
核桃细菌性黑斑病菌中脂蛋白预测及其特性研究
2022-01-14 13:28韩长志
江西农业大学学报 2021年6期
覃 悦,韩长志
(1.西南林业大学 生物多样性保护学院,云南 昆明 650224;2.西南林业大学 研究生院,云南 昆明 650224;3.云南省森林灾害预警与控制重点实验室,云南 昆明 650224)
【研究意义】近年来,核桃产业已成为云南破解贫困问题的重要高原特色产业之一,全省90%以上地区均有核桃种植,2017年云南核桃的种植面积(4 300万亩)、产量(115万吨)、产值(315亿元)均居全国之首[1-2]。然而,核桃炭疽病、细菌性黑斑病等病害严重影响着该省核桃产业的健康、有序和快速发展[3]。由黄单胞菌属细菌Xanthomonas arboricolapv.juglandis引起的核桃细菌性黑斑病是世界性的核桃病害,植株受害率高达90%以上[4-5],果实容易腐烂、早落,出仁率和含油率均降低,严重时落果率高达60%~80%。【前人研究进展】目前,国内外学者关于该病的研究,主要集中在病害症状、病原、侵染循环、检测技术、预测预报技术[6]、发病规律及防治[7-9]等方面。植物病原物中的分泌蛋白在实现侵染、定殖寄主植物过程中发挥着重要作用[10],学术界对植物病原真菌、卵菌等真核生物分泌蛋白的预测较多,而对植物病原细菌特别经济林木上的病原细菌研究较少[11]。【本研究切入点】原核生物脂蛋白最早在大肠杆菌中发现[12],之后诸多学者研究发现脂蛋白前体在胞质中合成,可以通过Sec 分泌途径以未折叠形式穿过胞质膜,也可通过双精氨酸转位TAT 途径或SecA 突变体以折叠形式穿过胞质膜。同时,脂蛋白信号肽对于锚定蛋白的正确定位、修饰和转运对细菌维持膜的稳定性、细胞分裂、营养获取等生理功能的发挥具有重要作用[13]。在核桃细菌性黑斑病菌分泌系统中SPaseI和SPaseII等不同信号肽类型的外泌蛋白还未被界定,前人对黄单胞菌中脂蛋白的研究主要集中在相关蛋白的表达、纯化以及结晶等[14]。前期项目组已经明确了CFBP2528、CFBP7179、CFBP8253、DW3F3、J303、NCPPB1447、Xaj417 等7 个核桃细菌性黑斑病菌菌株中分泌蛋白数量及其所具有的特征[15-16]。