
埋深和播种密度对豚草种子出苗及幼苗生长的影响
2022-01-17 08:47:40熊韫琦赵彩云赵相健
生态学报 2021年24期
熊韫琦,赵彩云,*,赵相健
1 中国环境科学研究院, 北京 100012 2 中国合格评定国家认可中心, 北京 100062
外来入侵植物是指从其原生范围迁移到新的生境,并对该地区生物多样性、生态系统结构功能、农林牧渔业生产和人类健康造成严重威胁的植物[1]。种子出苗和幼苗生长是植物生活史中最敏感也是最重要的阶段,影响着植物种群增长、扩散和延续[2],是入侵植物成功定植的关键时期[3],同时受到自身内在因素和外界环境因素的影响。影响种子萌发和幼苗生长的内在因素主要包括种子大小、种子活力、种子成熟度以及种子是否休眠等[4]。其中种子大小是最重要的性状,影响种子萌发、幼苗的建成、个体适合度以及群落更新等过程[5-6]。研究发现较小的种子更易扩散,并且更能保持种子库的持久性[7-8];而较大的种子更能抵抗环境压力,竞争能力更强[9]。因此,对外来入侵植物而言,种子大小严重影响了其传播扩散,以及与本地物种的竞争。
适宜的光照、水分、温度和氧气等是种子出苗的必要环境条件。在一定的埋深范围内,覆土可以给种子提供更适宜的环境条件,如温度、湿度等[10],还可以避免动物的取食[11],从而有利于种子萌发、幼苗出苗及生长。但是埋深过深会抑制幼苗的出苗和存活,因为深埋条件下,种子由于缺乏氧气、光照和温度变化而难以萌发,同时幼苗到达土壤表面的能量需求和时间增加,使得出苗前死亡率增加[12-13]。密度是自然界重要的选择压力之一,播种密度会影响可获得性资源的数量,使得植株间产生竞争,影响植物个体生物量和存活率[14-15]。研究表明,播种密度与入侵植物定殖呈正相关关系,因为高密度的入侵物种会引起资源竞争而排斥本地物种[16-17]。
豚草(Ambrosiaartemisiifolia),又名艾叶破布草或豕草,属双子叶植物纲菊科豚草属一年生草本植物,原产于北美,是一种被世界公认的恶性入侵杂草[18-19]。豚草严重影响了入侵地的生物多样性以及生态系统稳定性,给农业、渔业和牧业等带来巨大经济损失,其致过敏的花粉对公共卫生也造成极大影响,已被列入我国首批入侵物种名单[20-21]。为加强对该入侵物种的控制和管理,对其生活史重要阶段的种子出苗和幼苗生长过程的研究至关重要。目前,国内外对豚草的研究主要集中于豚草的入侵机制[22- 25]、分布和扩散[26- 28]、对环境因子的响应[20, 29- 31]、危害与防治[21, 32]等方面,而鲜少关注豚草出苗和幼苗生长阶段受种子特征、播种深度和密度的影响。外来植物在幼苗阶段的适应性直接影响了其成功入侵的可能性[33],因此,本文以豚草为研究对象,采用盆栽实验,研究了种子大小和埋深/播种密度对豚草种子的出苗和幼苗生长的影响,进而为豚草的防控和管理提供科学依据。
