
β-烟酰胺单核苷酸对衰老小鼠的抗氧化作用研究
2022-01-14 13:16:50王思蓉王言之李世芬王光路张静姝
甘肃科技 2021年20期
王思蓉,王言之,李世芬,3,王光路,张静姝,3△
(1.南京医科大学公共卫生学院,江苏 南京,211166;2.南京医科大学第一临床医学院,江苏 南京,211166;3.江苏省医药农药兽药安全性评价与研究中心,江苏南京,211166;4.石家庄以岭药业股份有限公司,河北石家庄,050035)
β-烟酰胺单核苷酸(NMN),是哺乳动物体内一种关键性辅酶烟酰胺腺嘌呤二核苷酸(NAD+)的前体物质,其主要功能由NAD+体现[1],参与许多关键过程,包括能量代谢,基因表达和DNA 修复。β-烟酰胺单核苷酸是人体内的固有物质,同时也可从如牛油果、西兰花等[2]蔬菜水果中获取,检测发现其含量在毛豆、牛油果中尤多[3]。David Sinclair 于2019年发表的《lifespan》书中介绍了关于β-烟酰胺单核苷酸的研究,引发免疫学界、营养学界的广泛关注,但目前对β-烟酰胺单核苷酸的作用及相关机制的研究仍然存在大量空白。根据现有报告,β-烟酰胺单核苷酸对退行性疾病(如阿兹海默症[4]、帕金森疾病[5])具有一定的治疗意义。补充一定剂量的NMN能增加NAD+的含量,可促进神经血管再生,起到线粒体保护、抗炎抗凋亡的作用[6]。对于某些罕见疾病如Leigh 综合征,给予NMN 能延长患该病小鼠寿命并缓解有关症状[7]。为探究β-烟酰胺单核苷酸更多的作用价值,现将β-烟酰胺单核苷酸作为受试物,分高、中、低3 个剂量组对小鼠灌胃并测量MDA、SOD、GSH 的指标含量,拟对β-烟酰胺单核苷酸对衰老小鼠的抗氧化作用进行评价,为相应的临床应用和产品研发提供实验数据支持。
1 材料和方法
1.1 配方筛选
抗氧化系列项目配方筛选,配方为单一NMN成分,设定人体推荐摄入量为250、125、60mg/人/日(60kg),按这3 个配方组合设计动物实验,NMN 由石家庄以岭药业股份有限公司提供。
1.2 实验动物及饲养环境
