
金钱松林挥发物季节性变化对空气负离子及微生物的影响
2022-01-14 13:28韦赛君吕嘉欣张汝民
江西农业大学学报 2021年6期
韦赛君,张 静,王 翔,吕嘉欣,王 彬*,高 岩,张汝民
(1.浙江农林大学 亚热带森林培育国家重点实验室,浙江 临安,311300;2.浙江省森林资源监测中心,浙江 杭州 310020)
【研究意义】植物挥发性有机物(volatile organic compounds,VOCs)是一类小分子化合物,包括萜烯类化合物、脂肪酸衍生物、苯丙类、苯类和其他氨基酸衍生物四类等[1],在植物的化学通讯、防御反应、生长繁殖、净化空气以及改善人体健康等方面起着重要作用[2-3]。【前人研究进展】针叶树种主要释放萜烯类化合物,油松(Pinus tabuliformis)释放VOCs中萜烯类化合物占88.7%[4]。这类物质在植物体内通过次生代谢途径合成后,储存于特殊的植物贮藏结构(如树脂管、腺体)[5],小部分释放到环境中,但释放量会受到温度和光强的影响[6-7]。植物VOCs可作为化学抗剂对抗叶表面或空气中的微生物,如萜类化合物(E)-橙花醇是茶树对抗病原菌的挥发性信号[8]、薄荷精油能显著抑制金黄色葡萄球菌(Staphylococcus aureus)生物膜的形成[9]、己烯醛能使尖孢炭疽病菌(Colletotrichum acutatum)失活[10];研究证明针叶树挥发油具有抑菌和抗真菌作用,使针叶林内的空气微生物数量显著低于空旷地[11]。植物在生长过程中释放的VOCs能促进空气离子化,增加森林中空气负离子(NAI)浓度[12],且植物结构越复杂,NAI浓度越高[13]。同时NAI能利用其单极性离子的特征和带正电的颗粒物(PM)相互附着沉降,能有效降低颗粒物浓度,改善空气质量[14],是衡量空气清洁度的指标之一。【本研究切入点】金钱松(Pseudolarix amabilis)是松科金钱松属的落叶乔木,目前对其研究大多集中在群落分布[15]、生物学特性[16]、遗传多样性[17]、病理学[18]和化学成分[19]等领域,关于金钱松释放VOCs对空气微生物抑制作用方面的研究还未见报道。【拟解决的关键问题】本研究以金钱松为对象,采用动态顶空采集法和热脱附-气相色谱-质谱(TDS-GC-MS)联用技术,分析了金钱松释放VOCs和春夏秋季林内空气VOCs组分和含量,并检测林内空气负离子浓度和空气微生物数量,揭示金钱松林释放VOCs的季节变化和抑菌效果,为金钱松生态学价值的利用和园林配置提供理论依据。
