
独一味逆境生理研究进展
2022-01-12 05:39路岳衡张凯敏耿贵工
中国野生植物资源 2021年12期
路岳衡 ,张凯敏 ,耿贵工 ,乔 枫 *
(1.青海师范大学生命科学学院,青海西宁810008;2.青海省青藏高原药用动植物资源重点实验室,青海西宁810008;3.青海大学农林科学院,青海西宁810016)
独一味(Lamiophlomis rotata)又名独步通、野秦艽、麦朵昌巴、吉布孜[1],归属唇形科,多年生[2]草本植物,原属糙苏属,现已从糙苏属中分离,隶属于独一味属唯一物种。广泛分布于我国西北地区,大部分集中分布在西藏,少部分零星分布在我国青海省、四川省、甘肃省和云南省[3],其生长海拔高度在3 100 米到5 100 米,生长环境多见于风化强度较高的碎石滩、退化的石质高山草甸、干旱化的河滩等地[4],贴地生长。独一味含有多种氨基酸和多种微量元素[5],环烯醚萜类化合物是其中主要的化学成分,如螃蟹甲苷、胡麻属苷、独一味素A等,其余化学成分为苯乙醇苷类(天人草苷B、独一味苷A 等)、黄酮类(木犀草素、芹菜素等)及脂肪酸、酚酸类化合物等[6-8],是我国西北地区特有的一种重要的药用植物[9]。独一味药材外观表面呈现淡黄棕色,质地坚硬,干枯,气味略显腥臭,味道微苦[10],其药理活性主要集中在治疗外伤出血[11]、减缓疼痛[12-13]、消炎抗菌[14]、抗肿瘤及增强人体免疫力[15]等,在临床有很好的开发应用前景[16],也是藏族、土族、门巴族等少数民族广泛使用的中草药品种之一[17]。
独一味株高25~100 mm,根及根茎伸长且较为粗厚,通常为圆柱形,直径一般为 20 mm 左右[18];表面通常具有棱,棱上有皱纹,颜色为棕黄色,形状大多数为扇形或者肾形,长60~130 mm,宽70~120 mm;叶片于基部丛生,通常4 枚,呈现两两相对对称分布[19]。轮伞花序长35~70 mm,呈短圆锥状;花冠长约12 mm,宽2 mm 左右,唇形近圆形,颜色一般为紫红色。通常在6~7月开花,果期在8~9月[20]。
独一味由于其生长环境的独特性,通常面临着多种恶劣的环境胁迫,诸如寒冷、高强度的紫外线辐射、干旱等,且其产区的绝对无霜期较短,独一味幼苗在无霜期内积累的生长量不足,抵抗环境胁迫能力较弱,在越冬后大量死亡,成活率不高,导致其产量较少[21]。此外,独一味作为许多成方药品的主要原料且无成瘾性的特点受到了人们的广泛关注,使得很多人受经济利益的驱使对独一味进行过度挖掘,超出了其本身可循环能力,进而加速了野生独一味资源的濒危进程[22]。在面临着濒危灭绝的风险下,当务之急是改善独一味的野生抚育和提高人工驯化种植栽培技术[23],并通过保护生物学的研究对其采取合理的保护与资源可持续性利用。
1 逆境胁迫
1.1 低温胁迫
过低的温度不利于独一味生长,是抑制其生长最重要的一个环境因素。低温寒害是高海拔地区植物常见的胁迫损害。一般来说,当温度处于零度及以上低温时(冷害),独一味具有较强的环境适应能力,其长势良好,对独一味幼苗或成株的影响不是很大,但当温度处于零度及以下低温时(冻害)[24],独一味抗逆能力下降较为明显,对其植株的迫害较为显著。
1.1.1 低温对独一味细胞膜系统的影响
细胞膜作为细胞与环境之间的接触面,是独一味遭受低温胁迫时最先被侵害的部位。低温会破坏独一味的细胞膜系统,导致其细胞质膜的透性增加,使细胞质膜的选择透过性下降,导致电解质会不同程度地外渗,电导率也会随之增大。造成这些现象的根本原因主要是由于低温会使细胞膜保护酶活性降低、内源抗氧化剂含量下降,致使膜脂过氧化能力降低,进而引起膜结构的破坏,导致独一味的损伤和死亡[25]。但有研究发现[26],在初遇低温时,独一味的相对电导率增大较为显著,但随着胁迫时间的不断推移,电导率的增长趋势趋于缓慢。推测其原因与膜系中磷脂及脂肪酸的不饱和性有很大程度的相关性。可能是由于低温使膜脂中不饱和度较高的脂肪酸和磷脂的含量增加,而随着不饱和脂肪酸和磷脂的含量上升,可降低膜结构变化所需要的温度,增强膜的流动性,提高其抗冷能力,说明独一味可以慢慢适应低温环境。
