
长白山林线过渡带树岛土壤微生物群落结构和生态功能
2022-01-16 05:35:28马锐骜王志瑞戴冠华于大炮
生态学报 2021年24期
马锐骜,吴 辉,王志瑞,戴冠华,于大炮,姜 勇,李 慧,*
1 中国科学院沈阳应用生态研究所,森林生态与管理重点实验室, 沈阳 110016 2 中国科学院大学, 北京 100049
林线过渡带是森林和其他植物地理群落之间的交错地带,包括从郁闭森林线(郁闭度大于0.2且树高大于3 m)到树种线(树高不足2 m 的矮曲木生长的上限)的区域[1]。在这一特殊地带,乔木处于生长极限临界状态,生态系统结构、功能和生态过程在很小的海拔梯度内发生剧烈变化,生物多样复杂,对全球气候变化及人类活动干扰均极为敏感,历来被生态学家视为生态系统脆弱区和全球变化预警区[2- 3]。在林线过渡带内,由于局部地形条件的差异,具有遮蔽的微地形提供了温暖、少风、高湿的微气候,形成斑块状或条带性的不连续树木分布集群,称为 “树岛”[4]。树岛内生存的树木通常能达到与较低海拔郁闭森林同样的高度和胸径,因此揭示树岛这一特殊生境的生态特征及其形成机制,对于预测未来气候变化下林线动态具有重要意义。
之前对于林线过渡带的研究大多关注地上植被群落结构特征[5- 8],对于生物地化循环过程的重要参与者——土壤微生物群落却知之甚少。土壤微生物是土壤养分循环的重要驱动者,直接参与碳、氮、硫、磷等养分的物质循环和能量代谢,并且与地上植被存在着紧密的养分交流和生态关联[9- 10],对维持生态系统结构和功能具有重要贡献。在林线过渡带同一海拔高度,有的生境能适宜高大乔木生长,形成不同规模的树岛;而有的生境则形成高山草甸或苔原等开阔地带。树岛内外温度、辐射、地上植被群落组成、凋落物和根系分泌物、土壤理化性质等环境因子均存在巨大差异,必然选择不同的土壤微生物群落;反之,土壤微生物群落结构和功能也将影响土壤养分循环,进而影响地上植被的生长和树岛扩张动态。
长白山北坡岳桦带主要分布在海拔1700—2000 m,在海拔1950 m左右形成急变型林[11](图1)。从海拔1900 m到2030 m为林线过渡带,在过渡带内,岳桦总体呈斑块状分布,形成大小不同规模的树岛。由于气候变暖,长白山北坡岳桦林线在过去160年内上移了80米,树岛面积不断扩张[12]。本文以过渡带内较大的树岛(2.7 hm2)为研究对象(图1),通过测定了土壤养分、土壤微生物呼吸和胞外酶活性,并采用宏基因组测序分析了土壤微生物群落结构组成和功能基因丰度,拟揭示树岛这一特殊生境的土壤微生物群落结构特征和潜在生态功能。同时,通过与同海拔的开阔区生境进行对比,从土壤养分和土壤微生物学角度,阐明树岛扩张的可能驱动机制。
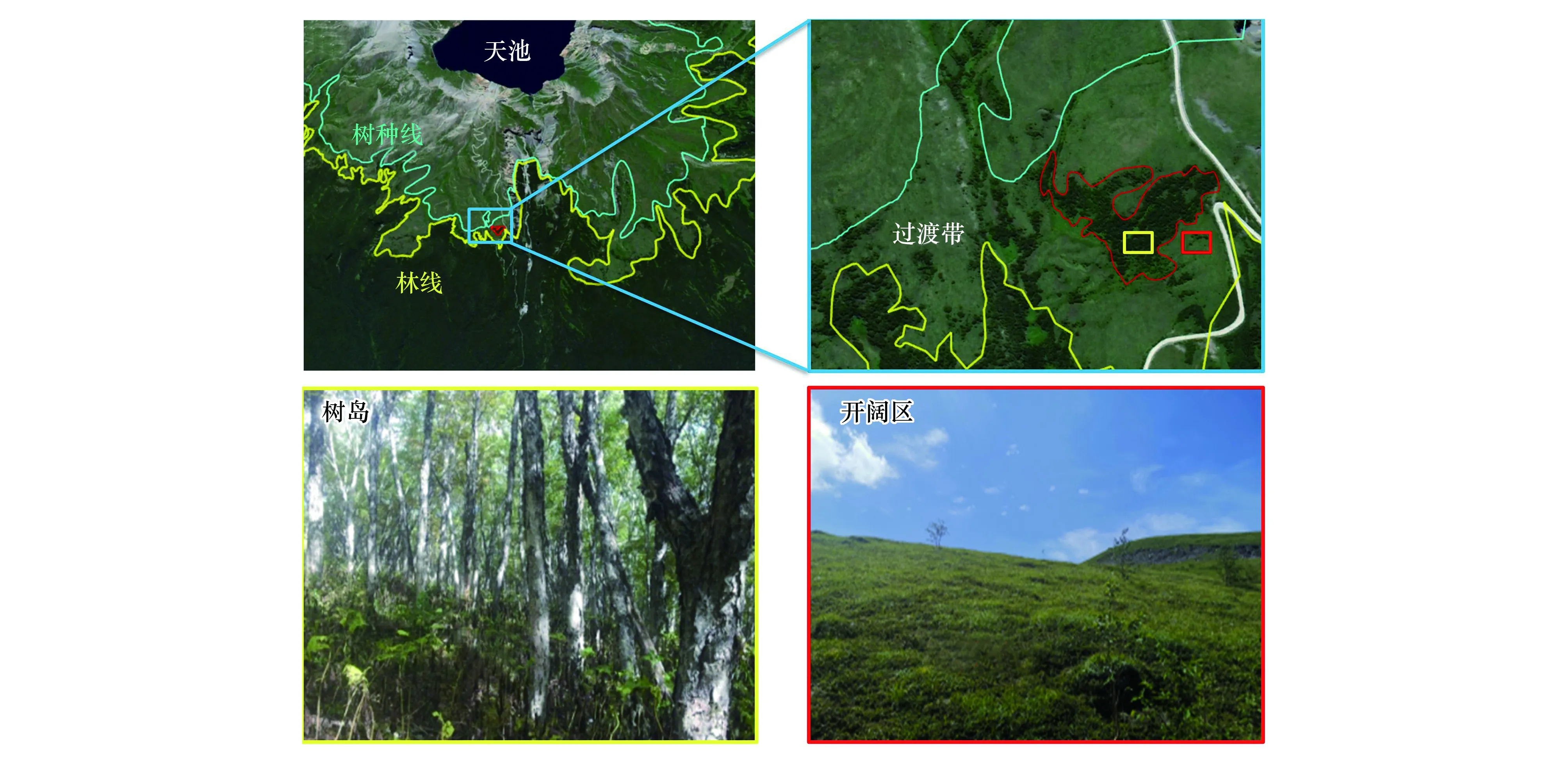
图1 长白山林线过渡带开阔区与树岛相对位置示意图 Fig.1 Schematic diagram of the open area and tree island in the treeline transition zone of Changbai Mountain treeline
1 材料与方法
1.1 研究区域简介和土壤样品采集
