模式动物(果蝇)体重高低杂种优势组合的转录组差异表达分析
2022-01-20 03:51:06孙新宇鲍海港吴常信
中国畜牧杂志 2022年1期
孙新宇,王 珏,凌 遥,鲍海港,吴常信
(中国农业大学动物科学技术学院,北京 100193)
杂种优势是指杂交后代在生长、繁殖、抗病力、抗逆性等性状上优于双亲均值的现象,受遗传与表观遗传、表型与环境因素的综合影响。杂种优势在动物遗传育种与繁殖领域均有广泛研究和应用,如探究杂种优势对猪精子的影响、结合性能测定和转录组学技术预测家畜杂交改良效果等。利用杂种优势是提高动物产量与质量的重要手段之一,至今仍广泛使用。目前杂种优势的遗传机理尚不明确,主要有两种假说,分别是显性假说和超显性假说,但均有局限,不能统一而详尽地解释所有现象。随着一些重要模式生物(如果蝇、水稻、拟南芥)的全基因组测序完成以及基因组学、蛋白质组学和表观遗传学技术的不断完善,解释杂种优势的遗传机理成为亟待解决的问题。果蝇很多基因与人类和哺乳动物的具有同源性,故常作为经典模式动物广泛应用于生物学和医学等相关领域。而且果蝇具有饲养成本低、表型区分明显、易大批量繁殖、试验周期短及效率高等优点,一直是研究者们研究杂交与近交的重要材料。本研究利用果蝇品系间杂交模拟畜禽生产中二元杂交,针对体重性状的杂种优势利用RNAseq及生物信息学分析方法对其遗传机理进行相关研究,以期为畜禽生产中体重杂种优势的探究提供新思路。
1 材料与方法
1.1 实验材料
1.1.1 实验动物 本实验选取5个黑腹果蝇突变品系,分别是极紫眼(dp)、黑檀体(e)、白眼(w)、残翅(vg)和黄体(y),均由中国农业大学动物科技学院模式动物实验室提供。