1.1.2 低温对独一味抗氧化系统的影响
自由基生物学的许多研究成果表明[27],环境胁迫对植物体造成的损害大多通过破坏自由基的代谢平衡来实现。通常,植物细胞中自由基的含量很低,若植物细胞产生多余的自由基,同样也会通过其他途径来对其清除,达到一种动态平衡状态,并不会影响植物体的生长。对于独一味来说,其体内维持自由基代谢平衡的防御系统主要有两类:酶促防御系统和非酶促防御系统。酶促防御系统主要包括过氧化氢酶(Catalase,CAT)、超氧化物歧化酶(Superoxide dismutase,SOD)、过氧化酶(Peroxidase,POD)等抗氧化酶类所进行调节的防御系统;非酶促防御系统是指由还原性谷胱甘肽、维生素A、维生素C、维生素E 和抗坏血酸等几十种广泛存在于植物组织中的具有还原活性的物质所构成的,两个防御系统起到协同作用共同维持独一味体内的自由基代谢平衡(图1),故这些物质的含量高低与活性大小对独一味清除体内多余的自由基含量起到至关重要的作用。以酶促防御系统为例:SOD 可以防止O2-转变为毒性较强的OH-,转而通过歧化反应使O2-转变为毒性较低的H2O2和O2,再通过CAT的催化作用,结合POD 进一步清除H2O2及代谢过程中产生的其他过氧化物,使其转变成无毒的水(图2),从而削弱它们对机体的攻击力,减少了对膜结构和功能的损害,从而避免了自由基对植株的伤害而使其得到保护。当独一味遭受低温胁迫时,其体内SOD、POD、CAT 等酶活性降低甚至失活,导致细胞内产生的多余的自由基不能被及时清除,使自由基大量累积,而自由基的累积会持续对膜系统造成损伤,引起膜脂过氧化,使细胞膜的渗透率增加,细胞膜的选择透过性下降,导致外界钙离子大量内流,内流的钙离子与钙调蛋白相互结合,激活了磷脂酶A2,引起不饱和脂肪酸的解脱,不饱和脂肪酸又在脂氧合酶的作用下形成羟基过氧化物,它能自发分解出丙二醛(Malondialdehyde,MDA),MDA 的累积可导致DNA 结构的损伤,蛋白质的变性,磷脂的降解,细胞膜的崩解,进而导致细胞损伤甚至死亡[28]。
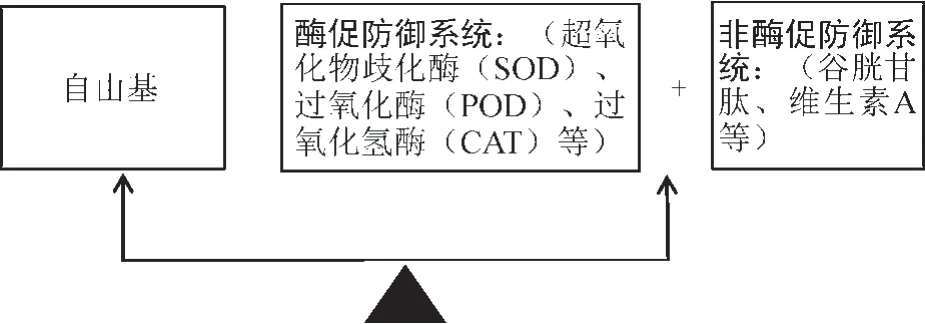
图1 自由基代谢平衡Fig.1 Free radical metabolic balance
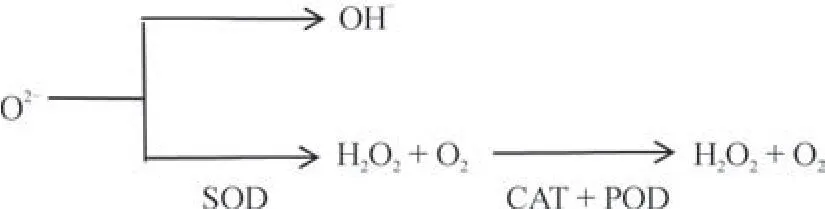
图2 酶促防御系统Fig.2 Enzymatic defense system
1.1.3 低温对独一味渗透调节物质的影响
植物细胞内的渗透压代谢平衡影响着植物生长,低温可减弱植物细胞质维持水的能力,使组织中非结冰水的含量减少,导致细胞失水从而使蛋白质和蛋白质胶体容易发生凝固反应而不利于植株生长,而植物体内的某些渗透调节物质可以维持植株的渗透压平衡,诸如无机离子、可溶性糖等,其含量与独一味的抗寒性呈正相关关系。当独一味处于低温条件下时,其体内可溶性糖含量增加,可能原因是因为低温诱导了独一味水解酶的活性,加速了淀粉的水解,同时在低温下一些消耗糖的生理活动减弱,导致可溶性糖的积累,从而提高了细胞汁液浓度,防止细胞过度脱水,降低了细胞质的冰点[29],从而起到保护独一味的作用。
1.1.4 低温对独一味光合作用的影响