【拟解决的关键问题】为了更好地解析该病菌的致病机理以及找寻防治该病害的作用靶标药剂,本研究通过LipoP v1.0[17]在线分析软件,选择11 个已经报道全基因组序列的核桃细菌性黑斑病菌菌株开展SPaseII 脂蛋白预测。同时,对以上蛋白所具有的跨膜区结构域、亚细胞定位和理化性质等特征进行明确,并通过遗传关系分析进一步明确该菌中脂蛋白之间的亲缘关系,为深入探索脂蛋白在其致病过程中的作用提供重要的理论支撑。
1 材料与方法
1.1 核桃细菌性黑斑病菌序列来源
核桃细菌性黑斑病菌11 个菌株中的全蛋白组序列来源NCBI 数据库,其菌株编号分别为Xaj 417、CFBP 2528、DW3F3、CPBF 426、CPBF 367、CFBP 7179、CPBF 427、CPBF 1521、CFBP 8253、J303、NCPPB 1447。
1.2 方法
1.2.1 脂蛋白序列获取 利用LipoP v1.0[17]在线程序预测具有脂蛋白信号肽的蛋白质序列。信号肽则用SignalP 5.0 Server[18]进行预测。
1.2.2 跨膜区结构预测 利用2 种不同跨膜区结构预测网站HMMTOP version 2.0[19]等对脂蛋白进行预测。
1.2.3 亚细胞定位分析 利用亚细胞定位分析软件ProtComp v9.0[20]对脂蛋白进行预测。
1.2.4 理化性质分析 利用理化性质测定程序对脂蛋白信号肽蛋白进行测定[21]。
1.2.5 转运肽预测 利用蛋白质转运肽线在分析软件TargetP 1.1 Server[22]对脂蛋白进行分析。
1.2.6 基本特征分析 利用Microsoft Excel 2010统计和分析具有脂蛋白中氨基酸残基和信号肽特征,并采用Origin 2019作图软件做图。
1.2.7 遗传关系分析 利用ClustalX[23]和MEGA X 软件[24]分别对细菌性黑斑病菌中脂蛋白序列开展多重序列比对分析和构建系统进化树。
2 结果与分析
2.1 脂蛋白基本信息
对11 个核桃细菌性黑斑病菌株进行脂蛋白找寻,总共获得1 718 个脂蛋白,每个菌株中平均含有156 个蛋白,所占比例为3.77%(表1)。进一步分析脂蛋白在全部蛋白中所占比例情况,发现菌株CPBF 367 中脂蛋白所占比例最高,为4.02%,而菌株CPBF 1521 中脂蛋白的比例最低,为3.62%(表1),其他菌株中脂蛋白比例则介于上述两者之间。
