic data of Coilia nasus from different sampling sites in the Three Gorges Reservoir Region

图 1 三峡库区短颌鲚采样点示意图Fig. 1 Sampling sites of Coilia nasus in the Three Gorges Reservoir Region
1.2 样品处理
使用精度为0.10 cm的量鱼板和精度为0.10 g的电子天平(YP6001N, 舜宇, 中国上海)测量短颌鲚样本的全长、体长、体重、胃肠重及性腺重等常规生物学指标, 鉴定样本性别并记录性腺发育分期。鳞片和脊椎骨的处理方法分别参照罗红波[21]和谢从新[22]。
1.3 数据处理与分析
在本研究中, 数据的统计分析采用Microsoft Excel(2016)、SPSS(IBM SPSS Statistics 20)、单因素方差分析法(ANOVA)、协方差分析法(ANCOVA)和t检验法, 数据采用平均值±标准误(即mean±SE)表示。图表绘制采用Microsoft Excel(2016)、Origin(2018)。
体长和体重的关系用幂函数W=aLb来描述; 肥满度K(%)的计算公式为K=(W/L3)×100%; 体长相对生长率(RL)计算公式为RL=(L2-L1)/L1(t2-t1), 体重相对生长率(RW)计算公式为RW=(W2-W1)/W1(t2-t1); 生长指标(C1t)计算公式为C1t=(lnL2-lnL1)L1/(t2-t1)。
用非线性回归拟合Von Bertalanffy生长方程(VBGF)、Gompertz生长方程(GGF)和Logistic生长方程(LGF), 3种方程表达式见表 2。以上3种生长方程的参数值估算采用试值法[23]。VBGF体长、体重的生长速度(一次微分)和加速度(二次微分)方程分别为和

表 2 拟合三峡库区短颌鲚生长方程的数学模型Tab. 2 Mathematical models describing the growth of Coilia nasus in the Three Gorges Reservoir Region
上述公式中W为样本体重(g),L为样本体长(cm); 体长和体重关系式中a为常数,b为指数;t1和t2分别代表单位时间的起点和止点;L2、L1和W2、W1分别为终止(t2)和起始(t1)时的体长和体重;L∞、W∞、k、t0、a为生长方程的参数。
2 结果
2.1 鳞片和脊椎骨的形态与年轮特征
三峡库区短颌鲚鳞片属圆鳞, 具一条辐射沟,鳞片的年轮标志为疏密型, 窄带(即窄而密的生长带)与宽带(即宽而疏的生长带)的交界处即为年轮(图 2A)。在使用鳞片进行年轮鉴别时存在副轮和幼轮的干扰, 其中副轮不清晰, 未形成完整的轮圈,幼轮与第一年轮较难区分。此外, 副轮及幼轮多出现于3龄以上短颌鲚样本的鳞片中, 1至3龄样本鳞片中很少存在。本研究中由于低龄短颌鲚(年龄≤3)的脊椎骨细小, 锥体凹面残留的脊索不易清理干净, 干扰年轮观察, 因此不宜用于年龄鉴定。高龄短颌鲚(年龄> 3)脊椎骨经处理后可见清晰的年轮(图 2B)。
2.2 体长、体重组成及性比
本研究459尾三峡库区短颌鲚样本中, 渔获样本的最小体长为9.80 cm, 最大体长为31.80 cm, 平均体长为(16.57±0.21) cm; 最小体重为3.30 g, 最大体重为118.40 g, 平均体重为(19.14±0.78) g。此外,样本体长多数集中在10.00—24.00 cm, 且样本数占比95.80%(图 3); 体重多数集中在3.00—48.00 g, 且样本数占比95.60%(图 4)。