1 材料与方法
1.1 实验地点
实验于2014年4—6月份在北京中国环境科学研究院顺义基地进行,该地区属于温带大陆性半湿润季风气候,年均气温11.5℃,年均降雨量约610 mm,80%降水集中于7—8月,年日照2750 h,年无霜期约为195 d[34]。
1.2 实验材料
豚草种子于2013年收集自顺义基地种植的豚草植株,从生长健壮的植株上摘取不同大小的成熟种子晾晒后置于4°C的干燥条件下保存。实验开始前,分别用10目、12目和14目的土壤筛(孔径分别为2.00、1.66 mm和1.43 mm)将豚草种子按大小分为三组:大(2.00 mm>d≥1.66 mm,L)、中(1.66 mm>d≥1.43 mm,M)和小(1.43 mm>d,S)。
1.3 研究方法
1.3.1实验设计
埋深实验设置3个种子大小(L、M、S)和3个埋深处理(2、4 cm和6 cm),每个处理9个重复。2014年4月21日进行播种,取高27 cm、内径31 cm、地径18 cm的塑料花盆81个,平均分为3组(每组27个)分别播种大、中和小种子;再将每组的27个花盆按9盆1组分为3组,分别对应3个不同埋深处理(2、4 cm和6 cm)。播种时,花盆中先填入部分培养基质,充分浇水后放入豚草种子,再分别铺上厚约2、4或6 cm的湿润土壤以实现不同埋深处理。每个花盆播种10粒同大小等级的豚草种子,试验用种子总计:3种子大小×3埋深处理×9重复×10粒=810粒,培养基质为去除表面凋落物的0—25 cm农田土与腐殖土2∶1混合。
播种密度实验设置3个种子大小(L、M、S)和3个播种密度处理(每盆2、4粒和8粒),每个处理4个重复。2014年4月15日进行播种,取高27 cm、内径31 cm、地径18 cm的塑料花盆36个,平均分为4组(每组12个)分别播种大、中和小种子;再将每组的12个花盆按4盆1组分为3组,分别对应3个不同播种密度处理(每盆2、4、8粒)。播种时,花盆中先填入部分培养基质,充分浇水后每盆放入2、4或8粒豚草种子,再填入约2 cm厚的湿润土壤覆盖种子,培养基质为去除表面凋落物的0—25 cm农田土与腐殖土2∶1混合。试验用种子总计:3种子大小×4重复×(2粒+4粒+8粒)=168粒。
1.3.2生长特征记录
两个实验均分别进行了50 d,播种后每天浇水保持土壤湿润,看见子叶则视为出苗。第50天记录每个花盆的豚草植株数目并进行收获,收获前分别用直尺和游标卡尺测量每株豚草幼苗的株高、基径,然后将植株挖出洗净后用直尺测量根长,随后将植株地上和地下部分分开后置于80℃烘干48 h,用千分之一天平称重。