选用上海西普尔-必凯实验动物有限公司繁殖的SPF 级ICR 健康8 月龄雄性小鼠50 只,饲养于南京医科大学卫生分析检测中心屏障系统,温度20~25℃,相对湿度40%~70%,生产许可证号SCXK(苏)2018-0006,合格证编号:20180006013099。
1.3 试验方法
以上3 种配方,均按人体推荐剂量的10 倍作为动物实验灌胃剂量,将200mg/人/日VC 剂量组作为阳性对照组,各剂量组用无菌水配制;同时设溶剂对照组,给与无菌水。小鼠适应环境至少3d,按血清中MDA 水平分为5 组,每组10 只。各剂量组小鼠给予不同受试样品按0.1mL/10g 体重进行灌胃,1次/d,连续30d。实验结束时取血测血清中脂质氧化产物含量,取肝测肝组织中蛋白质羰基含量、还原性谷胱甘肽含量及抗氧化酶活力。
2 结果
2.1 对小鼠一般情况的影响
小鼠灌胃前后体重见表1,数据处理知,各剂量组与溶剂对照组相比,各周体重均无统计学差异(P>0.05),反映NMN 对小鼠体重无明显影响。
表1 对小鼠各周体重的影响()
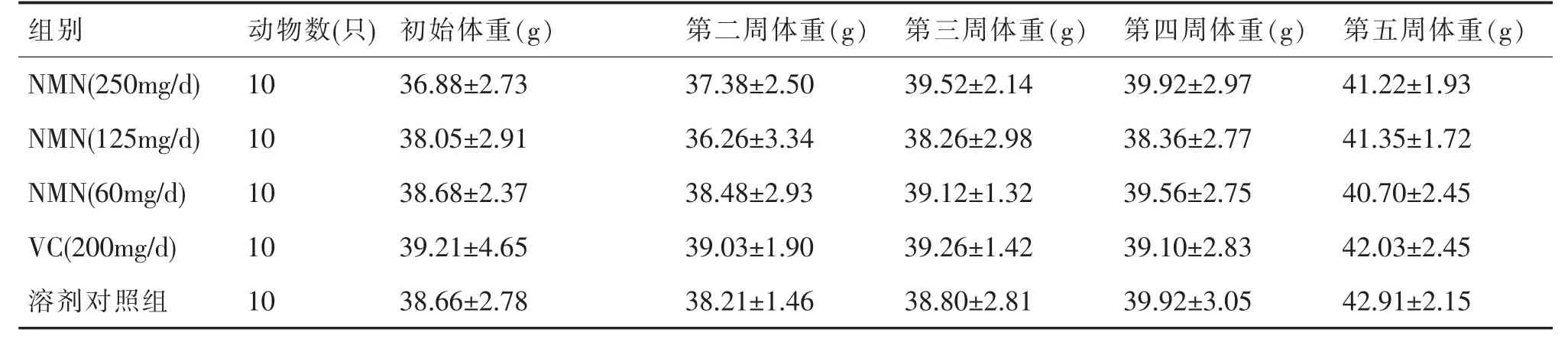
表1 对小鼠各周体重的影响()
2.2 小鼠血清中MDA 的影响
由表2 可见,除溶剂对照组外,各观察组实验后MDA 相对实验前均存在不同程度的降低,以NMN(125mg/d)、VC(200mg/d)组最为显著,差异有统计学意义(P<0.05)。
表2 小鼠血清中MDA 的含量()
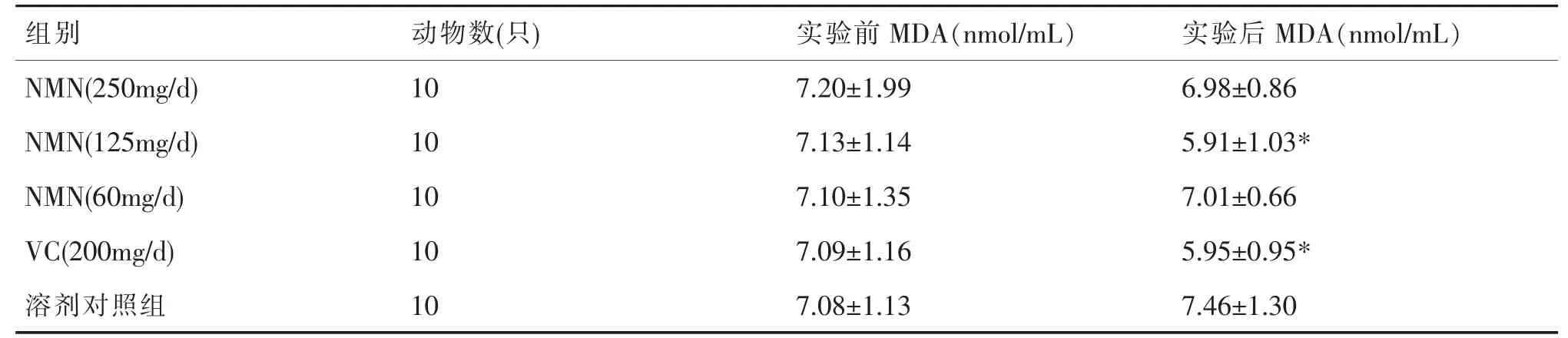
表2 小鼠血清中MDA 的含量()
注:“*”表示与溶剂对照组比较,P<0.05。
2.3 小鼠肝组织中蛋白质羰基含量
由表3 可见,和溶剂对照组相比,各剂量组小鼠的蛋白质羰基含量均有所减少,其中VC(200mg/d)组的差异有统计学意义(P<0.05),而NMN(125mg/d)组虽然相对阴性对照组蛋白质羰基含量平均值下降了26.09%,但差异无统计学意义,或与实验动物数量偏少存在一定联系。
表3 小鼠肝组织中蛋白质羰基的含量()
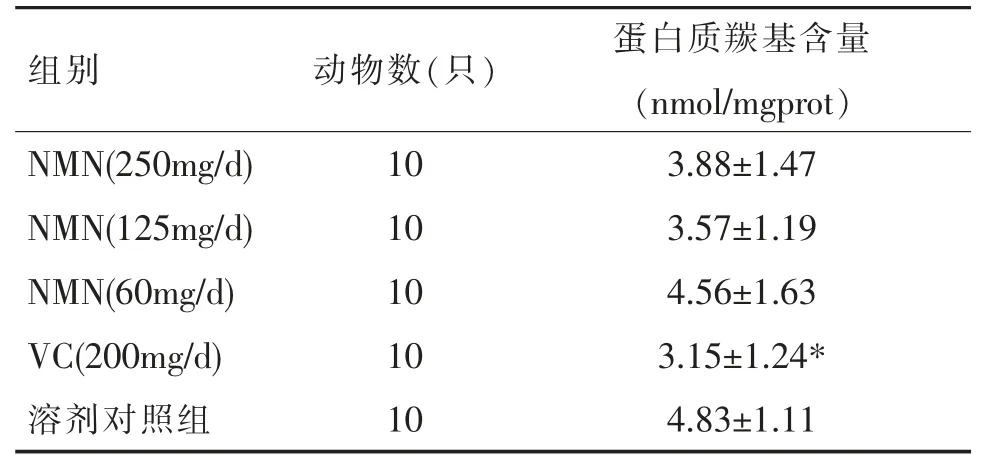
表3 小鼠肝组织中蛋白质羰基的含量()
注:“*”表示与溶剂对照组比较,P<0.05。