光合作用是植物积累碳水化合物以及为植物生命活动提供能源物质的主要途径,贯穿植物整个生命进程,光合色素尤其是叶绿素,在植物的光合作用中对光能的吸收、传递和转化起着极为重要的作用。若独一味在其遭遇低温前未达到一定的生长量,在条件适宜的情况下也很难恢复生长,而在低温情况下维持高的叶绿素含量是独一味在低温条件下能达到一定生长量的前提,一旦条件适宜,迅速恢复生长发育。但当温度过低时,其叶绿素含量下降,活性降低,叶绿体功能紊乱,植物进行光合作用的进程减慢,减少了次生代谢物质的含量[30],使其很难达到一定的生长量,从而不利于独一味的生长。
1.1.5 独一味染色体倍性与其耐低温适应性的关系
通过对高海拔地区诸多植物染色体倍性的测定发现(星叶草、固沙草、独一味等),很多植物的染色体构成均为二倍体,极少数植物的染色体倍性表现为多倍体,可能原因是由于当海拔过高时,多倍体植物抵抗寒冷、吸收氧气的能力就会下降,竞争不过同地区的二倍体植物,种群更新缓慢,在经过漫长的自然选择后而被淘汰,而二倍体植物多存活下来,故二倍体植物相对于多倍体植物能更好的适应寒冷缺氧的环境[31]。而独一味的染色体倍性为二倍体,因此在低温环境下其染色体的特性有利于其生长发育。
1.2 干旱胁迫
干旱的土壤首先改变的是土壤的紧实度,接着再引起土壤其他理化、生物性质的变化:如土壤所含的营养元素降低(磷、钾、锰等)、土壤的最大吸湿量、含水量大幅度减少,总孔隙度减少,通气性降低,土质板结,水分的渗透阻力增大等[32],使得很多植物很难在干旱的环境下生存,但颇为有趣的是在干旱环境下独一味有一套独特的生长适应机制,使其长势良好。
1.2.1 独一味的自身形态结构
独一味根茎粗壮,直径在7~20 mm;根茎伸入土壤较深,一般深度在50~100 cm 左右,实际已深入土壤母质层,有利于其在干旱环境下更好的从较深土壤中吸收水分和养分。独一味叶片较大多皱且贴地平展,能有效防止植株地表土壤水分及土壤毛细管持水蒸发,被毛、叶上表面非腺毛较多在白天可以减少蒸腾[33]。因此,由于独一味独特的形态结构,可以使其在干旱的土壤中有效保持水分,减轻因干旱而导致的缺水问题。
1.2.2 独一味根系分泌物
独一味根系分泌物的主要成分包括碳、氮、脂质类(亚油酸甲酯、亚麻酸乙酯等)、烷烃类化合物(十九烷、正十七烷等),另外还含有少量酚类。研究发现[34],独一味根系分泌物含量的多少影响着独一味的生长,当根系分泌物浓度较低时具有促进其生长的作用,而当根系分泌物浓度较高时则会抑制独一味的生长,并随浓度升高抑制作用增强。随着干旱程度的增加,独一味根系分泌物含量减少,从而对独一味植株的生长起到促进作用。
1.2.3 独一味叶绿体的变化
通常情况下,随着干旱胁迫加重,可造成植物叶绿体、线粒体膜系统受损,引起植物光合作用减弱、能量不足、淀粉粒减少、叶片颜色改变、叶绿体脱离细胞壁等一系列变化。但独一味多有不同之处:独一味为适应干旱胁迫,调节叶绿体的形状,使叶绿体呈现出梭形或长条形,保护膜系统不受到破坏,且所有叶绿体轮廓清晰,片层清晰可视,垛叠区片层交叉紧密,未出现松散现象。同时,基粒位置也加以调整,增加了叶绿素的合成,以充分利用光能,产生较多的光合作用产物淀粉粒(植物代谢的能源物质),以此来满足自身新陈代谢的旺盛,能量消耗的增多[35],从而抵御退化生境。
1.2.4 独一味生理指标的变化
在干旱条件下,植物器官或细胞通常伴随膜脂过氧化反应,该反应的最终产物丙二醛可与膜保护酶结合并发生交联作用而导致酶失活,损害了生物膜的正常结构和功能,同时植株体内总碳、总氮含量减少,植物细胞缺乏足够的水分支持而出现渗透胁迫。但独一味为了适应干旱环境,分别通过增强SOD、POD酶(植物体内比较重要的两个自由基清除酶)的活性,来有效清除独一味体内的活性氧自由基,降低MDA 的含量,从而保护了生物膜的正常结构和功能;面对渗透胁迫,脯氨酸主要起着渗透调节作用,通常情况下植物体内脯氨酸含量较低,但在干旱胁迫下,独一味会增加脯氨酸的含量至之前的十倍甚至百倍以上以维持细胞的渗透调节能力[35],从而更好的适应干旱环境。