1.1.2 试剂与仪器 高精度电子天平(德国Sartorius AG)、高速电动组织研磨杵(北京市天根生化科技有限公司);无水乙醇(国药集团化学试剂有限公司)、Trizol(美国Invitrogen公司)、异丙醇(北京市化学试剂有限公司)、氯仿(北京市化学试剂有限公司)、DNA 2000 Marker(北京市欣汇天科技有限公司)。
1.2 实验方法
1.2.1 果蝇杂交的设计 分别在5个果蝇品系中随机选取雌雄果蝇各10只,10 ♂×10 ♀各自纯繁一代(G代)后开展5×5完全双列杂交实验,每个杂交组合选择2只雄蝇与 5只雌蝇交配,得到的后代中杂种记为F,纯种记为G,各组合杂交方式见表1。
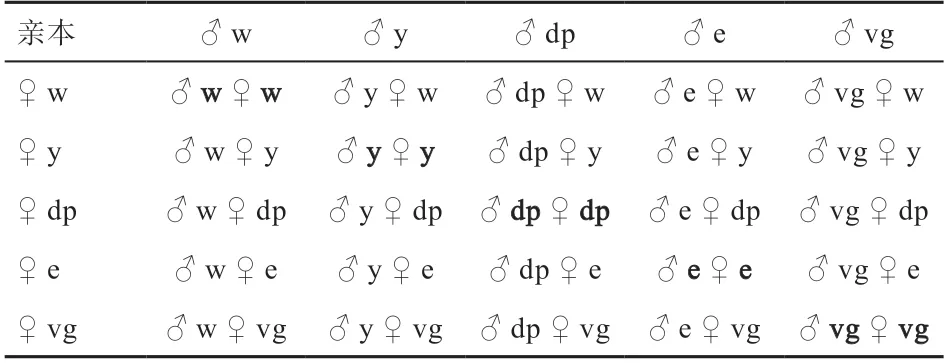
表1 5×5双列杂交表
整个杂交过程共做2次重复实验,上述所有杂交实验中,每个杂交组合均有3个有效重复,最终体重值为3个有效重复的均值。雌蝇均是取羽化8 h内的处女蝇进行后续实验。
1.2.2 体重杂种优势率的计算及检验 杂种优势是指杂种群体均值高于亲本群体均值的部分,一般用杂种优势率(H %)来表示。H%=(杂种均值-双亲均值)/双亲均值×100%。
本实验中F体重杂种优势的计算公式:H%=(F雄性体重均值—两亲本系雄性的体重均值)/双亲本系雄性的体重均值×100%。
雌果蝇羽化8 h内交配后即可产生受精卵,其体重会随着受精卵的发育、成熟而增大。为排除其他因素对体重的影响,本研究只记录雄性果蝇的体重。一次性称取10只雄果蝇的重量取平均值作为1个测量值,每个测量值称取3个有效平行以确保实验数据的准确性。
1.2.3 秩相关计算 本研究2次实验的秩相关计算公式:

式中,d为每对观察值所对应的秩次之差;n为对子数。