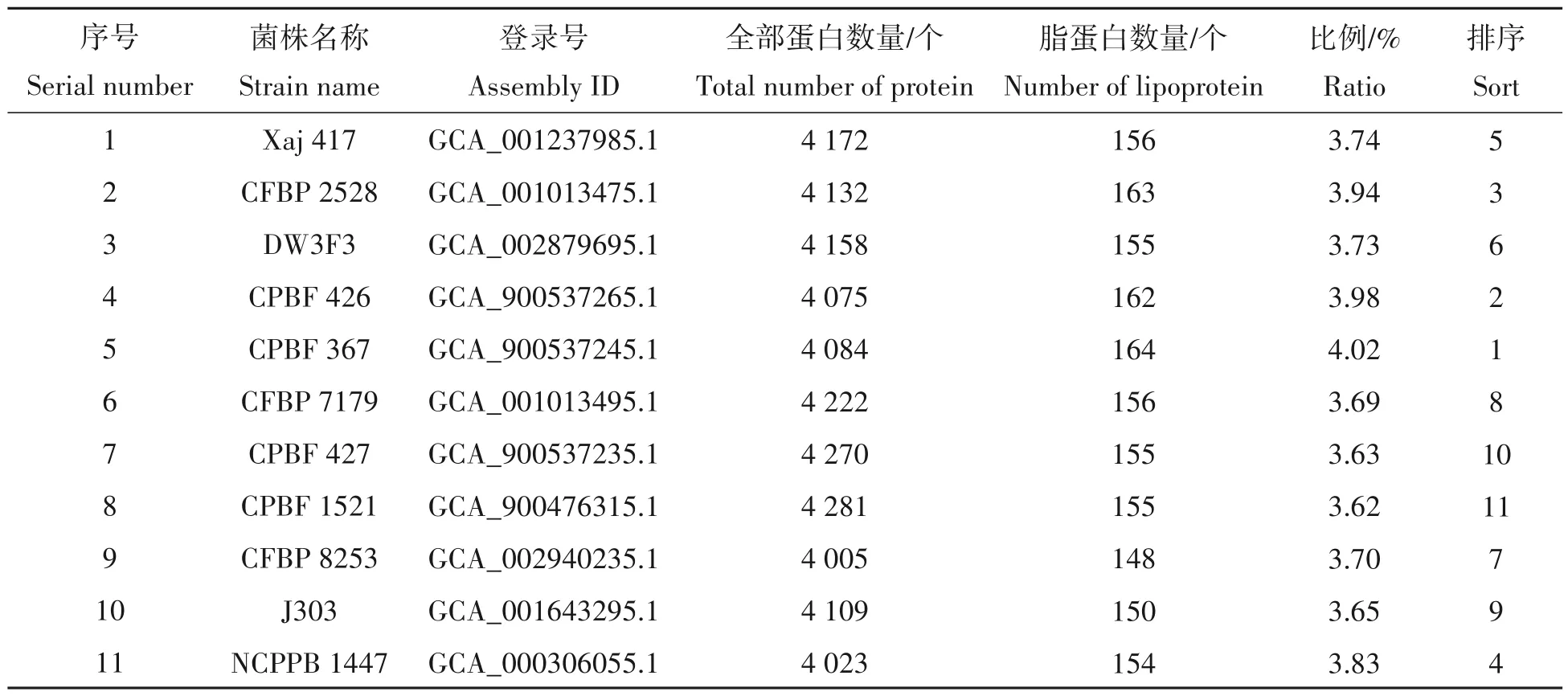
表1 核桃细菌性黑斑病菌脂蛋白的基本信息Tab.1 Basic information of lipoprotein in X. arboricola pv. juglandis
同时,分别对每个菌株中所含有的脂蛋白氨基酸长度以及其组成进行统计分析,结果表明,脂蛋白氨基酸长度集中于150~450 aa,其中长度在100~200 aa 的脂蛋白数量最多,平均为43 个,所占比例为27.56%(图1A)。在上述核桃细菌性黑斑病菌中,脂蛋白氨基酸组成以A(丙氨酸)所占比例最高,其范围为14.4~14.8%,其次为L(亮氨酸)和G(甘氨酸)氨基酸残基,含量范围为8.1~9.1%;而C(天冬氨酸)、H(组氨酸)、M(甲硫氨酸)、W(色氨酸)氨基酸残基含量最低,平均含量仅为1%~2%(图1B)。上述分析结果说明,脂蛋白作为重要的蛋白类别,其氨基酸长度较小。
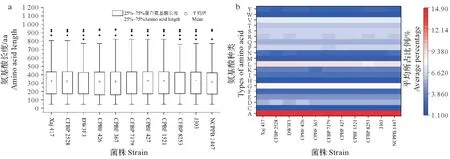
图1 核桃细菌性黑斑病菌脂蛋白的氨基酸长度及其组成情况分析Fig.1 Amino acid length and composition analysis of lipoprotein in X.arboricola pv.juglandis
2.2 脂蛋白跨膜结构域分析
基于HMMTOP version 2.0 跨膜结构域分析,1 718 个蛋白中具有1 次以上无跨膜结构域的蛋白数量仅有28 个,具体而言,在Xaj 417、CFBP 2528、DW3F3、CPBF 426、CPBF 367、CFBP 7179、CPBF 427、CPBF 1521、CFBP 8253、J303、NCPPB 1447中具有1次以上无跨膜结构域的蛋白序列分别有2、2、2、3、3、3、3、3、2、3和2条(表2),而其他蛋白则均不含有跨膜结构域。
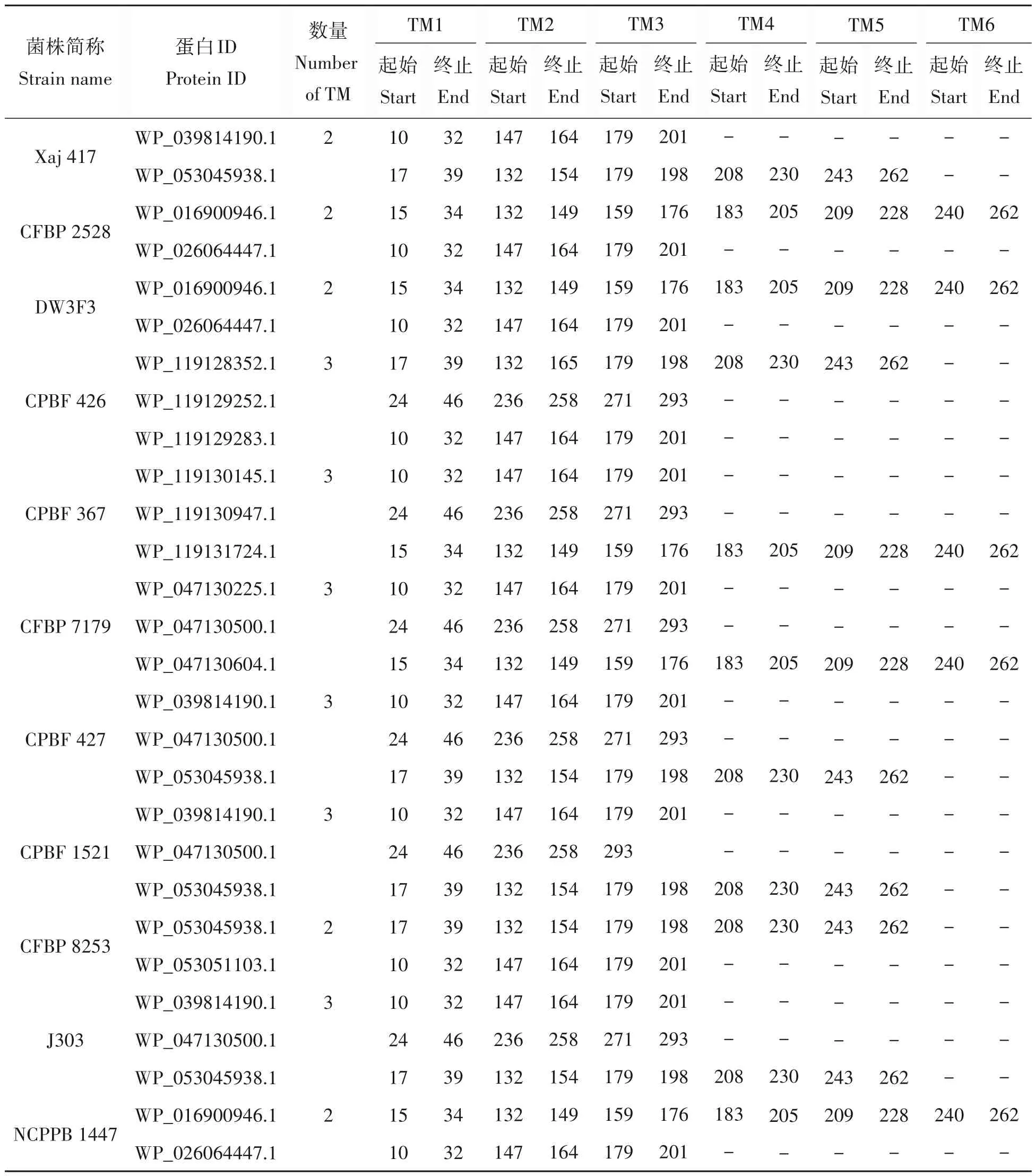
表2 核桃细菌性黑斑病菌中具有多个跨膜结构域的脂蛋白情况Tab.2 Prediction of transmembrane of lipoprotein in X. arboricola pv. juglandis
2.3 脂蛋白亚细胞定位分析