1 研究区自然条件与植被概况
研究区位于浙江省临安区天目山国家级自然保护区,其地理位置为30°18′30″~30°24′55″N,119°24′11″~119°28′21″E,属于中亚热带湿润型季风气候,年平均气温14 ℃,年降水量达1 390~1 870 mm,太阳辐射4 460~3 270 MJ/m2,相对湿度76%~81%。树种资源丰富,古树名木众多,植被从山麓到山顶垂直分布为常绿阔叶林、常绿阔叶混交林和落叶阔叶林,植被覆盖率达95%。
金钱松人工林面积约为900 m2,从中选择3个点进行实验,平均树高约为9 m,平均胸径8 cm,树龄为15 年左右,郁闭度0.6~0.8;林下草本层高5~80 cm,盖度45%~60%,主要植被有吉祥草(Reineckia carnea)、络石(Trachelospermum jasminoides)和白茅(Imperata cylindrica)等。
2 材料与方法
2.1 供试树种
以自然状态下生长健康、长势良好的金钱松为研究对象,随机选取5棵生长情况基本相似的金钱松,每棵作为一个独立重复实验。对照为空旷地(待建停车场,地面植被已整修干净,呈裸露土地状,约0.2 hm2),无树木影响,选取5个点进行采样(图1)。
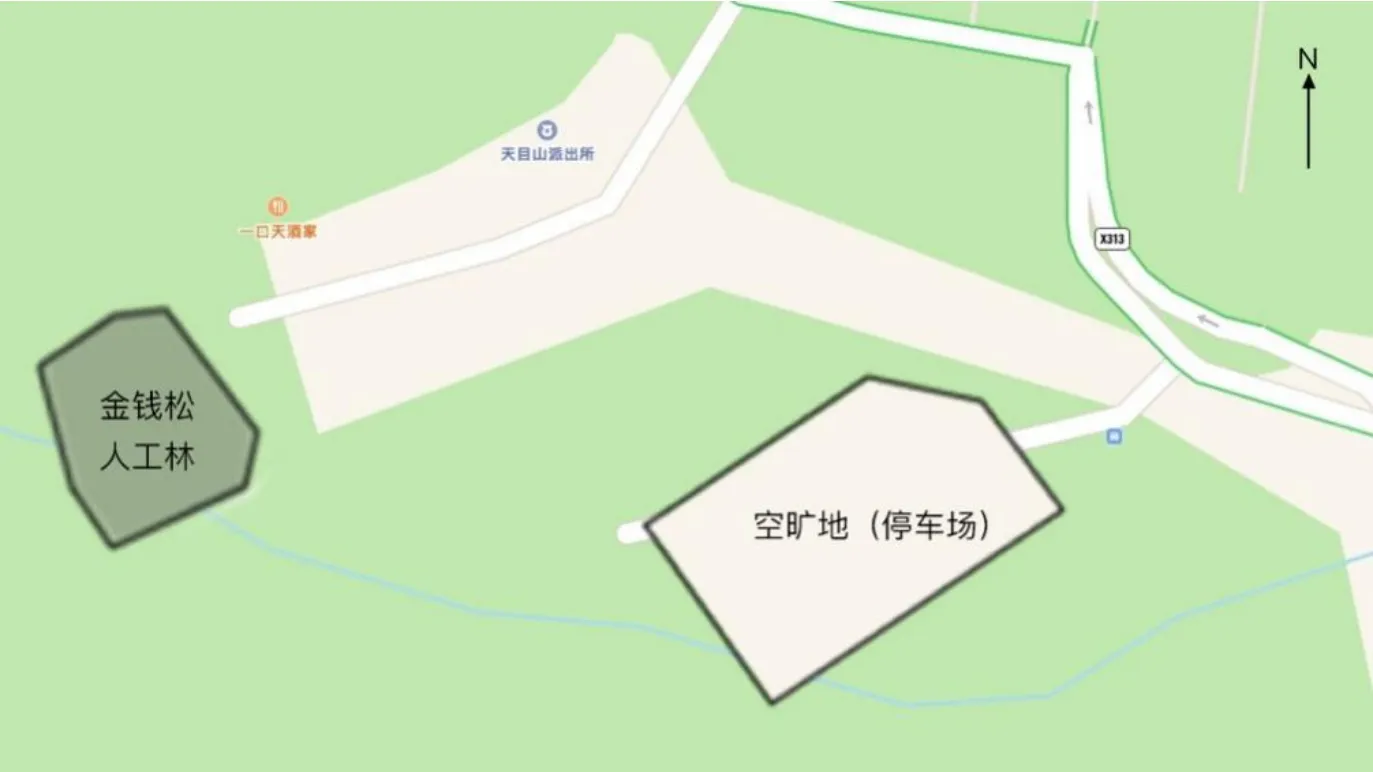
图1 采样点Fig.1 Schematic diagram of sampling points
2.2 方法
2.2.1 VOCs采集及分析方法 单株VOCs采集:2019年5月,利用QC-2型大气采样仪,采用动态顶空套袋法对金钱松向阳面茂盛枝叶VOCs 进行采集,采样气体循环流量为100 mL/min,采气时间60 min。采集后剪下枝叶,在烘箱80 ℃条件下干燥6 h 后测定干质量。林内VOCs 采集:2019 年5 月,7 月,10 月,利用QC-2型大气采样仪连接洁净硅胶管和装有吸附剂Tenax2GR(200 mg,60~80目)的采样管连接玻璃漏斗对金钱松林内空气VOCs 进行开放式采集,采集高度为1.5 m,采样气体循环流量为100 mL/min,采气时间60 min。VOCs 成分分析采用热脱附-气相色谱-质谱联用仪(TDS-GC-MS)法。仪器正常工作条件参数设置参照Gao等[11]的方法。
2.2.2 空气负离子测定 利用森林康养监测系统(YZQ-308A)测定空旷地(对照)和林内空气负离子,测定高度为1.2 m左右,在林内选取5个点,每个点在相互垂直的4个方向分别测定[20],待仪器稳定后每个方向连续读取5 个有效值,仪器每2 min 读取一次,测定时间为3 d,每天观测时间为07:00—19:00,每3 h 记录一次。