2.4 小鼠肝组织中SOD 及GSH-PX 活力
由表4 可见,各观察组小鼠肝组织中SOD 均高于阴性对照组,以NMN(250mg/d)组效果最明显,平均相对增长了72.9(U/mgprot),NMN(60mg/d)组次之,平均相对增长了49.9(U/mgprot)。NMN(250、125、60mg/d)及VC(200mg/d)组差异均有统计学意义(P<0.05)。
表4 小鼠肝组织中SOD 及GSH-PX 活力()
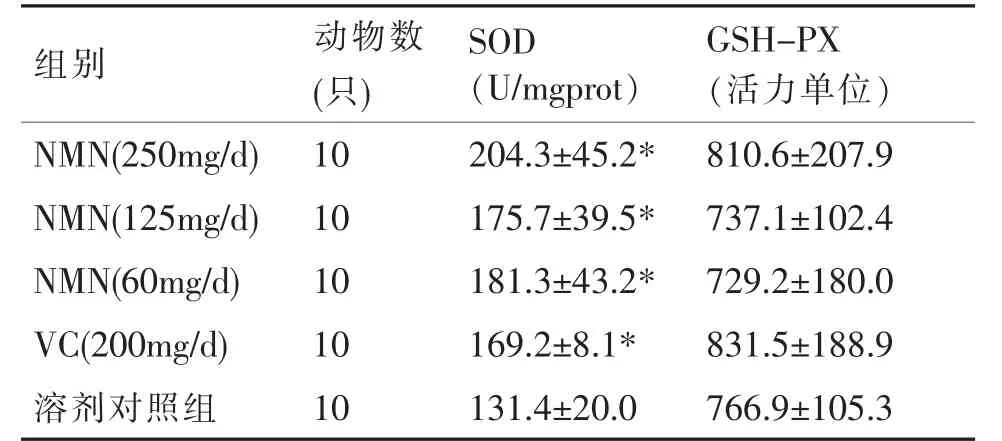
表4 小鼠肝组织中SOD 及GSH-PX 活力()
注:“*”表示与溶剂对照组比较,P<0.05。
2.5 小鼠肝组织中GSH 含量
由表5 可见,NMN(125mg/d)组小鼠肝组织中GSH 高于溶剂对照组,差异有统计学意义(P<0.05)。与VC(200mg/d)组比较,NMN(125mg/d)组小鼠肝组织中GSH 含量显著升高,差异有统计学意义(P<0.05)。
表5 小鼠肝组织中GSH 含量()
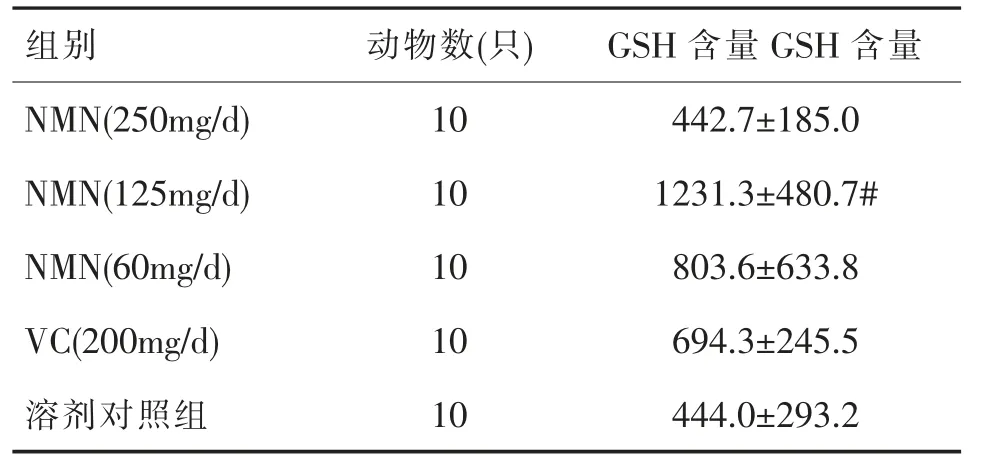
表5 小鼠肝组织中GSH 含量()
注:“#”表示与VC(200mg/d)组比较,P<0.05。
3 结论
参照 《保健食品检验与评价技术规范》(2003版)之抗氧化功能试验的结果判定,在本实验条件下,NMN 对衰老小鼠具有抗氧化作用,且NMN 在125mg/d 的剂量下对衰老小鼠的抗氧化作用明显优于250mg/d 和60mg/d 剂量组。
4 讨论
β-烟酰胺单核苷酸作为辅酶NAD+的前身,其发挥相应的功能作用由NAD+完成。因NAD+分子很难直接进入细胞,故补充NAD+是以服用NAD+前体形式吸收入血液,才能让其在细胞内得到利用和转化。Zhang Hongbo 等[8]已证实补充一定剂量的NAD+能改善小鼠的线粒体功能,延长寿命。但NAD+的含量水平以及其氧化还原状态却随年龄的增大而呈下降趋势[9],其导致年龄相关的NAD+减少的机制还未被阐明[10]。β-烟酰胺单核苷酸作为NAD+前体在补充NAD+的含量以及增强其相应功能具有重要意义,如口服β-烟酰胺单核苷酸能有效加快合成组织中的NAD+,并缓解小鼠年龄相关的生理衰退[11]。