1.2.4 果蝇总RNA的提取 采用Trizol法提取果蝇总RNA,通过Nanodrop超微量核酸蛋白测定仪和琼脂糖凝胶电泳测定总RNA的浓度和纯度,整个实验过程均在冰上进行,提取检测完成后于-80℃保存。
1.2.5 果蝇转录组数据生物信息学分析 经核酸蛋白测定仪纯度检测及凝胶电泳检测皆合格的RNA样品交由北京诺禾致源生物信息科技有限公司测序,质检合格后构建cDNA文库,然后基于Illumina HisSeq测序平台对文库进行测序。使用HISAT软件将所得样品转录组数据与Ensembl数据库中的黑腹果蝇参考基因组进行比对;用StringTie软件对每个样本转录本进行组装和量化;基于DESeq2软件分析差异表达基因和差异表达转录本。使用DAVID6.8 (https://david.ncifcrf.gov/tools.jsp)对差异表达基因进行GO和KEGG分析。
2 结果与分析
2.1 2次重复实验体重杂种优势率及检验
2.1.1 2次重复实验G雄性体重 为排除其他因素对体重的影响,本研究只记录雄性果蝇的体重。利用秩相关公式计算得r=1.000,查表知n=5时,显著性水平为0.05的临界值为1.000,该秩相关系数极显著。即G雄性2次排队结果重复性高(表2)。
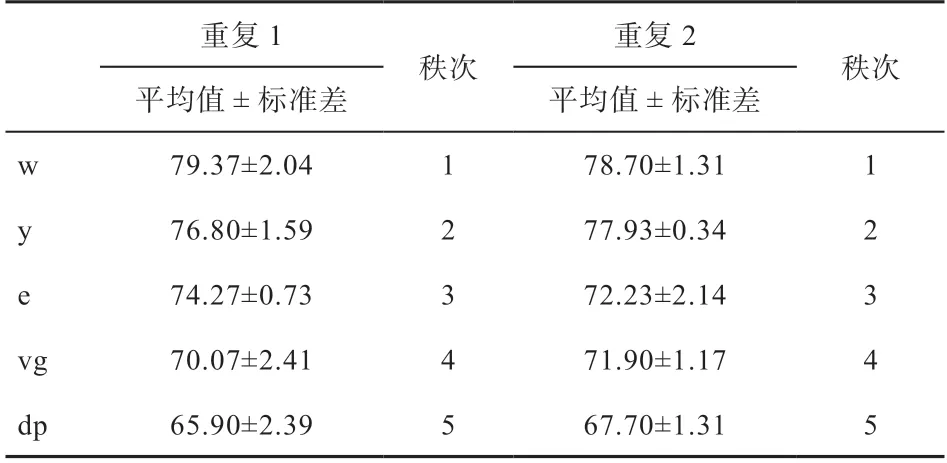
表2 G0雄性单只果蝇体重 μg
2.1.1 2次重复实验F代杂交雄性体重 结合该表所得数据,利用秩相关公式计算得秩相关系数r=0.681,根据Spearman秩相关系数界值表查得n=20时,显著性水平为0.05的临界值为0.447,因此该秩相关系数显著,即F体重杂种优势率2次重复排队结果一致性较好。杂种优势的显著性检验结果见表3,y♂vg♀、dp♂w♀、♂w♀e3个组合体重杂种优势率最高且差异极显著,而vg♂dp♀、dp♂y♀、♂w♀dp 3个组合体重杂种优势最低且差异不显著。故根据秩相关和显著性检测筛选得出3个体重高杂种优势组合(y♂vg♀、dp♂w♀、♂w♀e)和3个体重低杂种优势组合(vg♂dp♀、dp♂y♀、♂w♀dp)。