2.2.3 培养基和单体溶液的配制 按照沈萍[21]的方法配制细菌培养基(牛肉膏蛋白胨培养基)、真菌培养基(马丁式培养基)和放线菌培养基(淀粉琼脂培养基)。根据金钱松叶片挥发物的种类,从中选择释放含量较高的6 种单体,萜烯类和醛类各3 种(α-蒎烯、柠檬烯、石竹烯、辛醛、壬醛和癸醛),配制浓度参考Gao 等[11]。在无菌条件下制备单体(sigma 公司),在每个标准单体中加入2 滴吐温-80,用无菌纯水稀释100倍、200倍和1 000倍浓度,与培养基混合,单体∶培养基=1∶9。
2.2.4 微生物的采集 微生物的采集分别在2019 年5 月、7 月和10 月,选择晴朗无风或微风的天气,10:00—13:00,采用自然沉降法[22],在金钱松林内和空旷地各设5 个采样点,将带有不同培养基的培养皿分别置于离地面1.2 m 高和距离树干1.0 m 的平板支架上,打开培养皿盖,暴露在空气中10 min,采集空气中自然漂浮的微生物(细菌、真菌和放线菌),采集完毕用无菌纸包裹每个培养皿。单体实验:将带有不同浓度单体培养基的培养皿,于空旷地各设5 个采样点,采集方法同上。接菌的培养皿在恒温培养箱30 ℃条件下培养,细菌培养48 h 后计数菌落、真菌和放线菌培养96 h 后进行菌落统计。每个点的微生物连续采集3 d。
2.2.5 数据处理 通过分析获得VOCs 的总离子流量色谱图,对比NIST 2008 谱库,根据已报道的植物VOCs 保留时间,并结合手工检索对VOCs 各组分进行定性,然后选取特征离子进行进一步定量分析。金钱松单株释放VOCs 的含量采用单位采样时间内每克植物枝叶释放出的VOCs 特征离子峰峰面积进行定量。

其中N为培养皿中菌落平均数(个);A为培养皿的面积(cm2);T为打开培养皿皿盖的时间(min)。

所有数据均为5次重复的平均值±标准差。利用Origin9.0(Origin Lab公司,美国)数据处理系统进行数据统计、制图和方差分析。采用One-Way ANOVA 对金钱松林和空旷地分别进行检验,并进行Tukey多重比较(P<0.05)。
3 结果与分析
3.1 金钱松叶片释放VOCs成分分析
