
‘长林3 号’油茶传粉生物学特性与繁育系统的研究
2021-12-30 02:56陈文波罗家玲
凯里学院学报 2021年6期
蒋 瑶,陈文波,罗家玲
(凯里学院,贵州凯里 556011)
1 前言
油茶Camellia oleifera,又名油茶树、茶籽树、白花茶等,是山茶科Theaceae 山茶属Camellia常绿阔叶小乔木.油茶种子内含高达80%以上油酸,与椰子树、油棕树和油橄榄树并称为世界四大木本油料树种[1].主要分布于亚洲的热带和亚热带地区海拔100−500 m 的阳坡或半阳坡(坡度<25o)的山地[2−3].
油茶在我国栽培历史悠久,主要分布于长江流域以南的亚热带地区,贵州省主要集中分布于黔东南苗族侗族自治州的玉屏、锦屏、天柱、黎平等县[4].至2017 年,我国油茶栽培面积已高达400×104hm2,年收入约200 亿元[5].种植油茶能够调整农业经济结构,带动地方经济,对促进农村脱贫致富具有重大意义.
本试验所用油茶品种为‘长林3 号’,具有长势旺盛、高产稳产、抗病虫性强、含油率高等优点[6].该品种油茶喜阳光、湿润,耐贫瘠,怕寒冷[6],生命周期长,秋花秋实,开花时间从当年10 月至次年的1月,整个花期为80 d,单株为20 d,单花6−7 d[7].
近年来关于油茶的研究均为常规培育优质丰产技术,国内林业工作者通过无性系进行品种选育来解决提高油茶坐果率和花多果少的问题[8].探索丰产栽培[9]和病虫害防治技术[10]及使用组织培养[11]、油茶分子生物学[12]等技术进行的研究有很多,但仍未能有效解决油茶花多果少的低产问题.
繁育系统是当今生物学研究中较为活跃的内容之一,它指所有影响后代遗传组成的有性特征总和,包括花部综合特征、花部性器官的寿命、访花昆虫种类、开花样式以及交配系统等.
花粉必须在具有活力时到达适宜的接受柱头方能完成授粉过程,具有接受花粉的适宜柱头的花朵即处于柱头可授粉期.花粉保持活力的时间长短与柱头可授粉期的时间长短结合在一起,深刻影响着植物的自花传粉率与授粉成功率,特别是异花传粉的植物.因此,花粉活力与寿命、柱头可授性、花期相遇概率以及杂交指数、异交倾向与主要传粉媒介等便成为必须关注的问题.研究发现,油茶异花授粉率和坐果率均与昆虫活动规律密切相关,实际生产中油茶低产主要是由传粉昆虫种群和数量匮乏造成的.因此,有效传粉昆虫数量依然是提高油茶产量及坐果率的主要因素.
油茶普遍出现花多果少现象,使得油茶产量及出油率都不理想.该现象主要是因为油茶具有典型的自交不亲和特性,不存在无融合生殖和自花授粉现象,因此必须借助昆虫传粉实现异花授粉才能结实.
油茶是雌雄同株植物,坐果率低主要是受到花粉限制,为了改变这一现状,本试验以‘长林3号’油茶为试材,通过研究其开花物候、花粉活力、柱头可授性、人工授粉及套袋处理、并对其传粉者及其访花行为进行调查调查研究,初步探索油茶的传粉生物学特性和繁育系统,为油茶的增产或选育提供有用的数据和资料.
2 试验材料与仪器
本试验以5 年生的‘长林3 号’油茶为试验材料,在天气晴朗的上午(9∶00−11∶00)选择生长健壮、无病虫害、正常开花的植株并在其大蕾期进行人工授粉.
油茶试验地位于凯里学院继续教育学院同得利山庄,地理位置为26o31′35"N,107o53′11"E,海拔744.90 m,属亚热带湿润季风气候区,年平均气温为13.6−16.2℃,年均日照1000−1400 h,>10℃活动积温4765−5748 ℃,无霜期长达288 d,年相对湿度80%,年平均降雨量1200 mm.