对11个核桃细菌性黑斑病菌中脂蛋白序列开展亚细胞定位情况预测。结果显示,在所获得的1 718个脂蛋白中,定位在线粒体的脂蛋白数量最多,为66~75个;其次为细胞外,为50~62个;其他蛋白则定位在细胞核、质膜、细胞质、内质网、过氧化物酶体、溶酶体及高尔基体(图2)。
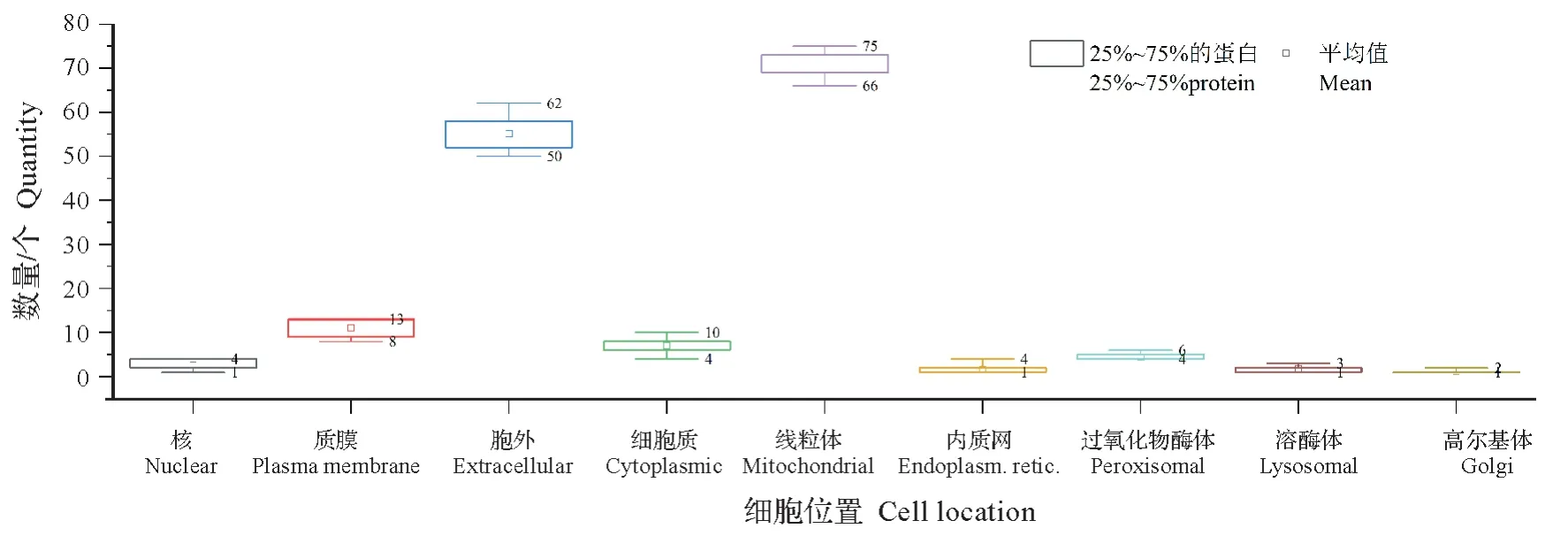
图2 核桃细菌性黑斑病菌中脂蛋白亚细胞定位情况分析Fig.2 Subcellular localization of lipoprotein in X.arboricola pv.juglandis
2.4 脂蛋白理化性质分析
进一步对1 718 个脂蛋白理化性质进行分析,结果显示,尤以理论等电点位于5.51~6.00 的脂蛋白数量最多,平均数量为32 个,所占比例为20.4%,其次为9.01~9.50,平均数量为18 个,所占比例分别为11.54%(图3A);就蛋白质不稳定性系数而言,每个菌株中平均有74个脂蛋白的不稳定性小于40,所占比例为47.44%,其余蛋白不稳定性系数大于40(图3B);就总平均亲水性(GRAVY)而言,总平均亲水性小于0的蛋白数量大于总平均亲水性大于0的蛋白数量,平均为-0.181(图3B),属于亲水性蛋白;就脂肪族氨基酸指数而言,共有139个脂蛋白氨基酸数量大于70 aa,所占比例为89.1%,其中在80~90 aa的脂蛋白数量最多,为60 个,所占比例为38.46%(图3C)。通过上述分析,明确11 个核桃细菌性黑斑病菌中的脂蛋白在理论等电点、不稳定系数以及总平均亲水性等方面存在着一定的差异,但是总体而言供试菌株中的脂蛋白在理化性质方面具有趋同的特性,该分析结果对于深入解析某一菌株中脂蛋白结构及功能从而研究其他菌种中脂蛋白特性提供了重要的理论支撑。

图3 核桃细菌性黑斑病菌中脂蛋白基本理化性质分析Fig.3 Theoretical isoelectric point analysis of lipoprotein in X.arboricola pv.juglandis
2.5 脂蛋白的信号肽及转运肽特征