1.3 紫外胁迫
随着海拔高度的升高,光照强度也随之增强,紫外辐射增强,特别是紫外辐射中的UV-B 辐射对植物的影响最大,可影响植物的生长发育以及生理生化反应,严重者还可导致植物死亡,具体表现在植株的株高、根粗、根茎芽数、叶展面积、叶片生物量、花序数、花序生物量、地上部生物量等都随着紫外辐射的增强而下降[36],仅对叶片数无显著影响;还会诱发植株体内的基因发生突变而导致其翻译的蛋白质结构改变或功能发生变化,从而影响着植物的生命活动。而独一味为抵抗紫外辐射,一方面在形态上通过增加叶片厚度和减少叶面积等机械防御直接应对,并且其叶密被白色疏柔毛,具皱较厚,可以一定程度折射紫外光线并抵抗UV-B 辐射对叶细胞的伤害[37];另一方面由于光照强度的增强,一定程度上刺激了独一味光合作用效率提高,增加了叶绿素的合成,产生了更多的次级代谢产物,且分别测定独一味不同器官部位的次生代谢产物含量发现,独一味地下根部和地上叶部的次生代谢产物含量的积累有显著差异,其叶中次生代谢产物含量明显比根部高,可能是由于地上叶直接接触紫外辐射,叶比根积累更高浓度的次生代谢产物含量从而降低高山上紫外线直接照射对其造成的伤害[38]。表明独一味通过其独特的形态结构以及特有的生理生化特点来适应高强度的紫外辐射,对防治紫外辐射的损伤具有显著作用。
1.4 外源钙离子对独一味抗寒性的影响
1.4.1 钙离子的功能
在植物体内有很多种化学元素,但作为植物生长发育不可或缺的元素仅有16种,分别是碳(C)、氢(H)、氧(O)、氮(N)、磷(P)、钾(K)、钙(Ca)等。而Ca2+可作为植物细胞信号传导系统中的第二信使参与细胞的信号转导,它不仅可以提高植物的抗病害能力,同时还可以调节无机离子运输,对植物产生的许多生理响应以及多种代谢过程都起着十分重要的作用[27]。通常情况下,植物细胞质基质中的Ca2+浓度与细胞外的Ca2+浓度维持着一种动态平衡,当植物感受到外界刺激时,细胞壁的pH 值及质膜内外的电位势发生变化,从而产生电势差使大量Ca2+内流到细胞中,增大了细胞内的Ca2+浓度,增加的Ca2+通过与植物体内的钙调素(能与钙离子相结合的蛋白质,在不结合Ca2+的条件下不具备活性,也叫钙调蛋白)相结合,使钙调素的构象发生改变,从而使结合了Ca2+的钙调素具备活性,成为一些酶的激活物,进而与相应的酶结合,结合后又引起酶的构象发生变化,使相应的酶从无活性状态转变为有活性状态,并继续向下游传递并级联放大,从而激发一系列生理生化反应[28]。
1.4.2 外源钙离子对低温胁迫下独一味的影响
当独一味处于低温条件下时,适宜浓度的CaCl2溶液可以增强独一味植株在低温下的生存能力,其原因一方面是Ca2+可以调节独一味细胞内的渗透压平衡(增加可溶性糖含量)以及提高SOD、CAT、POD等抗氧化酶的活性来减轻自由基对细胞膜的损害,保证细胞膜的完整性;另一方面Ca2+通过束缚带电荷的磷脂来诱导类脂更紧密的排列,增强膜的选择透过性,减少电解质的渗漏,使细胞膜的相对电导率和丙二醛含量维持在较低水平,从而减轻了对细胞的氧化损伤,降低了细胞膜系统的损害程度,保护了膜系统的稳态性和完整性。同时Ca2+对独一味体内的叶绿素具有保护作用,使其经历低温胁迫后仍然维持较高的叶绿素含量,是保证独一味在低温环境下具有一定生长量的前提,使独一味能够维持正常的生理活动,从而有效提高了独一味对低温的抵抗能力[29]。