所用的试验仪器主要有电子分析天平(AR224CN,北京赛多利斯仪器系统有限公司),光学显微镜(XSP−1C,北京普析通用仪器有限公司)和连续变倍体式显微镜(JSZ5A、上海博迅实业有限公司)等.
3 试验方法
3.1 物候期及花部特征观察
在试验中进行油茶鳞片松动期、花蕾期、初绽期、盛花期、凋落期、果实膨大期6 个时期的观察,并记录相关数据.花蕾期、盛花期、衰落期、凋谢期、果实膨大期均以第1次出现日期为记录标准.观察日期从2018年9月25日起至2019年4月5日止.
在观察油茶物候期期间,发现油茶上午8∶00−9∶00 开花,下午16∶00 花瓣会闭合.油茶鳞片松动期为9月30日至8日;花蕾期为10月10至12日;初绽期为10月13日至15日;盛开期为10月18日至23日;凋落期为10月24日至29日;果实膨大期为3月15日至3日.
随机标记18 株生长健壮无病虫害的植株且在每株树上选取3 朵当日绽放的花朵,用游标卡尺测定其花瓣(数量、长度、宽度、颜色)、雄蕊(数量、长度)、雌蕊(数量、长度)、柱头、胚珠大小等.
3.2 花粉活力、柱头可授性以及花粉量和胚珠数比率的测定
利用TTC 法和I2−KI 法测定花粉活力[16],联苯胺−过氧化氢法测定柱头可授性[17],通过Cruden R W[18]的标准计算花粉量与胚珠数之比(P/O),并鉴定其繁育系统类型.
3.3 杂交指数估算、传粉者的观察及人工授粉及套袋试验
根据Dafni A[19]的方法标准判断其杂交指数,利用韦维等[20]方法对传粉者进行观察,根据张丽芳等[21]方法进行人工授粉及套袋7 个不同处理试验:CK 为不做任何处理;T1为不去雄直接套袋;T2为同株异花授粉;T3为异株异花授粉;T4为去雄不套袋;T5为去雄套网袋;T6为去雄去雌套袋.(每个处理50朵花且重复3次,共计1050朵花).
3.4 数据统计及分析
本试验数据均采用Microsoft Excel 2010软件录入,后采用SPSS 23.0进行数据分析,花部特征采用描述统计、坐果率采用SNK法(P<0.05)进行方差分析.
4 试验结果与分析
4.1 油茶花部特征观察
‘长林3 号’油茶的花瓣为5 瓣,花白色并呈分离状态,花瓣形态各异,倒卵形,顶端常凹入或二裂,长2.78 cm,宽1.77 cm;花冠直径4.46 cm;雄蕊106 枚,长1.34 cm;花药淡黄色,长0.14 cm;花丝淡黄绿色,外轮花丝与基部连合,长短不一,形成杯状体子房;淡绿色的雌蕊1枚,长1.17 cm,柱头先端三浅裂,当柱头在被未绽放的花瓣包裹时,呈现弧形弯曲,花瓣绽放完全时,柱头顶部变尖、颜色偏绿色,略高于雄蕊群;子房上位,密被白色丝状绒毛;胚珠13 枚,白色,背圆腹扁;蜜腺位于子房外侧花丝内侧,分泌出的花蜜保存于花丝形成的杯状体中;花期10−11月(图1、表1).

表1 油茶花部特征

图1 油茶花部特征(A为油茶花朵,B为胚珠数与胚珠形态)
油茶单花花期为6−7天,花开放前鳞片松动(图2a),花瓣大部分于左右张开.花瓣张开后花药开始开裂,并在内全部裂开.但是如果气温较低(低于17℃)时,花药则在1天内逐步开裂.花瓣张开的第1天花药颜色为淡黄色,产生少量花蜜时,花瓣重新卷起闭合(图2b).第2天早上后花瓣重新张开,产生大量花蜜,并伴有淡淡花香气味,花药从淡黄色变成金黄色,傍晚时花瓣不再卷起(图2c).第3天仍然有很多花蜜,花瓣颜色鲜艳(图2d).第4天上午花瓣颜色鲜艳度略有降低,但不明显,同时还有花蜜下午花瓣开始进一步萎蔫(图2e).第5天花瓣有些萎蔫,颜色鲜艳度更有所降低.第6天花瓣开始脱落,花药颜色变深,出现少部分萎蔫(图2f).第7天花瓣萎蔫,花谢(图2g).