图 2 三峡库区短颌鲚鳞片(A)和脊椎骨(B)两种年龄鉴定材料年轮特征示意图Fig. 2 Annuli characteristics of scale (A) and vertebrae (B) of Coilia nasus from the Three Gorges Reservoir Region A鳞片: a. 辐射沟; b. 第1年轮; c. 第2年轮; d. 鳞焦; B. 脊椎骨: 1—5. 年轮A scale: a. scale groove; b. the 1st annual ring; c. the 2nd annual ring; d. scale focus; B bertebrae: 1 to 5. annual ring
性腺分析结果表明, 三峡库区短颌鲚样本1龄以上即可达性成熟, 通过观察性腺可辨别样本性别,目前三峡库区短颌鲚群体总体性比为2.16∶1(♀∶♂)。本研究459尾渔获样本中仍有4尾1龄样本未达性成熟。在性成熟样本中, 雄性样本有144尾, 其体长为9.80—29.80 cm, 平均体长为(16.44±0.44) cm, 体重为3.20—94.80 g, 平均体重为(19.15±1.63) g; 雌性样本有311尾, 其体长为10.10—31.80 cm, 平均体长为(16.71±0.26) cm, 体重为3.40—118.40 g, 平均体重为(19.33±0.92) g。雄性生物学最小型对应体长为9.80 cm, 体重为3.30 g, 年龄为1龄; 雌性生物学最小型对应体长为10.10 cm, 体重为3.40 g, 年龄为1龄。
2.3 年龄结构
年龄鉴定结果表明, 目前三峡库区短颌鲚群体年龄组成为1—6龄, 且1—3龄样本数占比90.85%。群体的平均年龄偏低, 年龄结构较简单, 为增长型种群, 在图像上呈金字塔形(图 5)。比较4条采样河流短颌鲚群体年龄组成发现, 仅大宁河和草堂河存在5龄、6龄样本, 草堂河5龄、6龄样本各1尾, 梅溪河样本最大年龄为4龄, 澎溪河样本最大年龄为3龄(表 3)。
2.4 肥满度
在本研究中, 三峡库区短颌鲚样本的平均肥满度为(0.34±1.46)‰。ANOVA分析显示, 只有1龄组[(0.35±2.17)‰,n=199]与2龄组 [(0.33±2.92)‰,n=118]肥满度存在显著性差异(P<0.05), 且1龄个体肥满度显著高于2龄个体。除此之外, 其余各年龄组之间肥满度不存在显著性差异(P>0.05; 图 6)。

图 3 三峡库区短颌鲚的体长组成Fig. 3 Body length distribution of Coilia nasus samples from the Three Gorges Reservoir Region

图 4 三峡库区短颌鲚的体重组成Fig. 4 Body weight distribution of Coilia nasus samples from the Three Gorges Reservoir Region