本实验记录了幼苗数、株高、根长、基茎、每盆总地上生物量和每盆总地下生物量;并计算出苗率(出苗数量/播种数量)、每盆平均株高、每盆平均根长、每盆平均基径、每盆总生物量(每盆总地上生物量+每盆总地下生物量)、单株生物量(每盆总生物量/出苗数量)、单株地上生物量(每盆总地上生物量/出苗数量)、单株地下生物量(每盆总地下生物量/出苗数量)以及根冠比(每盆总地下生物量/每盆总地上生物量)。
1.4 数据分析
先将相关数据进行对数转换(y=lg(x+1)),然后采用一般线性模型进行双因素方差分析(Two-way ANOVA)检验种子大小和埋深/播种密度及其相互作用对种子萌发和幼苗生长的影响。如若差异显著,则采用Duncan多重比较检验每个处理的差异性(α=0.05)。所有数据采用Excel 2016进行统计,SPSS 25.0进行分析处理。
2 结果与分析
2.1 种子大小和埋深对豚草种子出苗和幼苗生长的影响
种子大小对豚草的株高影响显著(P<0.05),对基径的影响极显著(P<0.01);除株高和根长外,埋深对其他指标均有极显著影响(P<0.01);种子大小和埋深的交互作用仅对基径的影响达到显著水平(P<0.05)。(表1)。
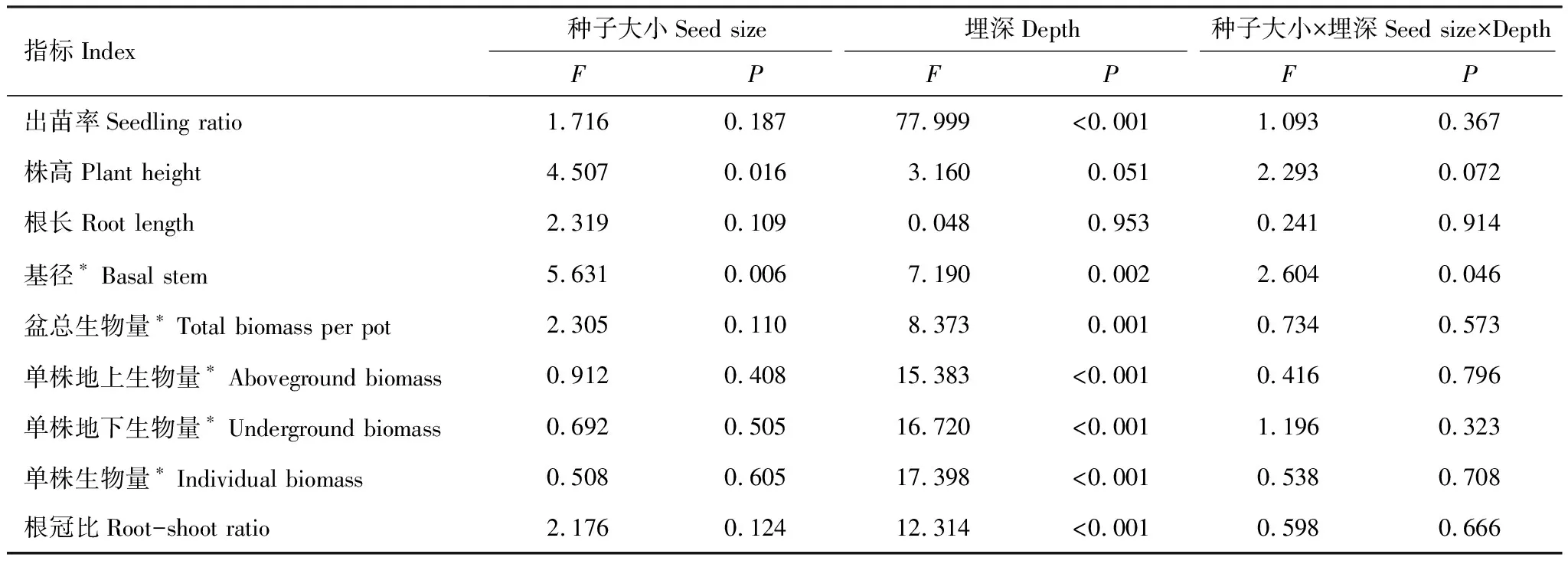
表1 种子大小和埋深对豚草种子出苗和幼苗生长影响的双因素方差分析
所有大小种子的出苗率均在2 cm时最大,达到47.8%—52.2%,并随埋深增加而显著下降;不同大小种子的出苗率在不同播种埋深间无显著差异。不同大小种子豚草的株高均在埋深2 cm时最高,大、中种子的株高在不同埋深间差异不显著,小种子的株高在埋深2 cm时显著高于4 cm时46.2%。在埋深2 cm和4 cm时,株高随种子大小增加而逐渐增加,而埋深6 cm时,中种子的株高最高;其中埋深4 cm时,大种子的株高较小种子显著增加了1.25倍,其余埋深处理下,豚草的株高在不同种子大小之间没有显著差异。不同大小种子的基径在2 cm埋深时最大,仅小种子的基径在不同埋深间差异显著,且埋深2 cm时的基径显著大于埋深4 cm时1.6倍;除埋深4 cm外,基径在不同大小种子间均无显著差异,埋深4 cm时,基径随种子大小增加而显著增加,大种子的基径分别为中、小种子的1.63、2.97倍(图1)。
中种子的每盆总生物量在不同埋深间差异显著,且埋深2 cm时较埋深6 cm时显著增加了26.3%,其余大小种子的每盆总生物量在不同埋深间的差异未达到显著水平;不同埋深下较大种子的每盆总生物量均最大,其仅在埋深4 cm时显著高于小种子39.0%,其余埋深下不同种子大小间无显著差异。
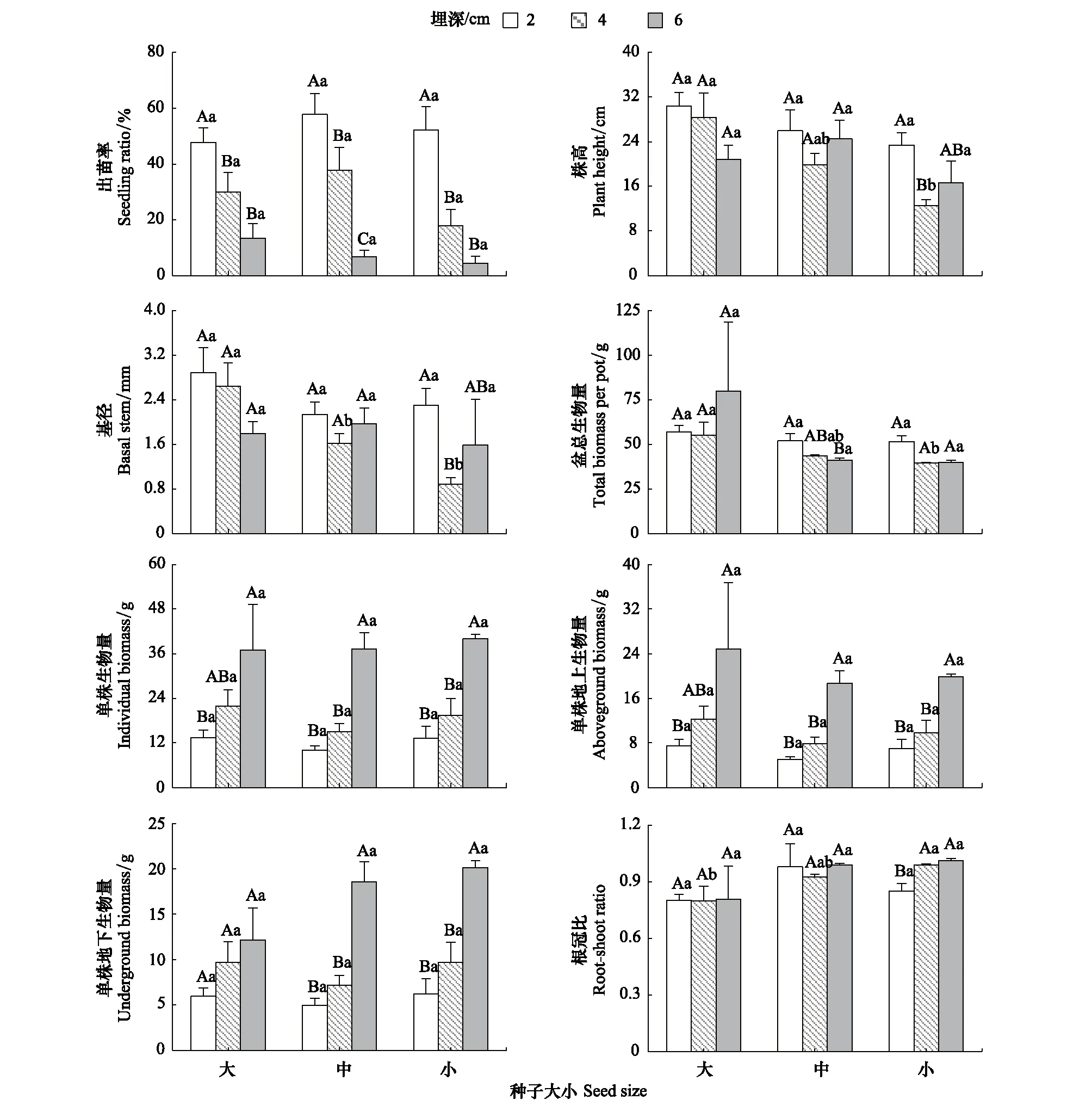