图2 油茶开花进程
4.2 花粉活力与柱头可授性
采用TTC 染色法和碘化钾(I2−KI)染色法进行染色观察,发现花粉的颜色均有明显变化.TTC 染液能够将花粉染成红色或者淡红色的,说明花粉具有活力,无色或者不变色则说明花粉没有活力,采用I2−KI 染液能够将花粉染成深褐色或者褐色的,说明花粉具有活力,无色或者不变色则说明花粉没有活力.因此,有活力的花粉说明花药开裂散粉,无活力的花粉可能没有花药或花药无法开裂散粉.
使用TTC 染色试剂染色时,需1 h 后才能明显地看到染色的花粉,这可能是因为油茶花粉对TTC 染色剂不太敏感;I2−KI 染色法染色10 min 后便能清楚看到染色的花粉.两者进行比较后发现TTC 染色法能直观清晰区分颜色,而I2−KI 染色法因颜色相近,不易分辨.试验结果表明:TTC染色法油茶盛花期花粉活力68.80 %,而I2−KI 染液法油茶盛花期花粉活力则为63.67 %,说明TTC 染色法更适合用于油茶花粉活力的测定.但遇到连续阴天,雄蕊发育不良,出现少量雄蕊瓣化,或者只有花丝没有花药,或者花药无法开裂散粉的现象.
油茶柱头可授期4~5 天,由图3 可知,在开花前1 天至开花后第7 天柱头可授性呈现先增强后减弱的趋势,开花前1天,产生47个气泡,柱头表面有些湿润;开花第1天产生87个气泡,较前1天的气泡增加52.02%,柱头可授性增强,便于昆虫访花时黏附更多花粉;在其花蕾绽放后的第2天,较开花第1天的气泡增加32.03%,柱头可授性达到最大值,柱头表面变得更加柔软湿润;在开花后第3天的柱头上可看到较前1天的气泡个数减少32.03%,柱头表面逐渐失去水分变得干燥;在开花后第4 天,产生70 个气泡,较前1 天减少45.31%,柱头表明失水更为明显,可授性降低;当花蕾绽放后的第5天时花瓣开始萎蔫,产生63个气泡,较前1天减少10.00%,可授性逐渐降低;第6天时花柱开始失水干枯甚至有的花柱开始变成褐色,气泡个数为0,说明失去可授性.结合光学显微镜(10×10)观察,油茶在授粉前,柱头上没有分泌物的产生;授粉后柱头上开始有分泌物覆盖,并黏住花粉,让其成为油茶花粉萌发的基质,当油茶在开花后的第5 天柱头则慢慢失去可授性直至开花后第6 天和第7 天失去可授性.因此,油茶柱头长约5 天的可授期可保证受精的正常进行,提高植物的结实率.
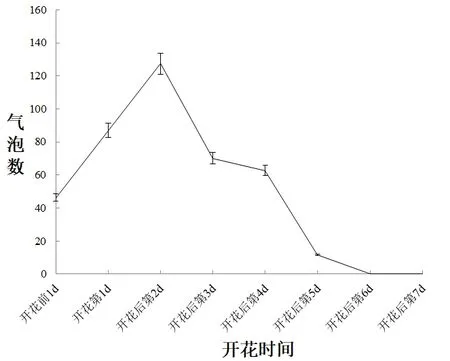
图3 不同开花时期对油茶柱头可授性的影响
4.3 P/O测定及杂交指数估算
测定单花的花粉量,其结果表明单花粉量为33117.00±109.57粒,胚珠数目为13±0.28枚,P/O为2547.46,根据Cruden R W[18]提出的标准将油茶繁育系统鉴定为兼性异交.有效花粉数量是异花授粉成功与否的关键,尽管油茶的花粉活力不高,寿命也不长,但由于花粉量较大(P/O 为2547.46),因此从总体上提高了油茶的生殖成功率.按照Dafni A[19]的方法对油茶的杂交指数进行测量(表1).花冠直径平均值为4.46±0.13 cm.花药开始开裂散粉,随后柱头具有可授性.柱头可授期较花粉寿命更长,但由于花药后期的花粉数量较少、活力降低且花丝干枯松动,少有昆虫访花,因此雌雄在时间上并没有分离(表1).在空间上,刚开放时雄蕊紧贴子房与柱头,随后雄蕊向上且向外伸长,雄蕊(1.34±0.13 cm)略高于雌蕊(1.17±0.02 cm),因此花内雌雄繁殖器官存在一定的空间分离,使得油茶杂交指数达到4(表2).根据Dafni A[19]的划分评判标准,油茶繁育系统为异交型,有较低的自交亲和性,传粉时需要传粉者.