将金钱松活体植株的VOCs 进行TDS-GCMS 分析,得到总离子流图(图2)。从中鉴定出酯类、萜烯类、醛类、醇类、苯类和酚类等共29种化合物(表1),酯类化合物共7种,占总挥发物含量的36.2%;萜烯类11种,占29.5%;醛类4种,占7.6%;苯类2 种,占8.3%;醇类2 种,占5.5%。主要物质有乙酸叶醇酯(25.2%)、α-蒎烯(11.8%)、3-己烯醇(5.1%)、水杨酸甲酯(4.6%)、石竹烯(4.1%)、瑟林烯(3.9%)、壬醛(3.2%)和柠檬烯(2.6%)等。

表1 金钱松叶片释放VOCs的主要成分Tab.1 The main components of VOCs from the leaf of P.amabilis
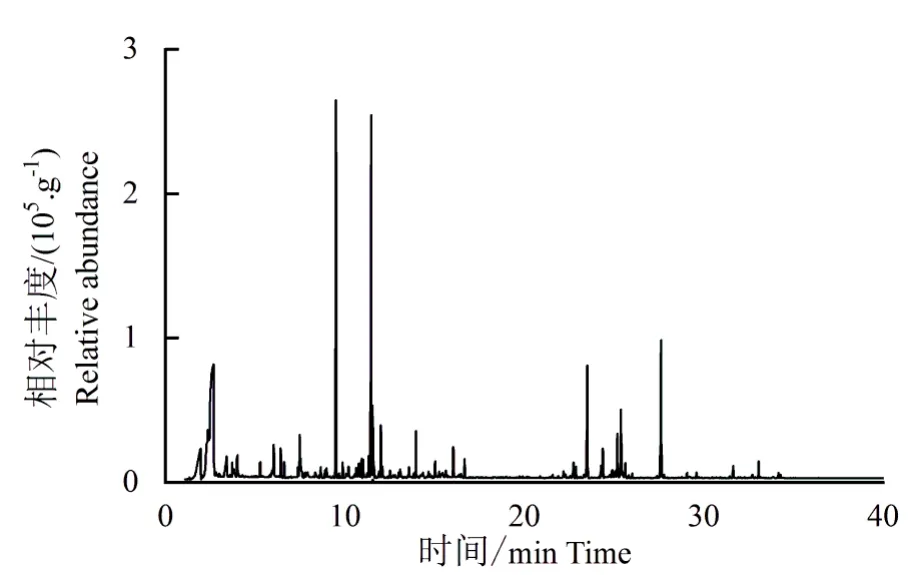
图2 金钱松叶片释放VOCs总离子流分析Fig.2 TIC of VOCs in leaves of P.amabilis
3.2 不同季节金钱松林空气VOCs分析
春、夏、秋季金钱松林共检测到8类27种化合物,主要有醛类、萜烯类、烷烃类、苯类和酯类等(表2)。金钱松林内空气VOCs与空旷地相比增加了α-蒎烯、柠檬烯、雪松烯和α-柏木烯等为主的萜烯类物质和辛醛、壬醛和癸醛为主的醛类物质。不同季节金钱松林内空气VOCs组分和含量存在差异(图3),夏季检测到27种化合物,主要有壬醛(5.5%)、癸醛(5.2%)、柠檬烯(4.9%)、己醛(3.5%)、二甲基辛醇(3.5%)和雪松烯(3.3%)等;春季检测到23 种,主要为糠醛(8.8%)、丁酸丁酯(6.1%)和柠檬烯(4.4%)等;秋季检测到18种,主要为柠檬烯(8.0%)、癸醛(8.0%)、壬醛(7.7%)和辛醛(7.3%)等。总VOCs含量在夏季达到峰值,约为春季的5倍,是秋季的150倍左右。
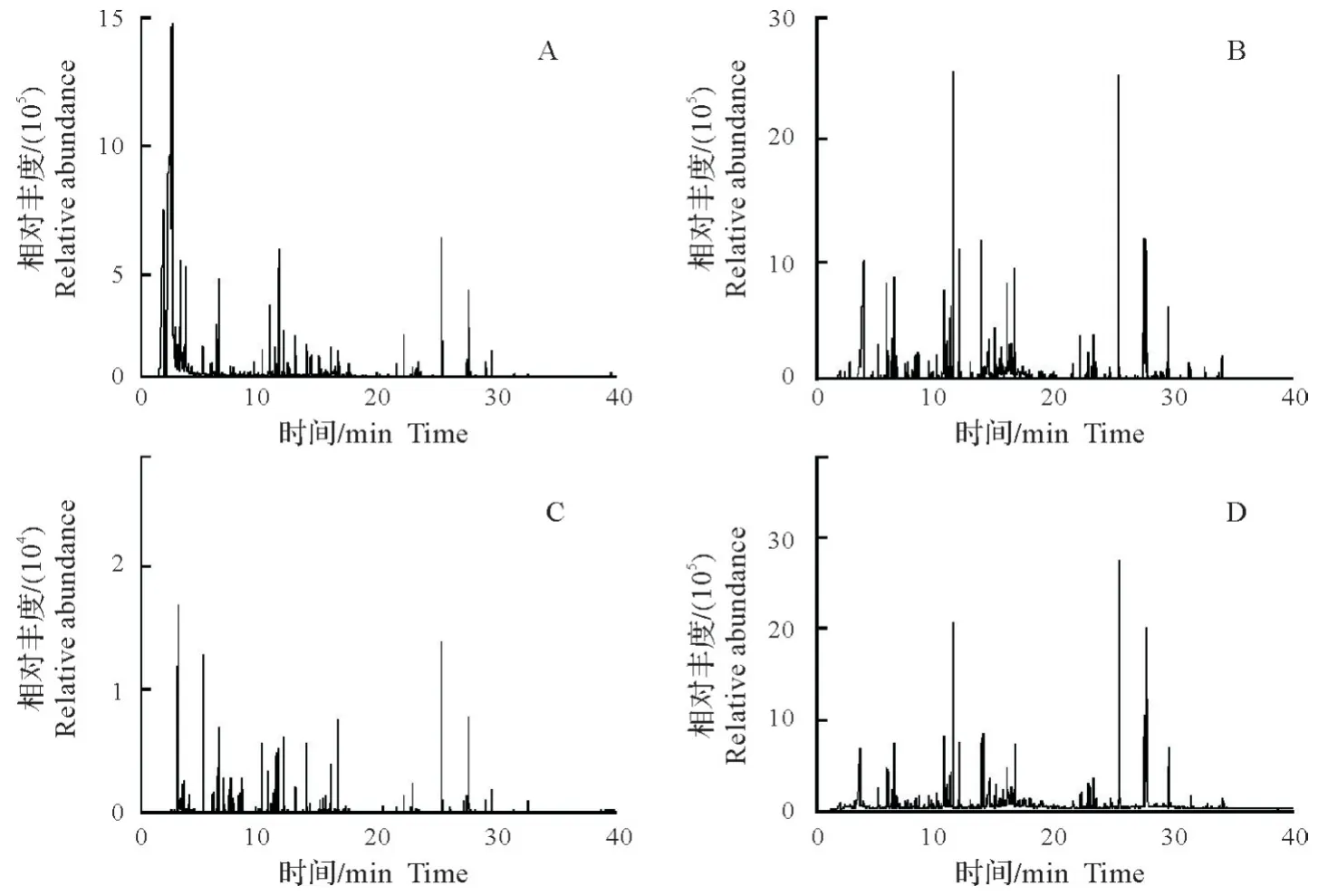
图3 不同季节金钱松林空气VOCs成分总离子流分析Fig.3 TIC of VOCs in air of P.amabilis forest in different seasons
3.3 不同季节空气负离子变化
金钱松林内空气负离子含量有明显的季节变化(图4)。林内空气负离子含量夏季最高,春季最低,夏季比春季增加了35.9%(P<0.05),比秋季增加了23.5%(P<0.05),春季与秋季之间差异不显著。金钱松林内空气负离子含量显著高于空旷地,林内含量在春、夏、秋季分别比空旷地增加了39.1%、72.3%和61.6%(P<0.05)。

图4 不同季节空气负离子含量变化Fig.4 Seasonal variation of negative ion content