通过SingnalP 5.0 分析,在11 个核桃细菌性黑斑病菌中,分别仅有7、7、8、9、13、9、9、9、7、8、8 条蛋白不具有信号肽序列,不属于典型分泌蛋白,而其他序列则具有信号肽序列。进一步对具有信号肽脂蛋白基本特征分析,结果表明,信号肽长度位于20~25 aa 的数量最多,平均为78 个,所占比例为50.24%,且25%~75%的脂蛋白信号肽预测可靠性位于25%~50%(图4A)。对信号肽切割位点特征进行分析,在上述1 718个脂蛋白中,尤以A(丙氨酸)所占比例最高,为21.52%,F(苯丙氨酸)、I(异亮氨酸)、W(色氨酸)和Y(酪氨酸)的平均含量最低,仅为0.27%、0.34%、0.1%和0.44%(图4B)。
此外,在信号肽切割位点-3、-2、-1、1、2 处,A(丙氨酸)、A(丙氨酸)、C(天冬氨酸)、A(丙氨酸)、P(脯氨酸)的含量最高,平均所占比例分别为38.08%、22.45%、28.19%、13.87%、20.15%(图4C)。氨基酸的组成在-3 和-1 位上相对比较保守,属于A-X-C 类型,在-3 位上,除A(丙氨酸)之外,C(天冬氨酸)、G(甘氨酸)、T(苏氨酸)、V(缬氨酸)平均所占比例分别为26.06%、7.7%、10.55%、5.8%,其余氨基酸含量所占比例均小于1%;在-1 位除了C 外,氨基酸A(丙氨酸)、G(甘氨酸)、P(脯氨酸)、S(丝氨酸)、T(苏氨酸)平均所占比例分别为28.04%、20.35%、1.36%、14.86%、5.37%,其余氨基酸含量所占比例均小于1%(图4C)。

图4 核桃细菌性黑斑病菌中脂蛋白的信号肽基本特征分析Fig.4 Theoretical isoelectric point analysis of lipoprotein in X.arboricola pv.juglandis
对上述1 718个脂蛋白开展转运肽分析,结果发现,具有转运肽的脂蛋白数量较少,且预测可靠性不高。除DW3F3菌株外,其他10个菌株中的一个蛋白(其蛋白ID为WP_016904744.1)具有转运肽,转运肽定位点均位于26~27,定位点氨基酸残基均为ARS-GH(表3)。该结果再次印证了之前对于核桃细菌性黑斑病菌中脂蛋白理化性质的分析,即该病菌中脂蛋白具有较大的相似性。

表3 核桃细菌性黑斑病菌中脂蛋白的潜在转运肽的可能性预测Tab.3 Possibility prediction of transit peptides of lipoprotein in X. arboricola pv. juglandis
2.6 脂蛋白的遗传关系分析