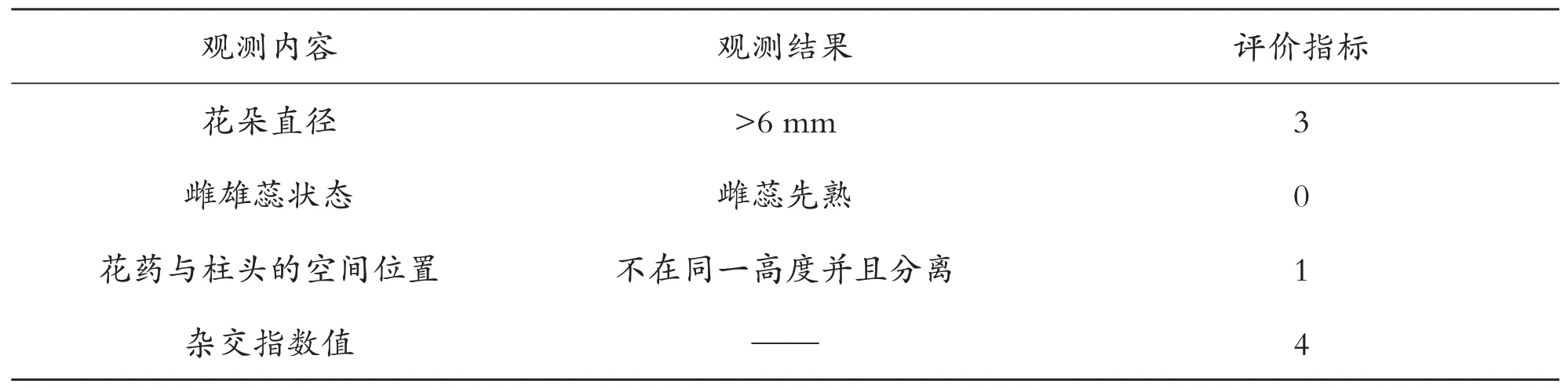
表2 油茶杂交指数(OCI)值
4.4 传粉者的种类
虫媒传粉对花粉流的形成起着决定性作用.试验期间,观察发现,访花昆虫类群包括蜂类(图4A和D),蚁类、鞘翅目的金龟子(图4C)及半翅目的蝽(图4B)等4种访花昆虫.

图4 传粉者种类(A、B、C、D分别表示大分舌蜂、菜虫春、金龟子和油茶地蜂)
大分舌蜂Colletes gigas和油茶地蜂Andrena Comellia访花部位均是花朵,访花时直接落于花中间,连续访花3−4朵,访花时间为9∶00−17∶00.参与油茶花授粉主要是雌蜂,雄蜂一般只采食花蜜不能进行有效传粉[20−23].蜂类一般具有访花速率快、飞行距离远、传粉效率高的特点,易于采集花粉.油茶地蜂比大分舌蜂更耐寒,成虫的活动期从10 月开始至翌年1 月,与油茶的花期高度吻合;油茶地蜂的蜜粉源植物与大分舌蜂一致,也具有寡访花性,也是油茶花期主要传粉昆虫[24−25].金龟子在花朵中间采食雌蕊,易于携带花粉,常常数只长时间聚集于同一朵花上活动,花间迁移较少,因此有利于自花授粉.