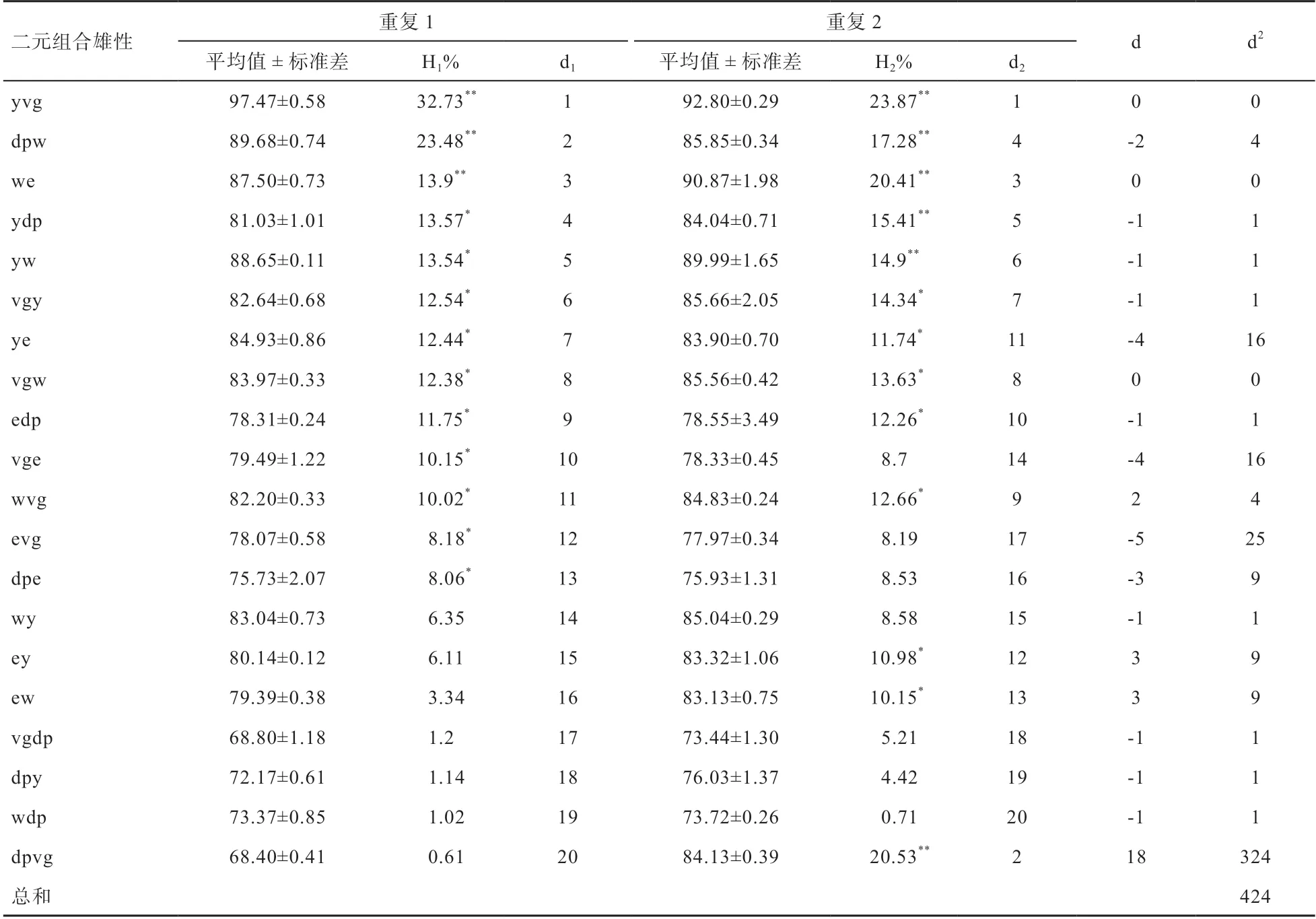
表3 F1代雄性单只果蝇体重 μg
2.2 RNA质量检测和RNA-seq相关性检测 所有提取的RNA样品经过Nanodrop超微量核酸蛋白测定仪检测后,OD/OD和OD/OD都在2.0左右。测序样品的样品浓度都在150 ng/μL以上,完整度(RIN)均在7.0上下(昆虫的检测标准RIN≥6.5即可),质量均为A,检测结果中各项指标均符合质量要求。琼脂糖凝胶电泳检测后的结果见图1,所有RNA样品的3个条带都清晰可见,且18 S较亮,28 S亮度较弱,符合果蝇RNA条带的质量检测要求,无明显拖尾弥散的降解现象,质量较好,可以用于后续实验。
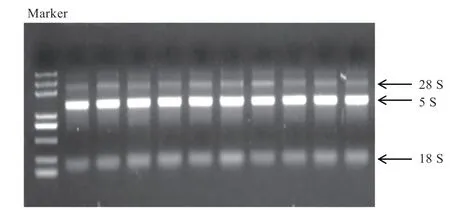
图1 黑腹果蝇RNA凝胶电泳图
本研究共设2个生物学重复,应用Pearson相关系数进行检验,得到任意样品2个生物学重复的绝对系数都在0.929~0.999,说明两重复相关性较高,可用于后续分析。
2.3 高杂种优势组合与低杂种优势组合间的差异表达基因功能分析 经DESeq2分析得到体重高杂种优势3个组合的差异表达基因共有2 659个,体重低杂种优势3个组合的差异表达基因有415个。对体重高杂种优势3个组合和低杂种优势3个组合的差异表达基因取交集,共有275个相同的差异表达基因(图2)。

图2 体重的高杂种优势组合和低杂种优势组合的差异表达基因