细菌性黑斑病菌脂蛋白信号肽1 718 个蛋白序列,通过氨基酸比对,去除重复序列后获得687 个蛋白序列,进一步利用ClustalX[23]和MEGA X 软件[24]分别进行多重比对分析和构建系统进化树。结果显示,细菌性黑斑病菌中的脂蛋白序列根据遗传关系可明显的分为6 个大类,表明该菌中大部分脂蛋白之间有较高的同源性以及较近亲缘关系(图5),其中Ⅱ类和Ⅳ类所含蛋白数量最多,分别有171 和124 个蛋白,Ⅲ类所含蛋白数量较少,有64 个蛋白。

图5 细菌性黑斑病菌脂蛋白信号肽序列之间的遗传关系Fig.5 The genetic relationship between the lipoprotein sequences in X.arboricola pv.juglandis
3 结论与讨论
本研究通过LipoP 分析,首先明确了11 个核桃细菌性黑斑病菌菌株中的脂蛋白情况,总共获得1 718 个脂蛋白,平均每个菌株含有156 个蛋白,所占比例为3.77%,氨基酸长度集中于150~450 aa;其次,通过在线分析软件HMMTOP、SignalP 等,明确脂蛋白具有1 次以上跨膜结构域不多,多数脂蛋白定位在线粒体核胞外,并且在理论等电点、不稳定系数以及总平均亲水性等方面存在着一定的差异,但是总体而言上述菌株中的脂蛋白在理化性质方面具有趋同的特性,且仅有少数蛋白不具有信号肽序列,不属于典型分泌蛋白,而具有转运肽的脂蛋白数量也相对较少,预测可靠性相对较低。同时,具有脂蛋白信号肽的蛋白序列是以信号肽切割位置为界,在-3、-2、-1、1位有4个保守的氨基酸残基,一般为LA(G/A)C,信号肽长度一般为20 个氨基酸残基[25],然而,本研究中所获得的诸多脂蛋白信号肽切割位点多为AACA,且在-3 和-1 位上相对比较保守,尚不清楚是何种原因造成该现象出现,有待于今后进一步解析。最后,通过遗传关系分析发现细菌性黑斑病菌中脂蛋白序列之间在具有较高同源性以及较近亲缘关系的同时,还可能面临着诸多选择压力,推测可能在进化过程中为了应对不同的生存环境而产生了较大的分化。上述研究结果为深入研究细菌性黑斑病菌脂蛋白功能提供重要的理论借鉴作用。同时,核桃细菌性黑斑病菌分泌系统中SPaseI 和SPaseII 等不同信号肽类型的外泌蛋白在同源关系上、功能类别上是否保守,是否都具有上述研究结果的(种内、种间的)独特生物学功能还有待进一步深入研究。