4.5 人工授粉及套袋处理
经授粉及套袋处理后,油茶坐果率有所提高(0−53.33%),且坐果率依次为T6 由图5 可见,在p<0.05 标准下,T2、T3、T4、CK 之间均无显著差异;T1、T4、CK 之间均无显著差异;T1、T4、T5之间均无显著差异;T1、T5、T6之间均无显著差异.坐果率T3>T2、>T6.坐果率之间出现差异性,可能是因为处理方式不同(T3为异株异花授粉、T2为同株异花授粉、T6为去雄去雌,无法进行有效授粉),因此,T3、T2与T6极显著差异. 图5 不同处理方式对坐果率的影响 油茶花期从当年10 月至11 月,单花花期为6−7 天,利用TTC 染色法花粉活力为68.8%,高于I2−KI 染色法花粉活力63.67%,单花花粉粒为33117.00±109.57 粒,13 枚胚珠,OIC 为4,P/O值为2547.46,坐果率0−53.33%,其大小为T6 对大部分的植物来说,开花是在绽放之后至花瓣开始凋谢而终止整个开花进程,而油茶开花进程则是开放和闭合交替进行的,有研究工作者将其称之为“二次开合”[22].油茶8∶00−16∶00开花,16∶00后开始闭合花瓣,可节约能量以延长花的寿命,适应访花昆虫少的情况.次日上午9∶00再次绽放,本试验与王湘南[23]研究一致,这与油茶开花习性和当地气候环境有关. 影响植物坐果率的内在因子是花粉活力和柱头可授性,外在因子是外界条件,要完成有效授粉需要有活力的花粉和柱头具有可授性,两者缺一不可[24].这与王湘南[23]研究一致.本试验发现,油茶的花粉活力与柱头可授性达到相对一致性. 植物会通过花朵下垂来提高昆虫访问频率,提高授粉成功率,避免花粉因阳光直射活力降低,延长雄性特征表达时间.试验过程中,发现了4 种传粉者.油茶地蜂是高度特化的传粉者,其选择向更特化的方向发展.油茶地蜂为寡食性的独居型昆虫,但主要取食普通油茶的花粉及花蜜,油茶地蜂在油茶林下的土中,油茶林附近的乔木林缘、空地处,油茶林周边的小路两旁、土坎下及空地草丛的地下筑巢.阴雨或低温天气,林间基本见不到访花传粉者,这与黄墩元[25]研究结果一致,可能油茶在冬季开花,限制了许多传粉者的活动.影响油茶传粉者有效传粉的两个重要因素是气温和光照,有效传粉者的缺乏和较短的柱头可授期是影响油茶坐果率低的主要原因. 据相关研究报道,自花传粉是被子植物进化的一种普遍趋势,是植物在恶劣环境中保证繁育成功的一种适应机制之一[26].在自然条件下,油茶能够结实,但坐果率较低,说明油茶自花授粉能结果.在自然条件下植物会自交的原因来源自身的单花花粉量、传粉者数量以及自花花粉和异花花粉在获得受精机会上的竞争能力有关[27]. 传统观念认为油茶是异花授粉植物,且自花不孕[28].本试验与高超等[28]研究不一致.廖婷等[29]以‘湘林’系列为试验材料,通过自花授粉研究其自交亲和性,结果发现‘湘林’系列的油茶存在较低的自交亲和性.本试验不去雄直接套袋,坐果率达20.00%,表明‘长林3号’油茶具有自花自交的能力,但自交坐果率较低.国家油茶科技攻关组曾在无性系间首次发现自花可孕植株,而且自花授粉率还很高[30],故‘长林3号’油茶自交不亲和性有待进一步证实.
5 结论与讨论
5.1 结论
5.2 讨论
猜你喜欢
安徽农业科学(2022年18期)2022-10-13今日农业(2022年15期)2022-09-20新疆农垦科技(2022年1期)2022-03-10科学之友(2021年12期)2021-12-23中国果业信息(2021年6期)2021-12-02新疆大学学报(自然科学版)(中英文)(2020年2期)2020-07-25大科技·百科新说(2018年3期)2018-04-11商业文化(2017年26期)2017-03-20山西果树(2014年3期)2014-07-15