长白山属于温带大陆性山地气候,夏季温暖多雨,冬季寒冷漫长。山顶和山下气候差异大,年平均降水量为750 mm,年平均气温分别为2.8℃和7.3℃。降水多集中在夏季(6—8月份)。本研究选择长白山北坡岳桦林-苔原过渡带(42°04′—42°05′N,128°07′—128°08′E)为研究对象,海拔范围2000—2030 m。如图1,在这一林线过渡带内,树岛中以乔木型岳桦(Betulaermanii)、牛皮杜鹃(Rhododendronchrysanthum)、小叶章(Deyeuxiaangustifolia)、笃斯越桔(Vacciniumvitis-idaea)为主。开阔区如图1D,植物主要以灌木型岳桦(Betulaermanii)、牛皮杜鹃(Rhododendronchrysanthum)、小叶章(Deyeuxiaangustifolia)、笃斯越桔(Vacciniumvitis-idaea)为主。2019年7月,分别采集树岛与同海拔高度开阔区土壤,每个生境选取3个20 m×20 m样方作为重复。在每个样方内,选取中心和四个角共5个样点采集土壤,混合成为一个土壤样品。采集土壤时,先去除凋落物层,采用直径3 cm土钻钻取表层(0—10 cm)土壤样品,相当于有机质层(O层)。土壤样品去除根系和石块后分为三份:一份自然风干,用于测定土壤理化性质;一份4℃条件保存,用于测定土壤呼吸、微生物生物量和土壤酶活性;一份于-20℃保存,用于提取微生物基因组并进行后续测序分析。
1.2 土壤理化性质测定
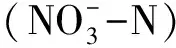
1.3 土壤微生物量测定
微生物量碳(Microbial biomass carbon, MBC)和微生物量氮(Microbial biomass nitrogen, MBN)微生物量磷(Microbial biomass phosphorus, MBP)采用氯仿熏蒸-浸提法,微生物量碳氮用0.5 mol/L K2SO4溶液浸提后,滤液用TOC分析仪(High-TOC, Elementar Vario MACRO cube, Germany)进行测定。微生物量磷采用0.5 mol/L NaHCO3浸提,钼锑抗比色法测定。
1.4 土壤呼吸测定
土壤基础呼吸(Basal respiration, BR)测定:称取15 g鲜土,置于150 mL三角瓶中,在25 ℃培养箱静置24 h。将室内温度调节至25 ℃,向三角瓶中泵入无CO2的空气,充气2.5 min后用红外气体分析仪(Li-COR 820)测定CO2的浓度和流速。底物诱导呼吸(Substrate induced respiration, SIR)的测定:25 ℃培养24 h后,向土壤中加入2 mL浓度为6%的葡萄糖溶液,静置1.5 h,测定其呼吸速率,之后测定步骤同土壤基础呼吸。碳有效性指数(Carbon availability index, CAI)为BR与SIR的比值。
1.5 土壤酶活性测定