分别对高杂种优势组和低杂种优势组特异的差异表达基因进行GO功能聚类和KEGG富集分析,根据值对富集到的结果进行显著性排序(<0.05),选取显著性排名前30的结果作图,GO功能聚类分析结果见图3和图4,KEGG富集分析结果见图5。富集结果表明,杂种优势低组合特有的差异表达基因所富集通路较少;杂种优势高组合特有的差异表达基因均显著富集到糖类代谢、脂类代谢通路以及高杂种优势组合聚类显著且低杂种优势组合没有的通路,如谷胱甘肽代谢通路(Glutathione Metabolism Pathway)和抗坏血酸和醛酸代谢通路(Ascorbate and Aldarate Metabolism Pathway)等通路。
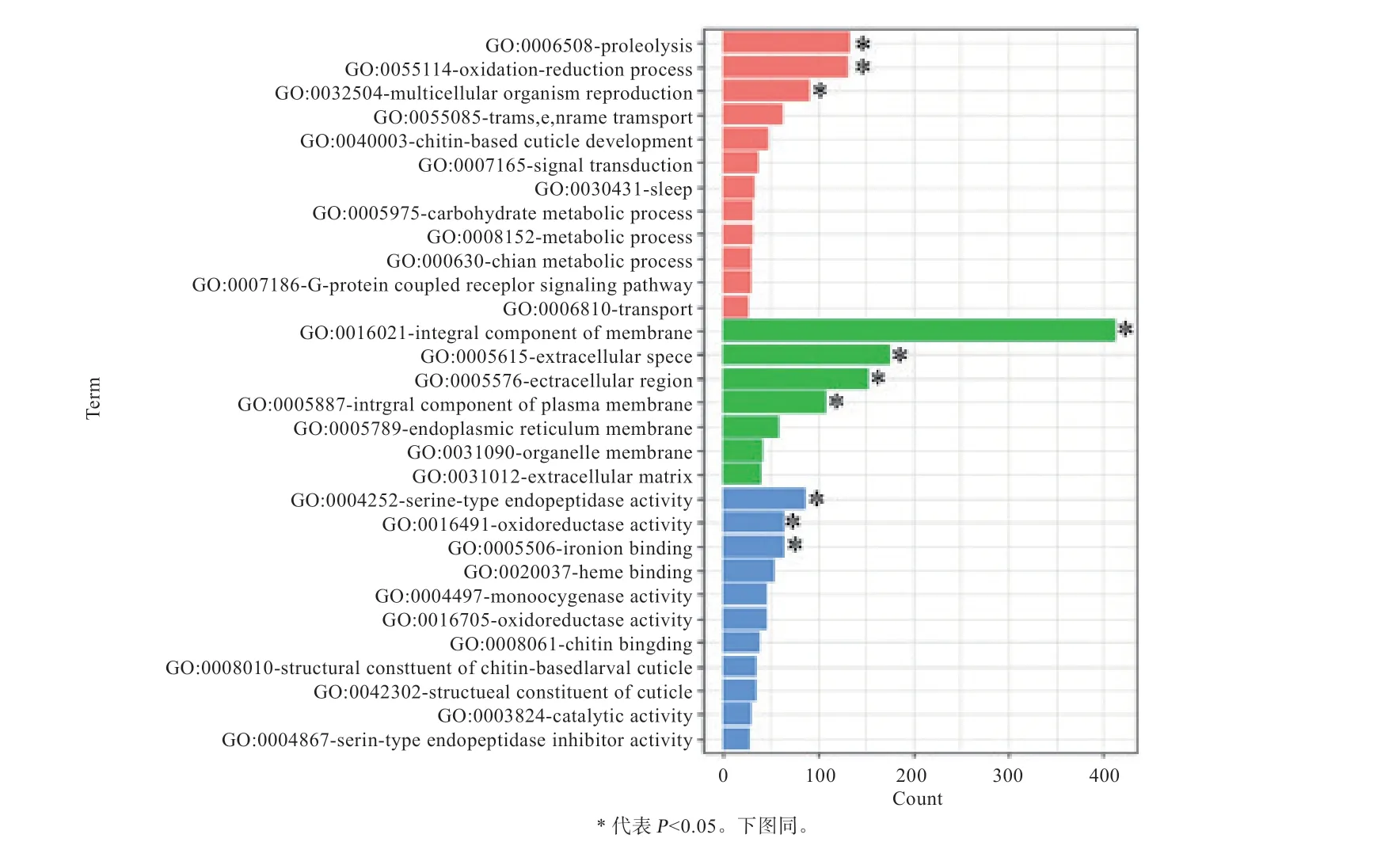
图3 体重杂种优势高组特有的2384个基因的GO功能聚类

图4 体重杂种优势低组特有的140个基因的GO功能聚类
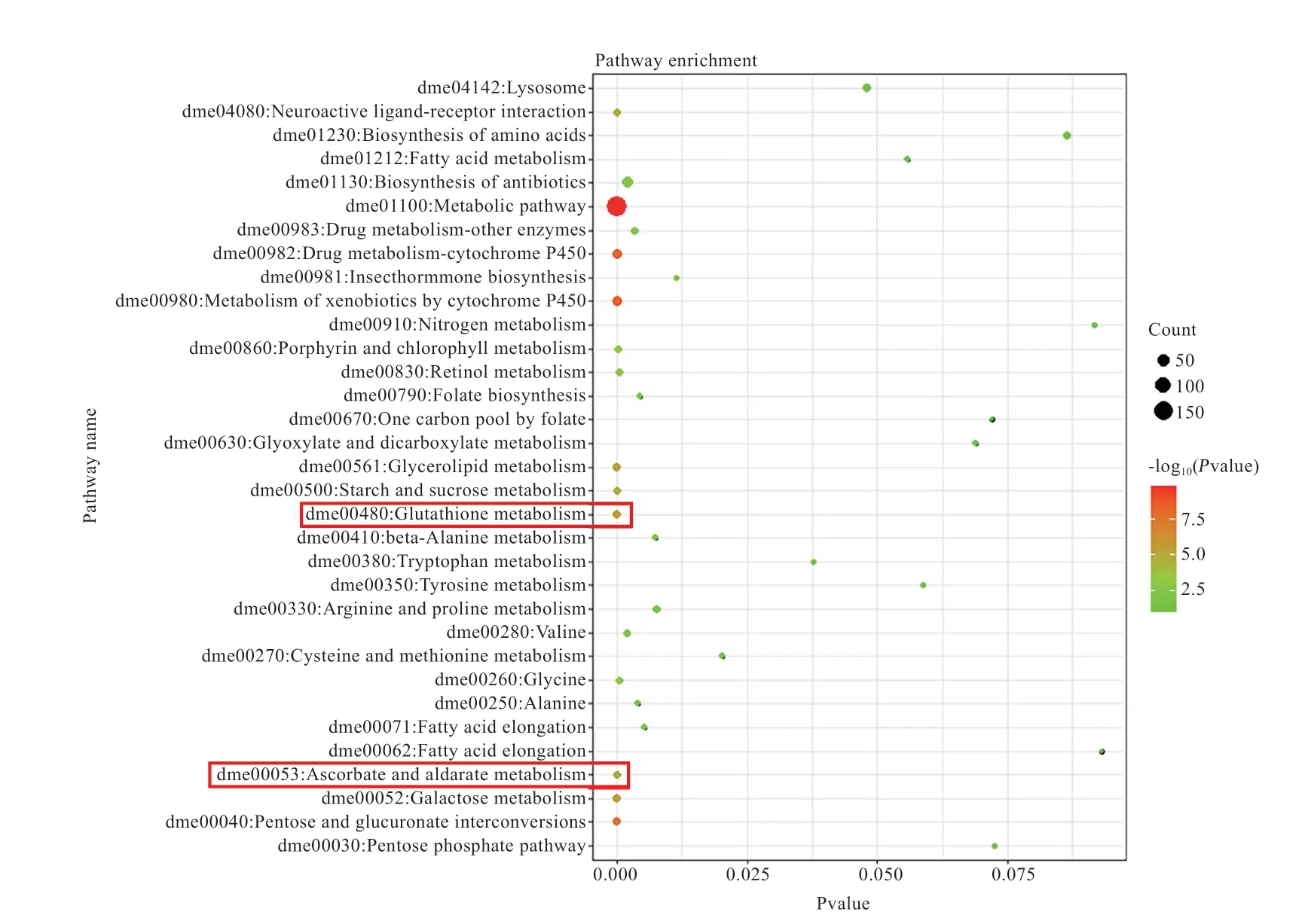
图5 体重杂种优势高组特有的2 384个基因的KEGG富集分析
3 讨 论
在畜禽生产中,直接利用5个品系进行二元配套系杂交会耗费大量的时间、资源、人力和物力,而果蝇凭借其强大的繁殖力和短暂的世代间隔,可以在短时间内实现一个庞大而完整的杂交体系,是理想的探究杂种优势的模型。本研究选取的5个果蝇品系差异较大,可以用来模拟生产水平不同的畜禽品种,例如以不同品系的果蝇体重模拟猪、牛、羊、鸡各配套系的体重,以果蝇9 d内成虫数的指标可以模拟蛋鸡产蛋后孵化数。本研究以果蝇为研究对象,针对体重性状杂种优势展开探讨,为实际生产中与体重性状相关的筛选和预测工作提供理论上的验证和支撑。本实验结果表明,对于果蝇体重性状,大多数杂种优势率都为正值,说明两品系杂交后代的表型值要优于双亲均值,即产生了杂种优势。