图1 不同种子大小在不同埋深处理下各指标的差异Fig.1 The difference of parameters among different seed size of A. artemisiifolia with different burial depth不同大写字母和不同小写字母分别表示同一大小种子的不同埋深间以及同一埋深下不同大小种子间差异显著(P<0.05)
随埋深增加所有大小种子的单株生物量、单株地上生物量和单株地下生物量均显著增加;不同埋深处理下,该3个指标在不同大小种子间没有显著差异。大种子的单株生物量和单株地上生物量在埋深6 cm时分别显著高于埋深2 cm时2.75倍和2.30倍,中、小种子的单株生物量和单株地上生物量在埋深6 cm时分别比埋深4 cm时显著增加了1.48、1.06倍和1.38、1.03倍。大种子的单株地下生物量在不同埋深间差异不显著,中、小种子的单株地下生物量在埋深6 cm时分别比埋深4 cm时显著增加了1.58倍和1.08倍(图1)。
不同大小种子的根冠比均在埋深6 cm时最大,为0.80—1.01,其中小种子的根冠比随埋深增加而逐渐增加,且埋深6 cm时显著高于埋深2 cm时18.7%,其余大小种子根冠比在不同埋深间差异不显著。在4 cm和6 cm埋深下小种子的根冠比最大,在2 cm埋深下中种子的根冠比则最大,不同大小种子间的根冠比差异仅在埋深4 cm时达到显著水平,且小种子的根冠比显著高于大种子23.8%(图1)。
2.2 种子大小和播种密度对豚草种子出苗和幼苗生长的影响
种子大小仅对出苗率具有显著影响(P<0.05),对其余所有指标的影响均未达到显著水平(P>0.05);播种密度对豚草幼苗的每盆总生物量、单株地上和地下生物以及单株总生物量有极显著影响(P<0.01);种子大小和播种密度的交互作用对所有指标均无显著影响(P>0.05)(表2)。
豚草种子在不同播种密度下均具有较高的出苗率,达到67.9%—100%,且播种密度对不同大小种子的出苗率无显著影响;除播种密度为每盆2粒外,出苗率受种子大小的影响不显著,在每盆2粒的播种密度下,出苗率随种子大小增加而逐渐增加,且大种子显著高于小种子36.8%(图2)。
不同大小种子豚草的每盆总生物量均在播种密度为8粒每盆时最大,其中大种子在播种密度为每盆8粒时显著高于每盆4粒时16.3%,中种子的每盆总生物量在不同播种密度之间无显著差异,小种子的每盆生物量在播种密度为每盆8粒时较每盆2粒时显著增加了9.9%,;种子大小对不同播种密度下的盆总生物量没有显著影响。所有大小种子的单株总生物量、单株地上生物量和单株地下生物量均随播种密度增加而显著降低;而种子大小对该3个指标的影响均未达到显著水平。其中,大、中、小种子在播种密度为每盆2粒时的单株生物量较每盆8粒时分别显著增加了2.19、2.42倍和2.91倍;单株地上生物量较每盆8粒时分别显著增加了2.14、2.27倍和2.75倍;单株地下生物量较每盆8粒时分别显著增加了2.25、2.58倍和3.07倍(图2)。