β- 1,4-葡萄糖苷酶(β- 1,4-glucosidase, BG)、N-乙酰-β-D-葡萄糖苷酶(N-acetyl-β-D-glucosaminidase, NAG)、酸性磷酸酶(Acid phosphatase, ACP)、芳基硫酸酯酶(Aryl Sulfatase, ARS)采用对硝基苯酚(ρ-nitrophenol, PNP)比色法测定,酶活性用底物裂解后释放的对硝基苯酚含量来表征,单位为mg PNP kg-1h-1。具体方法:称取1 g鲜土,加入250 μL甲苯,4 mL修正过pH的缓冲溶液和1 mL底物,混匀后在37 ℃下培养1 h。培养结束后加入终止剂,过滤,滤液用紫外分光光度计在410 nm处比色测量对硝基苯酚含量。纤维素水解酶(Cellulase, CEL)和蛋白质水解酶(Protease, PRO)的特异性结合底物分别为羧甲基纤维素钠盐和酪蛋白,具体方法为:称取1 g鲜土,加入250 μL甲苯,5 mL缓冲溶液和5 mL底物,混匀后分别在30℃条件下培养24 h,在50℃下培养2 h。培养结束后加入终止剂过滤,滤液离心后取5 mL的上清液与7.5 mL的碱性试剂混合,用紫外分光光度计分别在710 nm和700 nm处比色测量葡萄糖和酪氨酸含量。多酚氧化酶(Polyphenol oxidase, PPO)和过氧化物酶(Peroxidase, PER)分别与10 mL 1%邻苯三酚和4 mL柠檬酸磷酸盐(过氧化物加入10 mL 1%邻苯三酚和2 mL 5%H2O2)(缓冲至pH 4.5),并在30°C下培养2 h,培养结束后加入混35 mL乙醚萃取。乙醚浸提液在分光光度计430 nm进行比色。单位mg PPG kg-1h-1[13- 15]。
1.6 土壤DNA提取和宏基因组测序
土壤微生物DNA采用DNeasy® PowerSoil® Kit试剂盒提取(Qiagen公司),每个土壤样品称取0.35 g鲜土,提取步骤按照试剂盒说明书进行。提取完成后采用nanodrop 2000测定DNA浓度,取2 μg DNA样品,使用超声波破碎仪Covaris M220将DNA打断为约300 bp的片段。采用NEXTFLEX Rapid DNA-Seq Kit构建PE文库,包括接头链接、使用磁珠筛选去除接头自连片段、利用PCR扩增进行文库模板的富集、磁珠回收PCR产物。使用Illumina NovaSeq测序平台进行桥式PCR宏基因组测序(上海美吉生物医药科技有限公司),测序深度为每样品12 GB。
宏基因组序列分析选取软件fastp[16](https://github.com/OpenGene/fastp, version 0.20.0)对序列3′端和5′端的adapter序列进行剪切,并去除剪切后长度小于50 bp、平均碱基质量值低于20以及含未知碱基的序列,保留高质量的双端序列(pair-end reads)和单端序列(single-end reads);使用拼接软件MEGAHIT[17](https://github.com/voutcn/megahit, version 1.1.2)对优化序列进行拼接组装。在拼接结果中筛选≥300 bp的序列片段(contigs)作为最终的组装结果。使用MetaGene[18](http://metagene.cb.k.u-tokyo.ac.jp/)对拼接结果中的contigs进行ORF预测。用CD-HIT[19](http://www.bioinformatics.org/cd-hit/, version 4.6.1)对所有样品预测出来的基因序列进行聚类(设置90% identity、90% coverage),每类取最长的基因作为代表序列,构建非冗余基因集。选择核酸长度大于等于100bp的基因,并将其翻译为氨基酸序列。