本研究对体重高杂种优势组合特有的2 384个基因进行GO分析,在前30项富集条目中有较多基因与糖代谢、脂代谢有关,还发现有少量基因与氧化还原和亚铁离子活动有关,如oxidation-reduction process、iron ion binding等;经富集分析发现,糖代谢通路和脂代谢通路被显著富集到。其中糖代谢通路中包含(编码海藻糖合成酶)、(编码尿苷二磷酸葡萄糖醛酸基转移酶)等基因。海藻糖合成酶可以催化尿苷二磷酸和6-磷酸葡萄糖反应形成海藻糖。海藻糖不仅作为能源和合成细胞成分的葡萄糖存储仓库,还有结构或运输的作用、保护细胞膜和蛋白质免受应力的不利影响,对体重增加和生物抗逆起到重要作用。而存在于许多脊椎动物的肝微粒体中,是重要的生物转化酶之一。可以催化底物与尿苷二磷酸葡萄糖醛酸基结合,使其亲水性增加,利于从毒物和药物体内排出,对体重高优势的形成具有紧密联系。而富集到的脂代谢通路中含有(编码脂肪酸合成酶)、(编码活性调节的细胞骨架蛋白)等基因。FASN是一种基本的代谢酶类,在脂肪酸合成中至关重要。有研究表明,是脂肪沉积性状的候选基因之一,故推测也是造成果蝇体重增加的原因之一,进一步证实了这些代谢通路与体重高杂种优势的形成有紧密联系。
除了糖代谢通路和脂代谢通路被显著富集到,还有2条与谷胱甘肽和抗坏血酸代谢有关通路也被富集到(Glutathione Metabolism Pathway,Ascorbate and Aldarate Metabolism)。谷胱甘肽和抗坏血酸代谢通路在体重高杂种优势组合中被显著富集到而在体重低杂种优势组合中未被富集到,这暗示着体重杂种优势可能与机体抗氧化能力和药物代谢能力有着密切联系。谷胱甘肽几乎存在于机体的每个细胞中,它能帮助维持机体正常的免疫系统功能。