3.4 金钱松林内空气微生物季节变化
随着季节的变化,金钱松林内微生物数量有一定的波动(图5)。春季林内细菌数量最少,秋季最多,春季比秋季降低了31.2%(P<0.05);真菌和放线菌数量均在夏季最少,分别比秋季降低了34.0%和20.7%(P<0.05),放线菌夏季数量比春季降低了15.9%(P<0.05)。空旷地细菌数量在春、夏、秋季有一定的变化,春季最少,与夏季相比降低了16.2%(P<0.05);真菌和放线菌数量在春、夏、秋季无显著差异。金钱松林内微生物数量与空旷地相比明显下降,夏季的下降幅度最大,细菌、真菌和放线菌分别降低了45.7%、48.3%和36.9%(P<0.05);夏季细菌降幅比春季多5.9%,比秋季多23.5%;真菌降幅比春季多19.5%,比秋季多29.4%;放线菌降幅比春季多23.4%,比秋季多19.7%。

图5 不同季节金钱松林空气微生物的数量变化Fig.5 The quantity change of air microorganism in P.amabilis forest in different seasons
3.5 单体对空气微生物的影响
单体对空气微生物有一定的抑制作用(图6),1%时柠檬烯和石竹烯能显著抑制细菌生长,抑制率分别为38.1%和23.0%(P<0.05);0.1%时3种萜烯均能促进真菌的生长,随着体积分数升高,呈现出抑制作用;1%时α-蒎烯和柠檬烯对放线菌抑制率分别为41.7%和30.6%(P<0.05),0.1%时石竹烯对放线菌的抑制率为23.3%(P<0.05)。0.1%时壬醛对细菌的抑制率为14.3%(P<0.05),0.5%时辛醛和癸醛对细菌的抑制率分别为24.8%和42.6%(P<0.05);0.5%时辛醛和壬醛对真菌的抑制率分别为17.9%和21.1%(P<0.05),1%时癸醛对真菌抑制率为28.5%(P<0.05);0.1%时辛醛对放线菌抑制率为24.4%(P<0.05),1%时壬醛和癸醛对放线菌的抑制率分别为42.2%和51.1%(P<0.05)。

图6 单体对空气微生物的抑制作用Fig.6 Inhibition of airborne microorganisms by monomer compounds
6种单体中,醛类对微生物的抑制作用比萜烯更为明显,柠檬烯和石竹烯能抑制细菌生长;低浓度下萜烯类对真菌有促进作用;石竹烯对放线菌有明显的抑制作用;醛类对3种菌都有显著抑制作用。
4 讨论