使用Diamond[20](http://www.diamondsearch.org/index.php, version 0.8.35)将非冗余基因集的氨基酸最长序列与NR数据库进行比对(BLASTP比对参数设置期望值e-value为1e- 5),并通过 NR库对应的分类学信息数据库获得物种注释,然后使用物种对应的基因丰度总和计算该物种的丰度,与eggNOG数据库进行比对(BLASTP比对参数设置期望值e-value为1e- 5),获得基因对应的COG(Clusters of orthologous groups of proteins)功能,然后使用COG对应的基因丰度总和计算该COG的丰度。
1.7 数据统计
本文中数据均表示为平均数±标准误(n=3)。采用R 4.0.3统计分析软件中t检验(双边检验)比较树岛和同海拔开阔区土壤理化性质、土壤微生物生物量和基因相对丰度。用Response ratio计算树岛相对于开阔区土壤酶活性、优势微生物类群相对丰度及功能基因相对丰度的响应比(α=0.05)[21]。采用Pearson相关分析计算土壤微生物优势类群相对丰度、功能基因、土壤酶活性与土壤理化性质间的相关性。柱形图采用R语言ggplot 2软件包绘制,热图采用R语言pheatmap软件包绘制。
2 结果与分析
2.1 树岛与同海拔开阔区土壤养分差异
由表1可以看出,树岛的土壤理化性质异质性相对于开阔区更强(树岛的标准误高于同海拔开阔区)。树岛含水量显著高于同海拔开阔区(P= 0.02),树岛内土壤平均含水量为50%,而同海拔开阔区处土壤平均含水量为45%。树岛内土壤全碳全氮平均含量分别为138.5 g/kg和7 g/kg,而开阔区处土壤分别为101.2 g/kg、5.3 g/kg。t检验表明,树岛土壤的平均全碳、全氮含量显著高于开阔区(P<0.05),分别比开阔区高37%和32%。土壤平均含水量与TC、TN呈正相关关系(r=0.99和0.97;P<0.01)。树岛土壤可溶性碳氮(DOC和DON)及速效养分含量(速效磷、铵态氮、硝态氮)含量与开阔区没有显著差异(P>0.05)。
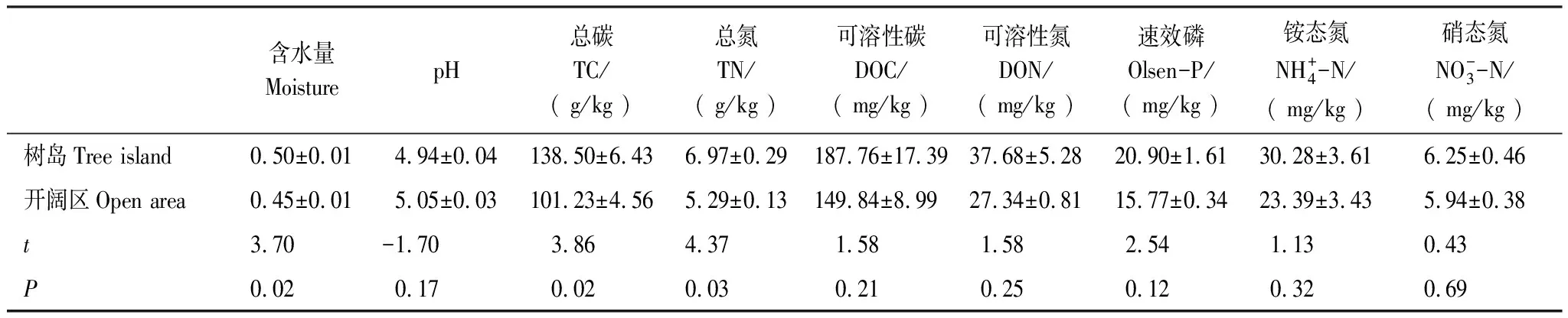
表1 树岛与同海拔开阔区土壤理化性质
2.2 树岛与同海拔开阔区土壤微生物生物量和活性差异
从图2可知,树岛土壤平均微生物生物量碳(MBC)和氮(MBN)均显著高于开阔区(P<0.05),分别高于开阔区77%和92%。树岛土壤平均基础呼吸(BR)和碳有效性指数(CAI)显著高于开阔区(P<0.05),底物诱导呼吸(SIR)没有显著差异(P>0.05)。
树岛土壤纤维素酶(CEL)平均活性显著高于开阔区(P<0.001),分别为664.03 mg Glu kg-1d-1和281.30 mg Glu kg-1d-1(表2)。开阔区土壤的多酚氧化酶(PPO)平均活性显著高于树岛(P<0.05),分别为229.00 mg PPG kg-1h-1和139.21mg PPG kg-1h-1。其他检测的6种土壤酶活性,包括β- 1,4-葡萄糖苷酶(BG)、N-乙酰-β-D-葡萄糖苷酶(NAG)、过氧化物酶(PER)、酸性磷酸酶(ACP)、芳基硫酸酯酶(ARS)、蛋白酶(PRO),在树岛及开阔区之间没有显著差异(P>0.05)。