谷胱甘肽上的半胱氨酸易于某些药物、毒素和结合,从而具有抗氧化、解毒的作用。抗坏血酸又称维生素C,具有强还原能力,与谷胱甘肽、亚铁离子一同具有抗氧化能力。目前,有关动物毒理效应的研究报道有很多,重金属污染物对动物机体的作用有引起细胞内活性氧过量产生,从而造成细胞内核酸和蛋白质的氧化损伤及膜脂过氧化,且重金属可与抗氧化酶和谷胱甘肽发生作用,影响胞内酶的活性与浓度,干扰体内氧化还原反应,进而影响生物体的生长、发育和繁殖。从上述代谢过程来看,推测这些潜在通路对畜禽生长发育和体重杂种优势有着紧密联系,然而对于差异表达基因的具体调控方式和通路作用机制目前还未知。
4 结 论
本研究以模式动物果蝇为实验对象进行二元杂交,利用RNA-seq对体重性状杂种优势高低两组分别进行测序分析,初步探究杂种优势的分子机理,筛选出了与体重杂种优势相关的潜在候选基因和代谢通路,主要有糖代谢通路、脂代谢通路和氧化还原通路等。
猜你喜欢
学苑创造·A版(2023年10期)2023-11-04 13:14:04
大自然探索(2023年11期)2023-03-01 09:04:36
学苑创造·A版(2022年3期)2022-03-29 23:32:16
蔬菜(2021年7期)2021-11-27 03:45:25
学苑创造·A版(2019年6期)2019-07-11 01:07:39
天然产物研究与开发(2016年6期)2016-06-05 10:29:26
山西农经(2016年5期)2016-02-28 14:24:37
分析测试学报(2015年6期)2016-01-13 06:19:00
集美大学学报(自然科学版)(2015年4期)2015-02-28 01:13:36
茶叶通讯(2014年2期)2014-02-27 07:55:39