一般而言,植物病原真菌在实现成功侵染植物过程中,经历了克服植物固有防御反应、基础免疫反应以及系统免疫等一系列防卫反应,从而实现对寄主植物的侵染、定殖以及系统操控等。脂蛋白几乎存在于所有细菌中,其通过N 端脂质结构锚定在膜中,并参与诸多重要的代谢过程,包括营养吸收、信号转导、粘附、结合和孢子形成、参与抗生素抗性的生成、转运和蛋白质的胞质折叠[26]。作为外泌蛋白中的一类,具有脂蛋白信号肽的蛋白则对于锚定蛋白的正确定位、修饰和转运,从而对细菌维持膜的稳定性、细胞分裂、信号转导、营养获取等生理功能的正常发挥具有非常重要的作用,同时还在细菌生理、致病性和耐药性等方面发挥着重要作用。由于脂蛋白是通过信号肽合成,并将其固定在细胞质膜上,而脂蛋白结构域存在于细胞周质或细胞外等特征,因此多将其作为靶标用作药物开发[27]等。近年来,国内外学者对植物病原真菌、卵菌及细菌等的分泌蛋白预测较多,包括本研究小组在内的诸多国内外学者对植物生物分泌蛋白进行了大量的生物信息分析,如对真菌[28]、卵菌[29]以及枯草芽孢杆菌、细菌性黑斑病菌[15]等植物病原物。前人对脂蛋白的研究主要集中在相关蛋白的表达、纯化以及结晶等[14]。本研究通过对核桃细菌性黑斑病菌脂蛋白的预测及特征分析,发现菌株中的脂蛋白之间具有较大的相似性,然而,上述研究仅针对1种病原物的预测分析,尚缺乏对于多种植物病原物中脂蛋白预测及特征分析等工作。由于天然状态下的生物多糖大分子的高聚合度和不可溶性,微生物的多糖降解酶需要分泌到胞外才能与底物接触,实现多糖的降解。酶组分在胞质中被合成前体到分泌到胞外或与细胞膜结合的过程中,脂蛋白在此过程中发挥了哪些作用?脂蛋白是否参与或影响胞吞作用?脂蛋白在细菌性黑斑病菌致病过程中占据怎样的地位等等问题还有待进一步验证。
猜你喜欢
当代水产(2022年2期)2022-04-26
生物技术通报(2021年6期)2021-08-11
健康体检与管理(2021年10期)2021-01-03
湖北农机化(2020年4期)2020-07-24
当代水产(2020年4期)2020-06-16
当代水产(2020年4期)2020-06-16
当代水产(2020年3期)2020-06-15
娃娃乐园·综合智能(2019年12期)2020-01-15
浙江农业学报(2017年3期)2017-04-08
中国烟草学报(2016年1期)2016-11-16