2.3 树岛与同海拔开阔区土壤微生物结构组成差异
树岛和开阔区土壤微生物优势类群相似(图3),其中变形菌门(Proteobacteria)平均丰度最高,分别占树岛和开阔区土壤微生物群落的40.5%和45.7%,放线菌门(Actinobacteria)其次,分别占35.7%和29.5%,放线菌门在树岛土壤中平均丰度显著高于开阔区(P<0.05);酸杆菌门(Acidobacteria)分别占比10.7%和11.3%,酸杆菌门在开阔区土壤丰度显著高于树岛(P<0.05)。其他相对丰度在1%—10%之间的优势类群还包括绿弯菌门Chloroflexi、疣微菌门Verrucomicrobia、拟杆菌门Bacteroidetes、厚壁菌门Firmicutes、蓝细菌Cyanobacteria、浮霉菌门Planctomycetes、子囊菌门Ascomycota。
2.4 树岛与同海拔开阔区土壤微生物功能基因差异
根据COG分类体系,功能基因被分为4大类(level 1):新陈代谢、信息储存和加工、细胞加工和信号传递及功能未知。4个大类别又可以进一步细分为26个亚类(level 2),其中树岛土壤宏基因组亚类中与转录功能(Transcription, COG K)、防御机制(Defense mechanisms, COG V)、细胞分裂相关的功能(Cell cycle control, cell division, chromosome partitioning, COG D)基因丰度显著高于开阔区土壤(P<0.05)。开阔区土壤中无机离子运输和代谢相关的基因(Inorganic ion transport and metabolism, COG P)显著高于树岛土壤(P<0.05)。碳水化合物运输和代谢(Carbohydrate transport and metabolism, COG G)基因的总体丰度在树岛与开阔区间无显著差异,但在构成COG G亚类的不同种COG(level 3)中进一步比较发现(图4),在树岛土壤中含有更多的COG0366和COG1472基因,即淀粉酶水解基因与糖基水解酶基因(β- 1,4-葡萄糖苷酶),而开阔区中表达了更多的COG0726和COG3459,即多糖脱乙酰酶及纤维二糖磷酸化酶。
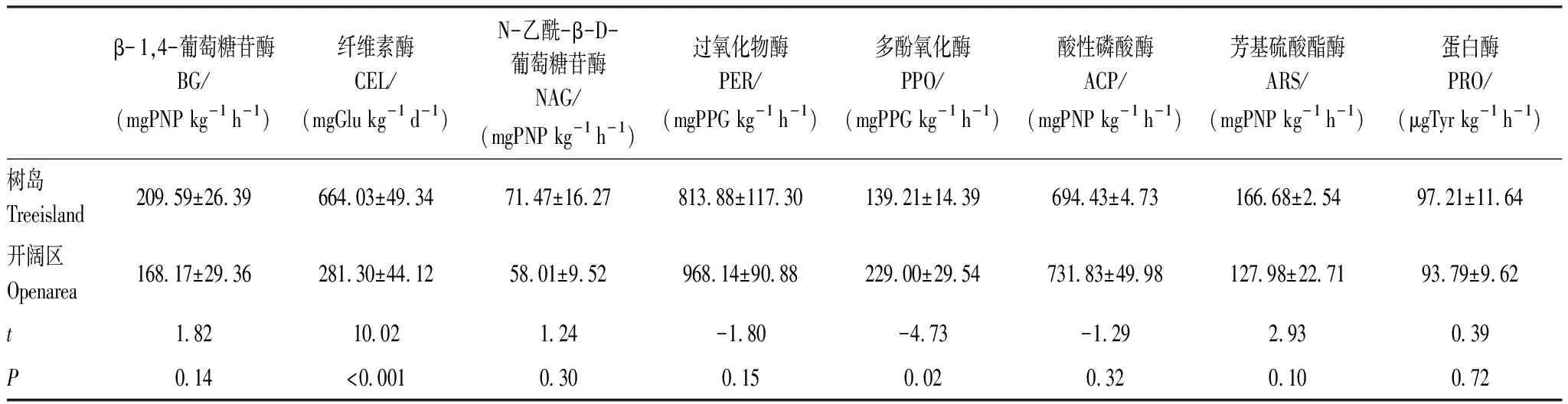
表2 树岛与同海拔开阔区土壤酶活性差异
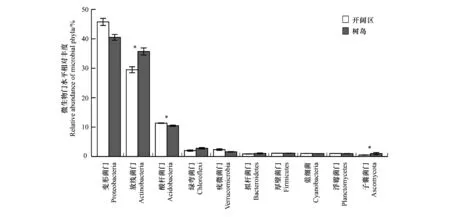
图3 树岛与同海拔开阔区土壤优势微生物类群相对丰度Fig.3 Differences in the relative abundance of selected microbial phyla between tree islands and open area at the same altitude*P<0.05,**P<0.01
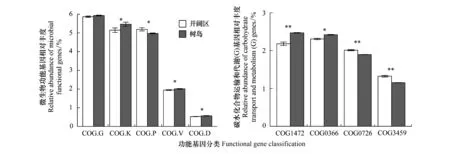