图 5 三峡库区短颌鲚群体的年龄组成Fig. 5 Age composition of Coilia nasus population in the Three Gorges Reservoir Region (n=459)
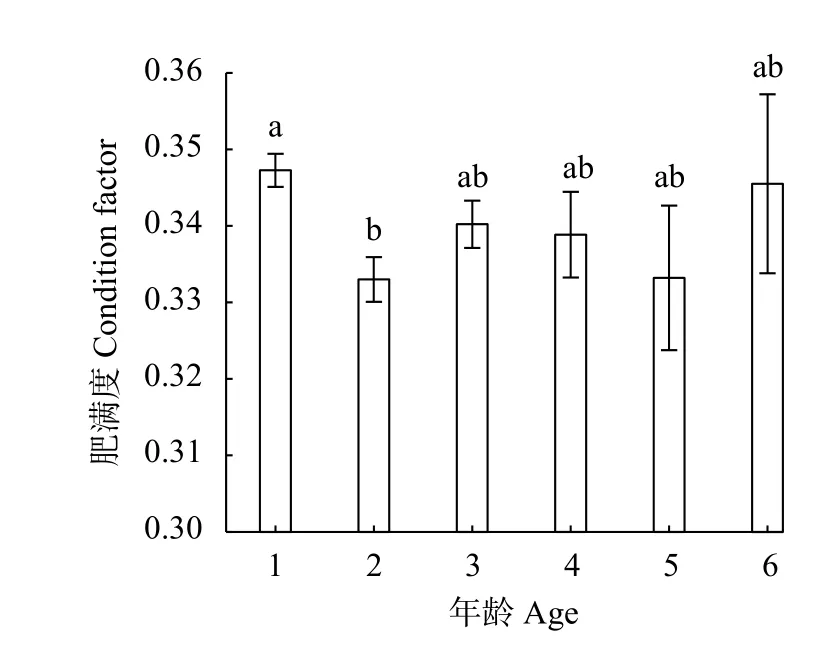
图 6 三峡库区不同年龄组短颌鲚肥满度Fig. 6 Condition factor of Coilia nasus from different age groups in the Three Gorges Reservoir Region
2.5 相对生长率和生长指标
本研究结果表明, 三峡库区短颌鲚3龄之前, 体长和体重的相对生长率均较大, 生长指标高, 生长较快; 3—4龄期间, 体长和体重的相对生长率减小,生长指标较低, 生长减缓; 4龄以后, 生长速度总体存在缓慢的上升趋势(表 4)。
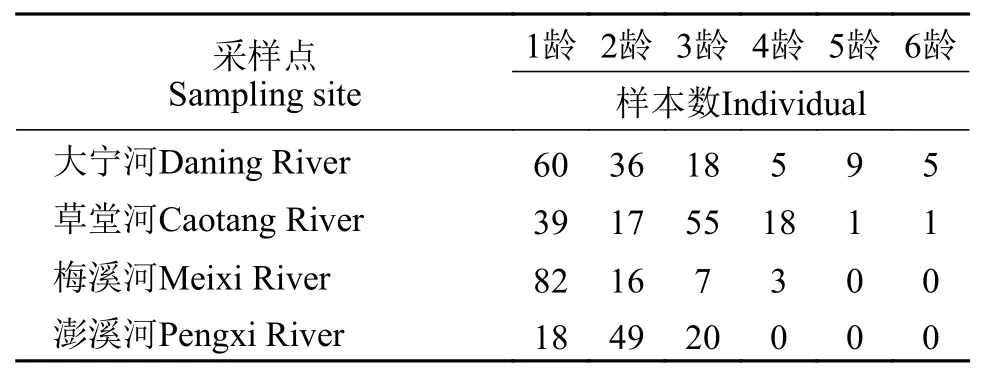
表 3 三峡库区不同采样点短颌鲚样本的年龄组成Tab. 3 Age composition of Coilia nasus from different sampling sites in the Three Gorges Reservoir Region

表 4 三峡库区短颌鲚的相对生长率和生长指标Tab. 4 Relative growth rate and growth index of Coilia nasus population in the Three Gorges Reservoir Region
2.6 体重-体长关系及生长方程
三峡库区现有短颌鲚群体的体重-体长关系式为:群体总体:W=0.0038L2.957(n=459,R²=0.99)

协方差分析(ANCOVA)显示三峡库区雌性、雄性短颌鲚的体重-体长关系无显著性差异(n=455,F=0.303,P>0.05), 体重-体长生长方程可合并, 群体总体:W=0.0038L2.957(n=459,R²=0.99)。由上式可知目前三峡库区短颌鲚群体体重-体长关系指数b=2.957, 经t检验, 指数b与3存在显著性差异(t=2.087,P<0.05), 表明其生长为异速生长类型(图 7)。
如表 5所示, 3种方程的拟合程度均较高(R2≥0.94), 但GGF和LGF拟合得到的L∞、W∞低于部分实测值最大值。同时, 考虑到VBGF被广泛应用于描述鱼类的生长规律, 本研究最终采用VBGF来描述三峡库区短颌鲚的生长。
统计得出雌性、雄性理论体长(Von Bertalanffy生长方程计算得出)与实测平均体长(表 6), 将二者进行配对t检验, 结果显示实测体长与理论体长无显著性差异(♀:t=-0.600,P>0.05; ♂:t=0.148,P>0.05), 说明理论体长可信, 即上述Von Bertalanffy生长方程能够较好地拟合三峡库区短颌鲚的生长规律。
对上述Von Bertalanffy生长方程分别求一阶和二阶微分, 得到三峡库区短颌鲚体长和体重的生长速度方程和生长加速度方程为:

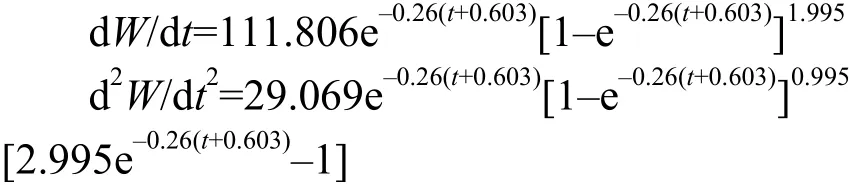
根据三峡库区短颌鲚体长和体重的Von Bertalanffy生长方程、生长速度方程、生长加速度方程, 分别做出雌、雄群体体长和体重的生长曲线(图 8)、生长速度曲线(图 9)、生长加速度曲线(图 10)。研究结果显示, 随着年龄的增长, 雌性和雄性体长生长速度均逐渐下降, 但速度一直为正值,体长生长加速度曲线逐渐上升但均为负值(图 9)。由此表明随着年龄的增长, 雌性和雄性体长生长速度下降的幅度逐渐减小。然而, 与体长生长不同,雌性、雄性体重生长速度和加速度曲线均具有明显的拐点, 在体重生长拐点处d2W/dt2=0, 即tr=ln3/k+t0, 将雌性、雄性的体重生长方程中的k、t0值分别带入tr=ln3/k+t0中, 得到雌性、雄性体重生长的年龄拐点分别为4.62龄、3.62龄(图 10)。在拐点年龄以前体重生长速度逐渐上升, 体重生长加速度逐渐下降, 二者均为正值; 在拐点以后体重生长速度逐渐下降但为正值说明体重增长减缓, 体重生长加速度先下降后缓慢上升, 且值为负值。

图 8 三峡库区短颌鲚体长、体重生长曲线Fig. 8 The growth curve of body length and body weight of Coilia nasus in Three Gorges Reservoir Region
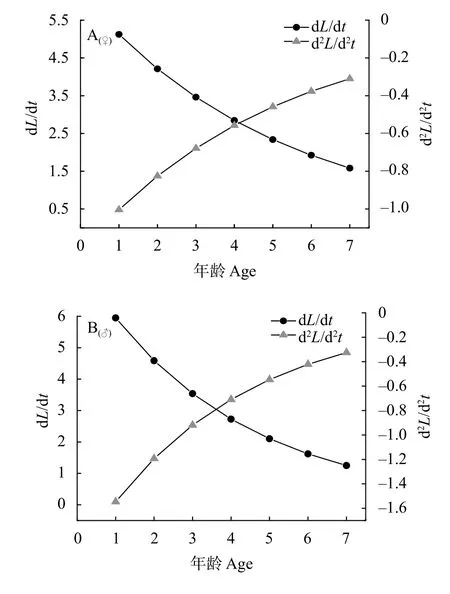
图 9 三峡库区雌性、雄性短颌鲚体长生长速度和加速度曲线Fig. 9 Growth in body length as dL/dt and d2L/dt2 curves of female and male Coilia nasus in the Three Gorges Reservoir Region

图 10 三峡库区雌性、雄性短颌鲚体重生长速度和加速度曲线Fig. 10 Growth in body weight as dW/dt and d2W/dt2 curves of female and male Coilia nasus in the Three Gorges Reservoir Region

表 5 三峡库区短颌鲚生长方程Tab. 5 The growth functions of Coilia nasus in the Three Gorges Reservoir Region