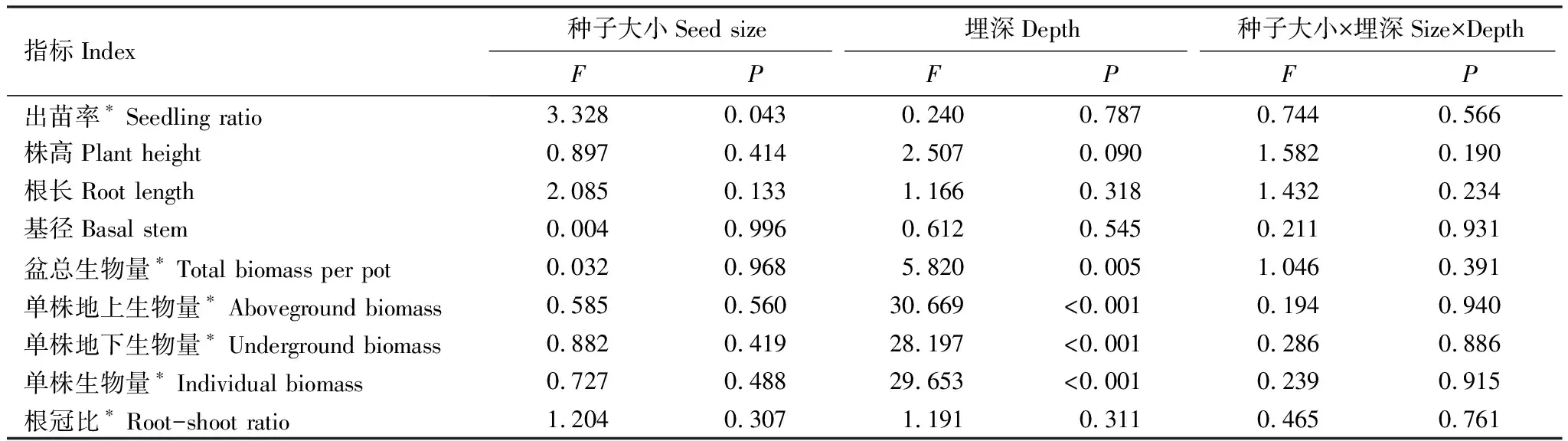
表2 种子大小和播种密度对豚草种子出苗和幼苗生长影响的双因素方差分析

图2 不同种子大小在不同播种密度处理下各指标的差异Fig.2 The difference of parameters among different seed size of A. artemisiifolia with different planting density不同大写字母和不同小写字母分别表示同一大小种子的不同播种密度间以及同一播种密度下不同大小种子间差异显著(P<0.05)
3 讨论
3.1 种子大小和埋深/播种密度对种子出苗的影响
大量研究表明种子大小对出苗有着显著影响[2, 12, 35-36],一般而言,大种子具有更多的能量以支撑种子萌发后出苗,而较小的种子更可能在萌发后,因无法到达土壤表面而死亡[37]。但本研究中,种子大小对出苗率无显著影响,可能是由于埋深实验中选择的豚草种子大小等级差异较小,使得种子大小之间的差异不能明显地表现出来,今后实验应加大豚草种子大小的分级差异。本研究发现播种深度对豚草种子的出苗率存在极显著影响,从埋深2 cm时的47.8%—52.2%下降到埋深6 cm时的4.4%—13.3%。前人的研究也证实了豚草种子的出苗率随埋深的增加而减少这一趋势[38-39]。因为播种的深度会影响种子周围的水分、光照、温度以及氧气含量等环境因素,埋深越深,不宜的环境会抑制种子的萌发和出苗,且出苗前死亡率也随之升高[40- 42]。对其他入侵植物,如白花鬼针草(Bidensalba)、假臭草(Praxelisclematidea)和胜红蓟(Ageratumconyzoides)[43]等的研究均证实了埋深的增加会抑制种子的萌发和出苗。因此,通过深耕等方式将种子深埋入土,降低豚草的出苗率,从而达到一定的防除效果。
另一方面,本研究还发现播种密度对豚草种子出苗率的影响不显著,这说明豚草的出苗不受播种密度的影响,无论在何种密度下,豚草的出苗率均能达到较高水平,这有利于它在不同生境中的传播和扩散,形成较高竞争力,这是入侵植物的一大特征[44]。此外,密度实验中,种子大小对豚草的出苗率影响显著,且总体上中种子的出苗率较高,与埋深实验结果的不同可能是因为两个实验中,豚草种子的大小等级差异较小,加之埋深和播种密度条件不同造成的误差。
3.2 种子大小和埋深/播种密度对幼苗形态特征的影响