图4 树岛与同海拔开阔区土壤微生物功能基因差异 Fig.4 Differences in the abundance of soil microbial functional gene between tree islands and the open area at the same altitude*P<0.05,**P<0.01; COG.G: 碳水化合物运输和代谢Carbohydrate transport and metabolism;COG.K: 转录功能Transcription;COG P: 无机离子运输和代谢相关的基因Inorganic ion transport and metabolism;COG.V: 防御机制Defense mechanisms;COG D: 细胞分裂相关的功能Cell division ;COG1472: β-葡萄糖苷酶基因Beta-glucosidase ;COG0366: α-淀粉酶水解基因Alpha-amylase ;COG0726: 肽聚糖/木聚糖/几丁质脱乙酰酶基因Peptidoglycan/xylan/chitin deacetylase ;COG3459: 纤维二糖磷酸化酶Cellobiose phosphorylase
2.5 土壤微生物群落组成和功能与土壤理化性质相关关系
由图5可知,酸杆菌门(Acidobacteria)相对丰度、多糖脱乙酰酶基因(COG0726)和纤维二糖磷酸化酶(COG3459)基因以及与木质素降解相关的土壤酶(多酚氧化酶PPO和过氧化物酶PER)活性与土壤水分(Moisture)、全碳(TC)、全氮(TN)、可溶性碳(DOC)、速效磷(Olsen-P)、碳有效性指数(CAI)、土壤呼吸(BR)呈负相关关系(图5右上蓝色部分)。而放线菌门(Actinobacteria)相对丰度、COG1472(糖基水解酶基因)以及纤维素水解酶(CEL)活性与土壤水分、全碳、全氮、速效磷、碳有效性指数、土壤呼吸呈正相关(图5右下红色部分)。由于氨氮、硝氮与所有微生物学指标的相关关系均不显著(P>0.05),因此没有在图中展示。
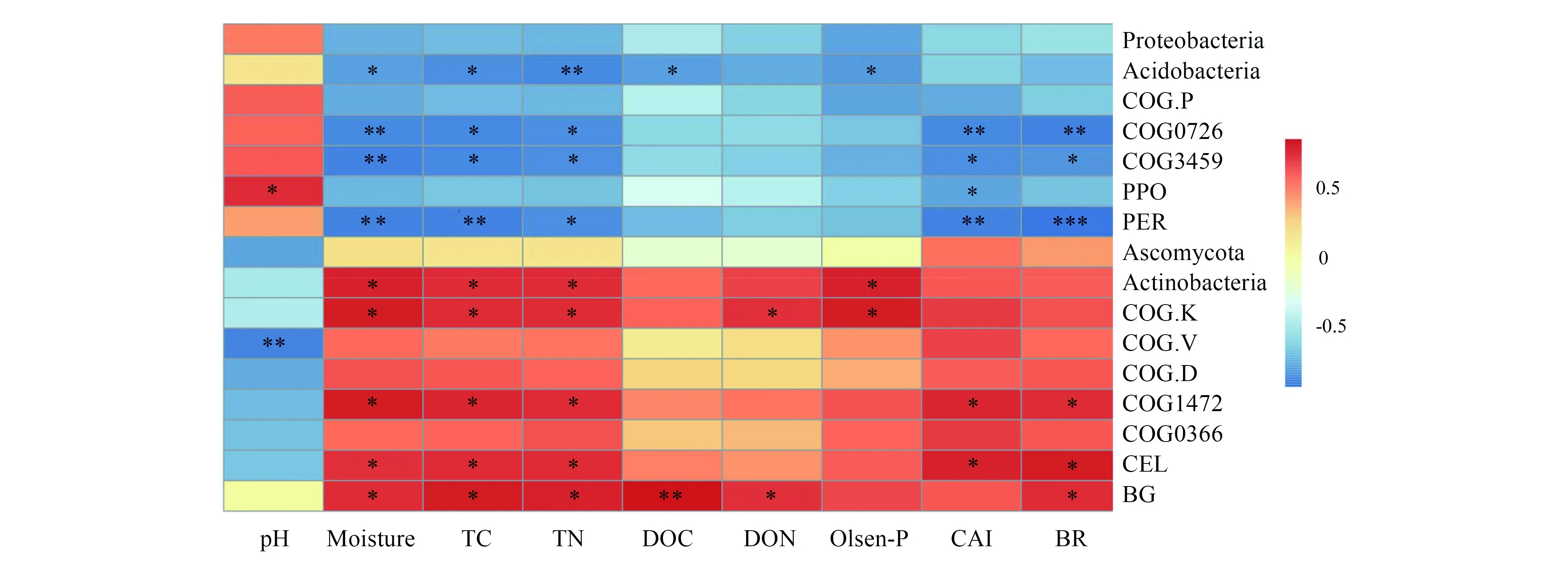
图5 微生物优势类群相对丰度、功能基因丰度和土壤酶活性与土壤理化性质的相关关系Fig.5 Correlations between soil properties and the relative abundance of specific phyla, functional genes, and soil enzymatic activities* P<0.05;** P<0.01;*** P<0.001;TC:总碳Total carbon;TN:总氮Total nitrogen;DOC:可溶性碳Dissolved organic carbon;DON:可溶性氮Dissolved organic nitrogen;CAI:碳有效性指数Carbon availability index;BR:土壤基础呼吸Basal respiration;PPO:多酚氧化酶Polyphenol oxidase;PER:过氧化物酶Peroxidase;CEL:纤维素酶Cellulase;BG:β-1,4-葡萄糖苷酶β-glycosidase
3 讨论
3.1 树岛土壤水养条件优于同海拔开阔区