表 6 三峡库区雌、雄性短颌鲚理论体长与实测体长Tab. 6 Theoretical and measured body length of Coilia nasus in the Three Gorges Reservoir Region
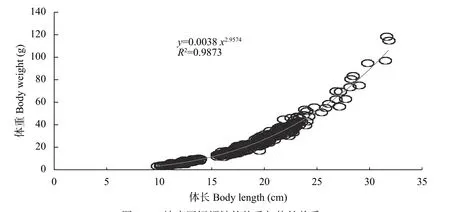
图 7 三峡库区短颌鲚的体重与体长关系Fig. 7 The relationship between body length and body weight of Coilia nasus in the Three Gorges Reservoir Region
3 讨论
3.1 三峡库区短颌鲚体型大小及生长规律
研究中三峡库区短颌鲚最大样本的体长为31.80 cm, 体重为118.40 g, 为雌性样本, 对应年龄为6龄。最大体长、体重结果与长江天鹅洲故道(湖北石首江段)[21]的报道相近, 但后者对应年龄却为4龄。吴斌等[24]报道显示鄱阳湖通江水道屏峰段(江西湖口江段)短颌鲚的最大体长、体重分别为31.00 cm、125.50 g, 但对应年龄不详。三峡库区短颌鲚平均体长为(16.57±0.21) cm, 平均体重为(19.14±0.78) g, 平均体长、体重不及鄱阳湖(江西湖口江段)短颌鲚群体(平均体长19.26 cm, 平均体重25.38 g)[25]。此外, 三峡库区短颌鲚体长多数集中在10.00—24.00 cm, 体重多数集中在3.00—48.00 g,占比达95.00%, 小型个体居多。上述长江中下游短颌鲚群体的体长与体重研究结果显示长江流域自上游至下游, 同等规格的短颌鲚个体对应的年龄逐渐变小[21,26,49], 即三峡库区短颌鲚群体与下游群体相比体型偏小。分析其中原因我们总结为以下几点: 首先, 查阅历史资料不难发现, 下游河段一般较上游河段年平均水温更高、营养更丰富和栖息地稳定性更高[27—29], 生境上的这些差异可能是引起短颌鲚不同地理群体生长状况不一致的原因。其次,三峡大坝的修建, 使原有河流型生境转变为河流型、过渡型和湖泊型三种不同的生境类型, 造成库区水域部分鱼类的适宜栖息地面积减少和破碎化,并对上下游鱼类的基因交流造成一定的影响, 进而导致其遗传多样性和生长性能的降低[30—35]。换言之, 三峡水库蓄水所引起的生境转变与破碎, 所引起的库区群体的遗传多样性和生长性能的降低, 可能是致使库区短颌鲚个体小型化的又一个原因。最后, 长江实行全面禁渔之前, 由于人类频繁的水利活动、环境污染和过度捕捞, 导致长江流域内鱼类资源受到严重影响, 小型化趋势明显[36]。综上所述, 引起三峡库区短颌鲚群体较中、下游群体小型化的原因, 可能与上述地理位置不同所造成的水文条件差异, 栖息地破碎, 以及多类型的人为因素有关, 上述推测也有待进一步的研究验证。
肥满度是反映鱼体在不同时期或不同水域营养与摄食情况的一个重要指标, 且随着鱼体体长和重量的关系变化[37]。三峡库区现有短颌鲚群体的平均肥满度为(0.34±1.46)‰, 相较于长江天鹅洲故道(湖北石首江段)与鄱阳湖(江西湖口江段)短颌鲚群体更大[21,26]。上述差异可能是由于短颌鲚进入三峡库区时间较短, 种群数量还远未达到环境容纳量, 即种群容量剩余量大, 饵料生物相对充足引起的。关于肥满度本研究还发现, 三峡库区短颌鲚仅1龄组与2龄组肥满度之间存在显著性差异(P<0.05),且2龄组个体肥满度更低。从体长相对生长率和生长指标研究结果可以看到2龄组的两个指标均为最高, 据此推断三峡库区短颌鲚在1—2龄生长期间,用于身体体长生长的能量有所增加, 导致体长与体重生长速度不同步, 从而进一步引起不同年龄组短颌鲚肥满度的差异。