研究发现[30],当对独一味喷施 15 mmol·L-1的CaCl2溶液时,其对低温的抵抗效果最佳,当喷施浓度继续升高时,外源Ca2+促进独一味抵抗低温的能力减弱,独一味各项生理指标又逐渐下降,说明外源Ca2+帮助独一味抵抗低温胁迫的缓解作用也有一定的局限性,可能原因是因为当喷施浓度过大时,过量的Ca2+得不到及时清除,致使其长时间停留在胞质内引起细胞的反馈调节,造成细胞骨架和膜结构的破坏,使膜透性发生变化,最终引起Ca2+中毒而使细胞内物质代谢失衡。说明要有效提高独一味的抗寒能力还需适宜浓度的外源钙离子[30]。
2 独一味的遗传多样性
从王京[39]等人对青海省玉树地区10 个不同种群共300 株野生独一味的遗传多样性分析可知,其多态位点率高达99.47%,独一味10个种群的总遗传变异Ht 为29.04%,每个种群的平均遗传变异Hs 为21.80%;有24.93% 的基因变异存在于种群间,75.07%的基因变异存在于种群内,基因流Nm 为1.505 5。在刘继梅[40]等人对分别来自青海省、西藏自治区、云南省共8 个不同种群188 个个体的野生独一味进行遗传多样性分析可知,独一味8 个种群的总遗传变异Ht 为78.7%,每个种群的平均遗传变异Hs 为54.6%,有36.46%的遗传变异存在于种群间,63.58%的遗传变异存在于种群内。二者对于独一味遗传多样性的分析结果整体相差不大,故可得出独一味的遗传多样性相对较高,说明独一味的进化潜力很大,这对于独一味适应恶劣多变的自然环境是非常有利的,使其能够在漫长的自然演变进化历史中,更好的形成自然生理与环境相互适应机制,提高自然成活率,从而保证独一味的生长和繁殖。但对于某些地域的独一味来说其遗传多样性却相对较低,分析其原因发现遗传多样性较低的区域主要有两个方面:一是人为活动较为频繁的区域,二是地震多发地带。因此,为了更好的实施对独一味资源的合理保护与开发,就需要运用具体合理的措施,如迁地保护或就地保护,制定适宜的育种方案等,以此来保护独一味的遗传多样性,提高独一味对环境的适应能力。
3 总结与展望
植物在遭受逆境胁迫时往往表现出生长迟缓、植株矮小、代谢能力下降、生长量不足等特点。但独一味在面对逆境胁迫时可通过代谢调节、生长发育调节、渗透调节等多种方式来提高其抗逆能力,减少胁迫造成的损害,保证独一味的生长发育。同时,再结合对独一味遗传多样性的分析,以期为独一味资源的合理开发应用与保护提供理论参考,研究其抗逆性,也为改善独一味野生抚育和提高人工驯化种植栽培技术提供科学理论依据。
综合已有文献分析,目前国内外对独一味的研究主要集中在其药理活性、临床医学应用以及化学成分的分析方面,对独一味潜在的抗逆基因以及抗逆基因的表达调控等方面仍未被探明,探索独一味植物的抗逆性基因及其表达方式或其分子生物学机理,将有利于展开分子育种、分子杂交等优良品种的开发。同时,研究独一味的抗逆机制,也可为其他高原植物的抗逆机制提供参考,从而为培育出更多的具有抗逆特点的植物提供更多的理论依据。
猜你喜欢
真空与低温(2022年6期)2023-01-06
真空与低温(2022年5期)2022-10-13
当代水产(2020年4期)2020-06-16
当代水产(2020年3期)2020-06-15
劳动保护(2018年8期)2018-09-12
科学中国人(2018年8期)2018-07-23
中国资源综合利用(2017年4期)2018-01-22
红领巾·萌芽(2017年5期)2017-06-23
爆笑show(2016年7期)2017-02-09
少儿科学周刊·儿童版(2015年10期)2015-11-07