近年来,针对抗氧化剂的研究方兴未艾,而新型抗氧化剂在疾病防控和治疗方面的作用受到越来越多的关注。自由基、ROS 及其衍生的氧化应激物对可对机体健康造成危害,如异常衰老及其衰老相关疾病[12]、心血管系统疾病、代谢综合性疾病等[13]。利用β-烟酰胺单核苷酸的抗氧化作用,清除多余生成的自由基、ROS,降低其对机体组织的损害,已然成为延缓衰老、治疗疾病的新的靶向。补充β-烟酰胺单核苷酸从而增加NAD+含量,能有效抑制活性氧的生成、减少线粒体断裂,为急性脑损伤和神经退行性疾病提供治疗思路[14]。Natalie 等[15]指出β-烟酰胺单核苷酸能够有效逆转老龄小鼠血管功能和氧化应激,是应对老年心血管疾病的有效干预措施;Wang Tao 等[16]报道NMN 转移酶3 通过调节NAD+的水平对兔骨髓质充质干细胞具有抗氧化应激的效果,但其抗氧化的相关机制还尚未明确。另外,活性氧是恶性肿瘤的诱因[17],可引发肿瘤细胞的增殖、提高其耐受性,基于此,深化对β-烟酰胺单核苷酸抗氧化机制的相关研究可以为抑制恶性肿瘤的产生及发展开拓领域[18]。不论作为NAD+前体还是就其本身而言,β-烟酰胺单核苷酸的潜藏价值都值得人们的深入研究和探讨评估。
猜你喜欢
特产研究(2024年1期)2024-03-12 05:40:56
世界科学技术-中医药现代化(2022年3期)2022-08-22 00:33:26
肝博士(2022年3期)2022-06-30 02:48:28
云南化工(2021年11期)2022-01-12 06:06:18
Journal of Sport and Health Science(2019年6期)2019-11-26 07:30:53
分析测试学报(2015年9期)2015-12-17 16:44:27
西华师范大学学报(自然科学版)(2015年3期)2015-02-27 15:31:18
药学与临床研究(2014年3期)2014-03-06 13:35:35
癌变·畸变·突变(2014年2期)2014-03-01 04:39:42
无机化学学报(2014年6期)2014-02-28 17:32:05