植物挥发物是一类小分子次生代谢产物,可以作为生长发育的调控信号,也是抵御胁迫、保护自身的重要物质[2],是植物进化过程中对自然环境适应的结果[23]。不同树种释放的VOCs 成分和含量均有差异,毛白杨(Populustomentosa)、栓皮栎(Quercus variabilis)和色木槭(Acer mono)等阔叶树主要释放异戊二烯[24],而侧柏(Platycladus orientalis)和油松等针叶树主要释放单萜类化合物。Aydin 等[25]研究以7 种松属(Pinus)、4 种刺柏属(Juniperus)和4 种其他树种为例的针叶树种,发现其平均单萜类释放率约为69%。已有研究发现金钱松叶片挥发油成分中主要为萜烯类物质,其中α-蒎烯和石竹烯含量最多[26]。植物VOCs的合成和释放受到温度和光照的影响,Jardine等[7]研究发现植物叶片中单萜排放来源于光合作用,VOCs 在黑暗中几乎停止排放,在中午排放速率达到峰值[6]。本研究中金钱松叶片释放的VOCs 以酯类(乙酸叶醇酯)和萜烯类物质为主,与前人研究结果有所差异,可能是采集方法不同所致。金钱松林内VOCs与空旷地相比增加萜烯类和醛类物质,这些物质主要由金钱松枝叶释放;林内夏季VOCs含量显著高于春秋两季,可能是夏季光照强,金钱松处于生长旺季,光合速率快,为VOCs 的合成提供能量和碳源[27];同时温度升高会显著提高VOCs 合成酶的活性[28];其次金钱松合成萜烯类化合物储存于树脂管中,其角质层通透性随着温度升高而增加,释放大量VOCs[29]。
NAI产生的天然能量来源包括宇宙射线、紫外线辐射、雷暴和闪电等自然电晕放电、瀑布冲击(勒纳德效应)和植物来源等[14]。森林中NAI的产生主要是通过绿色植物冠层和叶尖的放电以及光合作用的光电效应[12],一些植物的针状叶形有助于空气电离产生负电荷,另外在紫外线作用下植物叶片发生光电效应,使NAI浓度增加。NAI能促进植物生长,在电荷作用下能吸附、沉降空气颗粒物,还能减少空气中微生物的数量[14]。本研究金钱松林内NAI 浓度显著高于空旷地,与冯鹏飞等[30]研究结果一致,群落结构越复杂,NAI浓度越高,由高到低依次为乔灌草、乔灌、乔草、灌草、乔、灌、草和无植被区[13];金钱松林内NAI浓度在夏季达到峰值,可能是夏季降水增多,整体湿度大,原子核凝结半径和结合速率增加导致离子迁移率的降低,当NAI和水分子结合形成负离子团时,其寿命也更长[14];同时夏季强烈光照和高温条件为金钱松叶片的光电效应提供能量,NAI 浓度随着温度和光强的升高而增加[31-32];其次有研究证明游憩林VOCs 总量与NAI 浓度同步变化,酯类物质与NAI 浓度呈显著正相关[12],本研究金钱松VOCs 中酯类含量较高,对NAI的形成有促进作用。