体重-体长关系式中的b值反映鱼类的生长类型,b接近于3为等速生长, 反之, 则为异速生长;a值为生长的条件因子, 饵料基础和水文条件较好时条件因子较大[23,38]。同时,a值与肥满度之间存在正比关系, 在一定条件下肥满度也可用a值表示。三峡库区现有短颌鲚群体体重-体长关系式中指数b=2.957, 该值接近于3, 与长湖(湖北荆州江段)、长江天鹅洲故道(湖北石首江段)和鄱阳湖(江西湖口江段)短颌鲚群体相近, 但生长类型不一致[19,21,26],这可能与水域环境条件差异、采样季节及生长类型确定方法不同有关[22,39]。本研究中条件因子a=0.0038, 这与鄱阳湖(江西湖口江段)短颌鲚生长的条件因子相近[40], 而较长湖短颌鲚生长的条件因子更大[19]。鄱阳湖为中国最大的淡水湖泊(平均水温为20.30℃, pH为7.50, DO为7.20 mg/L, 且水质状况健康)[41,42], 其水文条件适宜短颌鲚的生存[20]。此外, 短颌鲚在鄱阳湖渔获物中所占比例也侧面反映了短颌鲚对鄱阳湖生境的高度适应性(以鄱阳湖湖口水域为例, 平均单网短颌鲚的重量占渔获物总重量的(12.77±6.89)%; 平均单网短颌鲚的个体数占渔获物总数量的(19.46±20.78)%; 9、10月份, 单网短颌鲚的个体数占渔获物总数量的50%左右)[25]。短颌鲚在三峡库区和鄱阳湖两水域生长的条件因子相近, 可以推断出对于短颌鲚而言三峡库区目前饵料基础和水文条件也较好。
三峡库区短颌鲚生长系数k=0.217, 这与长湖(湖北荆州江段)、长江天鹅洲故道(湖北石首江段)和鄱阳湖(江西湖口江段)短颌鲚的生长系数研究结果相近[19,21,24]。Branstetter[43]认为k值在0.20—0.50属快速生长鱼类, 据此推断三峡库区短颌鲚为快速生长鱼类。本研究由Von Bertalanffy生长方程拟合三峡库区短颌鲚生长规律求得的雌、雄个体渐近体长分别为L∞=38.567 cm(♀)和L∞=34.711 cm(♂), 渐近体重分别为W∞=183.029 g(♀)和W∞=143.599 g(♂)。上述渐近体长L∞和渐近体重W∞数据远大于实测最大体长和最大体重, 且大于长湖(湖北荆州江段)和鄱阳湖(江西湖口江段)短颌鲚的渐近体长和体重研究结果[19,24], 表明三峡库区短颌鲚目前还具备较大的生长潜力。此外, 关于生长加速度本研究结果显示三峡库区雌、雄短颌鲚的体重生长拐点年龄分别为4.62龄(♀)和3.62龄(♂), 远大于性成熟年龄, 属于衰老拐点, 三峡库区短颌鲚在达生长拐点之前即可进行多次繁殖, 促使种群世代更新快, 这将有利于进一步保证种群的增长和延续[44]。
3.2 三峡库区短颌鲚种群的演替趋势及扩散规律
性比是鱼类种群结构的重要指标, 其可通过影响种群繁殖率进而造成种群数量变动[45], 三峡库区现有短颌鲚种群的总体性比为2.16∶1(♀∶♂)。历史研究发现, 长湖(湖北荆州江段)短颌鲚群体性比为1.28∶1(♀∶♂)[19]、长江天鹅洲故道(湖北石首江段)短颌鲚群体性比为1.19∶1(♀∶♂)[21]、鄱阳湖和赣江(江西湖口江段)短颌鲚群体性比为1∶1.7(♀∶♂)[40],三峡库区短颌鲚种群的性比结果均高于以上3个群体。一般而言, 在一个鱼类繁殖群体中, 雄性个体数量增加引起的性比变化, 会使群体繁殖潜力呈现降低的趋势, 且使雌性占比增加是种群维持和增加个体数量的重要繁殖策略[44,46]。因此, 根据本研究结果可以推测三峡库区现有短颌鲚群体的繁殖潜力较长江中下游群体高, 种群具备较高持续增长能力。