种子大小可以影响早期幼苗的建成,从而影响植物在异质生境中的定殖[11, 45]。本研究发现埋深处理下,种子大小显著影响豚草幼苗的株高和基径,整体上较大种子产生的幼苗的株高和基径高于小种子。这是由于较大的种子由于储存的营养物种更多,资源获取比小种子更有优势,更有利于幼苗的生长。许多其他研究结果也与本研究结果相同,例如Mao等[9]对中国山东半岛北部沿海外来植物日本黑松(PinusthunbergiiParl.)种子发芽和幼苗早期生长的研究发现,大种子和中种子的幼苗高度和基径均高于小种子,即较大的种子比小种子在生长中更有优势。而播种密度处理中,株高、根长、基径均不受种子大小的影响,这可能是因为播种密度实验中种子埋深较浅,削弱了种子大小的影响。
播种埋深影响幼苗光照及养分等营养物质的摄取,从而影响幼苗的生长。本研究发现豚草幼苗的株高和基径受埋深影响显著,且在浅埋深下最大,但仅小种子的株高和基径在不同埋深下的差异达到显著水平。这可能是由于在较深的播种深度下,种子将分配更多的能量用于萌发和出苗,而用于幼苗早期生长的能量减少,同时埋深更深的种子所需的出苗的时间更长,从而导致出土后的生长期缩短,使得幼苗生长受限,并且小种子更易受埋深的影响[38, 46]。对辽东栎(Quercuswutaishanica)[11]、黑松(PinusthunbergiiParl.)[9]、泡泡刺(Nitrariasphaerocarpa)[12]等的研究,同样也证实了较浅的埋深下幼苗的株高更大。另一方面,豚草幼苗的株高、根长、基径受播种密度的影响不显著。说明播种密度对豚草早期幼苗的生长影响不大,因此它能够在不同密度环境下生长定殖,促进其在异质生境中的成功入侵。
3.3 种子大小和埋深/播种密度对幼苗生物量的影响
在两个实验中,种子大小对每盆总生物量、单株总生物量、单株地上生物量和单株地下生物量的影响均未达到显著水平,而埋深和播种密度对以上4个指标均具有极显著影响,表明埋深和播种密度对豚草幼苗生物量的影响大于种子大小的影响。种子大小对根冠比的影响仅在埋深4 cm时达到显著水平,且根冠比随种子大小的增加而显著下降,说明更小的种子会将生物量更多的分配给根以促进幼苗的生长。
随埋深增加,豚草幼苗的单株总生物量、单株地上和地下生物量均呈显著增加趋势,而每盆总生物量在不同埋深下无显著差异。埋深对豚草幼苗生物量的促进关系可能是由于较深的埋深更有利于根获取土壤养分促进地下部分的生长,吸收的养分运送到地上部分进而促进了茎和叶的生长。虽然豚草幼苗的单株生物量随埋深增加而增加,但是越深的播种深度意味着越低的出苗率,因此每盆总生物量在不同埋深间差异不大。豚草幼苗的根冠比在较深埋深下更大,表明较深的埋深更有利于根的生长,豚草幼苗将更多的能量分配给地下部分,从而导致根冠比的增加,这与徐玲玲等[47]对枫杨(Pterocaryastenoptera)种子的研究结果相似,这是植物体应对埋深的一种生态适应[48],且这种影响在小种子中表现更为明显。豚草的这种通过调整生物量分配以适应不同埋深环境的策略,促进了其在不同生境中的入侵。