植物通过释放VOCs 来抑制空气中微生物的生长或者是加速附着在颗粒物表面细菌的沉降,最终达到改善空气质量的效果。毛地黄钓钟柳花(Penstemon digitalis)通过释放芳樟醇来保护花组织免受微生物的破坏[33];樟子松(Pinus sylvestris)释放的的萜烯类和五角枫(Acer truncatum)释放的醛类物质对空气中细菌的生长有明显抑制作用[34-35],在夏季抑菌作用最强[34]。本研究发现金钱松林内空气微生物数量显著低于空旷地,夏季数量低于其他两季,可能是金钱松林具有的萜烯类和醛类物质有效抑制了空气微生物的生长,而且夏季金钱松释放的VOCs 含量最多,抑菌效果好。有研究发现5 种针叶树浸提液对空气微生物有抑制作用,并通过单体实验证明柠檬烯、β-蒎烯、壬醛、癸醛和苯甲醛是主要抑菌物质[11]。为了确定金钱松林VOCs 中具体抑菌物质,从林内VOCs 特有的物质和金钱松释放VOCs 中选取含量较高的6 种单体进行微生物的抑制实验,3 种萜烯中,柠檬烯和石竹烯对细菌有显著抑制作用,在高浓度下能抑制真菌生长;3 种醛类对细菌、真菌和放线菌均有显著抑制作用,单体浓度越高,抑制作用越强。因此金钱松释放的VOCs 中柠檬烯、石竹烯、辛醛、壬醛和癸醛可能是对林内空气微生物起抑制作用的主要物质。
综上所述,金钱松释放的萜烯类和醛类化合物对空气微生物有抑制作用,同时金钱松释放的VOCs能促进空气离子化,形成大量NAI,夏季林内VOCs 和NAI 含量均高于其他季节,微生物数量明显减少,可能是VOCs 和NAI 共同作用产生抑菌效果,从而提高林内空气质量。由此可见,在未来园林配置以及森林城市建设过程中,不仅要考虑植物的观赏性,还要结合其释放的VOCs成分和抑菌性,关注其保健方面的研究。
猜你喜欢
作物学报(2022年12期)2022-10-14
江苏农业科学(2021年13期)2021-09-12
科学之谜(2021年4期)2021-07-09
分析测试学报(2020年3期)2020-05-08
林产工业(2020年2期)2020-03-30
国际呼吸杂志(2019年22期)2019-12-09
橡塑技术与装备(2018年21期)2018-02-19
汽车与新动力(2017年6期)2018-01-06
食品工业科技(2014年21期)2014-12-16
应用海洋学学报(2014年4期)2014-11-22