一般来说, 环境对鱼类产生的影响以及鱼类自身对于环境的适应可以在其种群年龄结构中有所体现[44]。以本研究结果为例, 三峡库区现有短颌鲚群体年龄组成为1—6龄, 且还发现部分雌性怀卵样本, 上述结果表明短颌鲚在三峡库区已经形成了可持续种群, 且其对库区的生存环境已产生了适应。此外, 本研究结果还发现目前三峡库区短颌鲚种群年龄组成偏低, 年龄结构较为简单, 且年龄组成在图像上呈金字塔形, 种群为增长型。通常一个未开发的自然种群, 若资源量迅速增大就有大量的补充个体, 渔获物的年龄组成则偏低, 在适宜环境下该种群数量会激增, 最终会逐渐形成年龄结构复杂的稳定种群[37]。因此, 综合年龄结构研究结果可以推测未来三峡库区短颌鲚种群数量仍有继续增加的趋势。关于年龄结构, 本研究还发现4个采样点仅大宁河和草堂河存在5龄、6龄样本, 梅溪河样本最大年龄为4龄, 澎溪河样本最大年龄为3龄。群体年龄组成在空间分布上的这一差异, 也从侧面反映了短颌鲚在三峡库区的扩散规律, 即短颌鲚在三峡库区扩散的起点可能为长江巫山段及其下游, 且种群扩散的方向为沿长江自东向西, 向上游扩散。这一推测也得到了杨帆[47]研究的印证, 其基于线粒体DLoop序列初步比较了三峡库区与洞庭湖(湖南岳阳江段)短颌鲚群体的遗传多样性, 发现2个群体遗传距离高度接近, 并据此推测三峡库区短颌鲚群体可能来源于长江下游。
Liu等[48]研究指出未来气候变化将增加中国内陆淡水鱼类(包括外来鱼类)的栖息地适应性, 从而增大外来鱼类入侵风险。随着鱼类耳石微化学技术近年来在鲚属鱼类研究中的广泛应用, 现已证实单纯利用上颌骨的长短并不能作为有效判别长江刀鲚资源群体中过河口性溯河洄游个体和淡水定居个体的标准[49,50]。换言之, 单从形态学上判别的“短颌鲚”有可能属于过河口性溯河洄游群体, 而“长颌鲚”也有可能属于淡水定居型群体。上述研究结果也从渔业资源的个体和种群生态学角度, 反映了目前短颌鲚在长江流域具有较强的生境适应力。此外, 不断变化的社会经济活动, 也可能影响或增加外来鱼类的入侵途径[51]。近年来, 中国西部的人类活动有了明显的增长, 这可能会进一步加剧短颌鲚在长江上游的扩散。因此, 可以推测未来随着气候变化, 人类的社会活动增加, 以及种群数量的继续增长, 短颌鲚在三峡库区和长江上游的分布范围将持续扩大。但是短颌鲚种群在三峡库区, 及长江上游水域的扩散速度和范围还有待今后进一步研究验证。
3.3 三峡库区短颌鲚资源变动可能引起的问题
截至目前, 关于短颌鲚的研究表明其能在淡水中完成生长、发育和繁殖等整个生活史, 食性为肉食性, 其以江河中浮游动物、小鱼虾和鱼苗为食,又可作为大型肉食性鱼类的饵料, 且具有的适应力强、性成熟早和繁殖率高等生物学特性[9,20,21], 因此, 短颌鲚在其分布水域的生态系统食物网中占有着重要地位, 且具有能够迅速定居并建立稳定种群的能力。就本研究结果来看, 三峡库区短颌鲚种群目前呈增长型, 三峡库区短颌鲚资源量的进一步扩增可能将对本土其他水生生物形成潜在威胁, 比如侵占三峡库区部分鱼类的生态位(特别是营养生态位), 破坏三峡库区水域生物多样性, 即通过上行效应和下行效应影响本土水域其他水生生物资源量,并最终威胁到三峡库区水域生态系统的安全与稳定。综上所述, 在本研究的基础上, 未来有必要开展短颌鲚对三峡库区水域生态系统的影响及其风险评价的相关研究, 以期为我国淡水鱼类种群动态监测、生物多样性保护和资源种群的科学管理提供技术支撑。
猜你喜欢
今日农业(2022年15期)2022-09-20
天天爱科学(2022年9期)2022-09-15
昆明医科大学学报(2022年1期)2022-02-28
湖南电力(2021年1期)2021-04-13
红土地(2018年7期)2018-09-26
猪业科学(2015年12期)2015-10-31
中国海洋大学学报(自然科学版)(2014年9期)2014-02-28
中国工程咨询(2012年9期)2012-02-14
中学生物学(2008年6期)2008-08-29
雕塑(1995年4期)1995-07-12