本研究发现,树岛土壤的水分含量显著大于开阔区,这可能是由于在开阔区处,没有树荫遮蔽,蒸散量较大。也可能是微地形导致的,树岛位于一个地势相对低洼的区域,由于水分径流作用使树岛土壤水分富集。岳桦的径向生长受水分的影响,优越的水分条件是形成以高大乔木为主的树岛的先决条件[22]。反之,树岛内树木的定植又增加了其土壤水分保持能力。树岛土壤的全量碳、氮库均显著大于开阔区。树岛的全氮含量达到6.97 g/kg,更接近于位于海拔1800 m的郁闭岳桦林(6.5 g/kg),而同海拔开阔区全氮含量仅有5.29 g/kg, 更接近海拔2000 m林线过渡带全氮含量(4.6 g/kg)[23]。树岛内岳桦凋落物和根际分泌物向土壤中输入养分,增强了土壤肥力,而开阔区位置没有高大的乔木型岳桦,凋落物少,养分贫瘠。碳有效性指数(CAI)是用基础呼吸除底物诱导呼吸计算得到的,反映了微生物在添加底物葡萄糖前后呼吸速率的差异,从一定程度上代表了微生物的碳饥饿程度[24]。当CAI较高时,代表微生物处于碳饱和状态,指示环境的养分状态相对较好。研究中,树岛土壤的CAI相对较高,即相对于开阔区位置,其代表一种富营养生境;而开阔区土壤CAI较低,表征一种相对寡营养生境。本研究中,并未发现树岛土壤的速效养分(铵态氮、硝态氮、速效磷)含量与同海拔开阔区有显著差异(表1),这可能与采样季节有关。在夏季,前一年的凋落物已基本完成分解过程,释放到土壤中的速效养分在春季和夏季被地上植被和土壤微生物利用,而这时又没有新鲜凋落物输入,因此,速效养分在树岛内和毗邻的开阔区位置无显著差异。这一结果也提示,在未来工作中,在生长季不同时间采集土壤样品进行相关分析是十分必要的。
3.2 树岛土壤微生物的r-策略倾向和同海拔开阔区土壤微生物的K-策略倾向
r-K策略通常用来描述动植物的生活史特征,一般来说,r-策略生物在资源充足的情况下使固有生长速率最大化,而K-策略生物在资源有限的情况下能够竞争并存活下来。后来,微生物学家把r-K策略的概念借鉴到微生物生态学研究中[25],并逐渐巩固扩展[26]。r-策略微生物适合在富营养生境存活,生长速率高,生境适应广泛,对养分输入响应迅速,偏好利用相对容易降解的碳组分,例如多糖、纤维素等。由于r-策略微生物快速而频繁地调节新陈代谢并使用耗能的跨膜转运蛋白来获取营养,因此其基因组中与细胞运动(N)、防御机制(V)、转录(K)和信号转导机制(T)相关的基因都较K-策略微生物高[27]。此外,r-策略微生物参与简单碳水化合物降解基因丰度高,例如编码糖基水解酶、淀粉水解酶多糖和纤维素水解酶的基因。因此,r-策略微生物也通常具有较高的纤维素水解酶活性。K-策略微生物更适合于在寡营养环境下生长,生长缓慢,生境范围窄,偏好利用难降解碳组分[26],例如木质素,土壤酶活性也表现出降解木质素的多酚氧化酶和过氧化物酶含量更高[26]。因此,K-策略微生物也通常具有较高的木质素酶活性。一般来说,细菌中的酸杆菌门被定义为寡营养菌,即K-策略微生物;而放线菌被划分为相对富营养菌,即r-策略微生物。真菌中的子囊菌被认为具有r-策略倾向,而担子菌具有K-策略倾向。
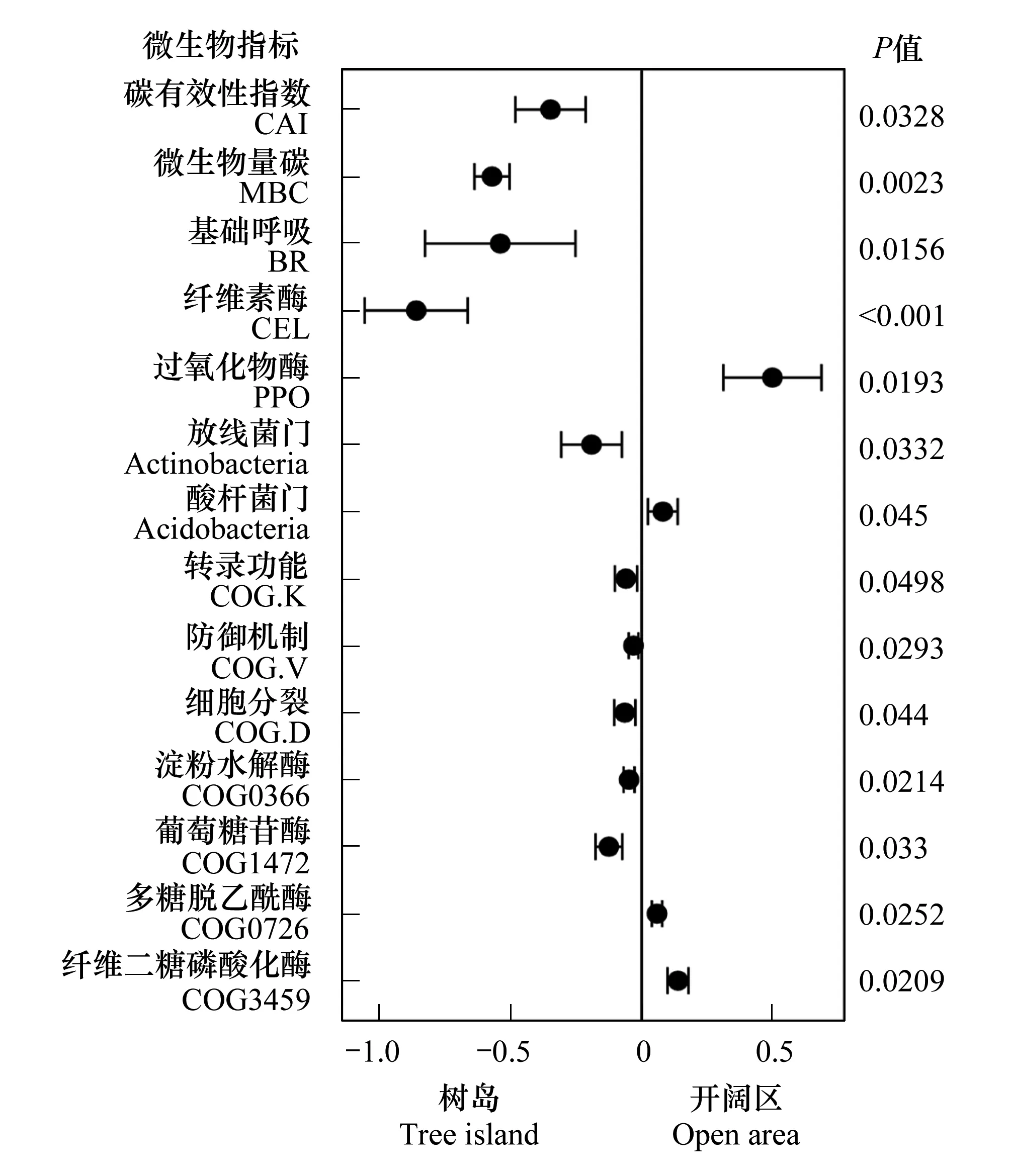
图6 树岛与同海拔开阔区土壤理化性质和微生物指标响应比 Fig.6 Response ratio of soil physicochemical properties and microbial parameters between tree islands and the open area at the same altitude