播种密度会影响幼苗的种内竞争,密度越高,水分、营养等资源的竞争越激烈,同时幼苗叶片相互遮挡,影响光照的获取,从而影响光合作用,抑制幼苗的生长[15, 49]。本实验中,豚草幼苗的单株总生物量、单株地上和地下生物量均随播种密度的增加而显著减少,而每盆总生物量总体上随播种密度增加而呈上升趋势。随播种密度的增加,由于土壤营养物质有限,幼苗为了获取养分而产生激烈的竞争,从而导致单株幼苗的生长受限,生物量积累受到抑制。Leskovšek等[29]的研究也与本研究结果一致,即播种密度对豚草生物量积累具有负效应。但是播种密度对每盆总生物量的促进作用表明高密度不会抑制豚草种群的整体生长。在野外,入侵植物豚草通常会形成高密度的单优群落,阻碍其他本地植物的生长,这可能是豚草成功入侵的原因之一。由于竞争可以抑制豚草的生长,因此可以选择竞争能力强的本地植物,与豚草进行养分和光能等资源的竞争,使得本地种获得更大的生物量,使入侵植物豚草处于不利地位。这种替代控制方法是抵御外来杂草入侵扩张的有效途径之一[50]。
综上所述,种子大小对豚草出苗和生长的影响均较小,但总体上较大的种子的生长比较小的种子更具有优势。本研究中豚草种子大小的分级差异较小,可能导致了种子大小的影响效应不明显,因此今后研究中应加大种子大小的分级差异。豚草种子的出苗主要受到埋深的影响,较浅的埋深更有利于豚草种子的出苗;不同播种密度下,豚草种子的出苗率均较高,达到67.9%—100%,有利于它在不同生境中的传播和扩散。豚草幼苗生长受埋深和播种密度影响更大,且小种子更易受埋深和播种密度的影响。浅层埋深幼苗的株高和基径更大,埋深对豚草幼苗生物量具有促进作用。播种密度会影响豚草幼苗的种内竞争,抑制其生物量积累。由此可见,豚草在浅层埋深处理下能够保持较高的出苗率,拥有较大的株高和基径;播种密度较小的条件下,豚草植株生物量更大,即豚草在浅埋深、低密度生境中更容易入侵成功。因此,为有效管理和防治豚草,应在春耕前对土壤进行深翻,增加种子的埋深,减少豚草的出苗率;还可以采取替代控制,种植竞争能力强的本地植物,抑制豚草的生长和扩散。
猜你喜欢
草食家畜(2022年5期)2023-01-02 02:53:26
辽宁农业科学(2021年5期)2021-11-03 07:19:30
四川林业科技(2019年5期)2019-07-07 07:14:30
安徽农学通报(2019年1期)2019-02-14 02:25:08
湖北林业科技(2019年6期)2019-01-06 02:14:29
新农业(2018年6期)2018-04-18 07:30:36
湖北林业科技(2017年2期)2017-05-25 18:44:32
新农业(2016年19期)2016-08-16 12:00:18
现代农业(2016年5期)2016-02-28 18:42:34
农村农业农民·B版(2014年6期)2014-08-08 21:17:51