本研究发现,树岛土壤中与微生物r-策略相关的指标,包括纤维素水解酶活性、放线菌相对丰度、与细胞生长和基础代谢相关的功能基因丰度、与简单糖类化合物水解相关的基因丰度(淀粉酶水解基因与糖基水解酶基因)均高于同海拔开阔区土壤。反之,开阔区土壤中与微生物K-策略相关的指标,包括多酚氧化酶活性、酸杆菌相对丰度、复杂化合物(肽聚糖、几丁质、乙酰木聚糖和纤维素二糖)降解基因丰度较高(图6)。以上证据表明,由于树岛土壤养分更丰富,碳有效性高,土壤微生物群落倾向于采取r-策略;而开阔区土壤养分贫瘠,碳有效性低,土壤微生物群落倾向于采取K-策略。土壤酶活性的差异还可能与凋落物质量有关。在开阔区域,凋落物主要来自灌木型岳桦和其他小型灌木,如牛皮杜鹃、笃斯越桔;而在树岛内,凋落物除了来自灌木,还主要来自岳桦落叶。阔叶凋落物纤维素含量比灌木凋落物纤维素含量高[28],因此诱导了更高的纤维素酶活性。相关分析也证明,过氧化物酶、乙酰木聚糖降解、纤维二糖磷酸化酶基因等K-策略指标与指示土壤养分状态的指标呈负相关关系,包括碳有效性指数、土壤全量碳氮等;而r-策略指标则与土壤养分呈正相关关系。在之前的研究中,也有学者提出,碳有效性指数、呼吸和有效氮等土壤养分指标可以作为指示微生物r-K策略的环境因子[29]。土壤微生物这种养分利用策略反过来也会调控土壤养分周转和地上植被生长,形成正反馈效应,从而进一步促进树岛扩大或者向海拔更高处上移。
4 结论
本文研究了长白山岳桦林线过渡带树岛内外土壤微生物群落结构和潜在生态功能的差异,结果表明,树岛土壤的养分库存更高,碳有效性大,微生物生物量高,土壤微生物倾向于采取r-策略,包括较高的纤维素酶活性、富营养微生物相对丰度、与细胞生长和基础代谢相关的功能基因和分解简单碳水化合物的功能基因丰度。反之,开阔区土壤养分库和碳有效性较低,微生物生物量低,土壤微生物倾向于采取K-策略,包括较高的多酚氧化酶活性、寡营养微生物相对丰度和降解复杂碳水化合物的功能基因丰度,和较低的维持细胞生长和基础代谢功能基因丰度。土壤养分增加和微生物群落采取r-策略也可能是导致树岛形成和规模扩张的潜在驱动机制,研究结果为理解林线形成机制和预测未来气候变化条件下林线动态提供了科学依据。但本研究仅重点关注土壤养分和微生物在树岛形成过程中的作用,实际上,树岛的形成可能还与物理环境因素(例如温度、降水、风、雪等)[30]和植物生理因素(例如生长抑制和碳水化合物不足[31- 32]有关),后续还需加强树岛形成和林线动态的综合驱动机制分析。
猜你喜欢
散文诗(2021年22期)2022-01-12 06:14:04
中成药(2021年5期)2021-07-21 08:38:32
落叶果树(2021年6期)2021-02-12 01:28:54
文苑(2020年12期)2020-04-13 00:54:10
环球时报(2019-05-23)2019-05-23 06:17:36
上海包装(2019年2期)2019-05-20 09:10:52
海峡姐妹(2019年1期)2019-03-23 02:42:46
少儿美术(快乐历史地理)(2018年7期)2018-11-16 05:31:12
中成药(2018年2期)2018-05-09 07:19:49
微生物与感染(2015年1期)2015-02-28 17:42:37
