
寒武纪生命大爆发与地球生态系统起源演化
2021-12-27 09:24:28张志飞胡亚洲陈飞扬张志亮陈延龙任心宜姚金龙李国祥郭俊锋
西北大学学报(自然科学版) 2021年6期
张志飞,刘 璠,梁 悦,胡亚洲,陈飞扬,张志亮,陈延龙,任心宜,姚金龙,李国祥,郭俊锋,华 洪
(1.西北大学 地质学系/大陆动力学国家重点实验室 陕西省早期生命与环境重点实验室,陕西 西安 710069;2.中国科学院 南京地质与古生物研究所,江苏 南京 210008;3.长安大学 地球科学与资源学院,陕西 西安 710054)
地球是全人类和所有已知生命赖以生存的唯一家园,起源于距今46亿年前太阳系的原始星云,惨遭陨石轰击、放射衰变、原始重力收缩等过程达到熔融状态,并在自转和重力作用下形成圈层结构——地核、地幔、地壳。最原始的地壳约在40亿年前出现,这一变化将地球演化历史分为地壳出现之前的天文时期和之后的地质时期 (见图1)。作为地球上目前最高等的智慧生物——人类,对大陆起源和生命演化这一科学问题自古以来都十分关注[1-4]。关于生命起源,科学家们曾提出了多种猜想,海底热泉、地下湖泊,甚至地壳深部裂缝起源等等[1,4],而保存在岩石中大量的古生物化石和生物地质记录无疑成为地史演化和生命起源的最直接证据[5-8]。

图1 地质年代、大陆演化与重要地质阶段划分,以及本文提出的地球生态系统三大演化阶段,主要讨论了地史生物群落演替、生态环境和生态界面与生态类型及构成的不同演化阶段Fig.1 Geologic Time Scale with geological stages of continental evolution and the coupled composition and divisions of Earth′s ecosystem in to the Three Macro-Evolutionary Phases, proposed herein.The Three Macro-evolutioanry phases are comprised of the Preliminary, Primary and Modern Evolutionary Ecosystems (PREE, PEE and MEE), based on the geobiological data from biological communities (autotrophy/producers and heterotrophy/consumers) and their evolution, ecological environments and ecological interfaces to sustain marine communities composed most of animals and microbial algae, or to maintain terrestrial communities most encompassing plants and animals and microbes
诚然,随着地球大陆和生命的出现[7,9-12],由有机生命与无机环境构成的地球生态系统开始了一系列不可逆转的演变。有机界进化宏观上无疑是有序性,随着地球历史的演化,地球生命的发展经历了从非生命到简单生命[7-8],从原核到真核,从单细胞到多细胞,从无组织器官到有组织、有器官,从水生到陆生等系列进化过程[13]。在这个过程中,随着生物的进化,生物本身组织化、器官化程度的提高和多样性的增长,生态系统也自然趋向复杂化和有序化。同时,生态系统本身是开放的,为了自身的稳定,需要不断输入能量。能量和信息也自然从组织水平低的系统向组织水平高的系统流动。这一演化规律也为地质年代与年代地层划分提供了重要的化石依据。从老到新,地质学家一般将地层划分为太古宇、元古宇和显生宇,对应的地质年代分别为太古宙、元古宙和显生宙(见图1)。显生宙全球地质年代和年代地层的划分和对比,也主要根据古生物的演化阶段和不同类群分类阶元的出现为依据(见表1)。

表1 地质年代和年代地层单位的划分与生物分类阶元和演化的关系
生命最早起始于天文时期之后的太古宙,是生命演化的初级阶段,以最原始的生命形式——原核生物为代表[6-7,14],包括具光合作用的蓝细菌(又称蓝藻)和厌氧的古细菌,在营养方式上应多属于自养生物类型。 在元古宙,原核生物向真核生物演化,在距今约27至25亿年前,地球氧气含量明显增加,为地球真核生物出现提供了关键条件。最早的真核生物的分子化石证据(甾烷)发现于澳大利亚北部沉积岩中(约27~25亿年前)[15],保存形态的最早化石证据发现自加拿大19亿年前燧石层内[16]。当前,我国发现的最早真核生物化石证据来自北方串岭沟组(18~17亿年前)和山西永济汝阳群(15~12亿年前)等地层内的大型藻类和疑源类化石[17-19]。
到了元古宙晚期,真核生物的多样性明显提高,多个保存有真核生物的化石库陆续被发现并报道,真核生物碳质压膜化石在世界多个大陆板块发现[18,20-21]。从新元古代开始,多细胞后生动物发生辐射,大型带刺疑源类化石形态发生较大分异,生物的营养方式已出现自养生物和异养生物两种类型,这些异养型生物主要以埃迪卡拉生物为代表。我国华南也相继发现了大量的埃迪卡拉纪宏体生物群,如蓝田生物群、石板滩生物群和高家山生物群等等[22-24],此类生物主要以表栖固着生物为主,生态虽然单调,但高度上显示出一定的生态分层[25]。成层出现的遗迹化石为埃迪卡拉纪晚期生态系统的复杂性提供了有力证据[23,26]。这些宏体化石或者印痕化石虽有学者认为可以与现生的真后生动物类型相类比[27-28],但具骨骼、长眼睛、有附肢、能滤食、会运动的两侧对称动物却明显突然大量出现在寒武系底部,指示了隐生宙—显生宙地球生物和环境的重大转变[29-32]。这样,地球在经历了长达35亿年的前寒武纪原始积累,从菌藻席主导的原始生产者海洋生态系统开始向动物消费驱动的显生宙海洋生态系统转变[33-37]。在新元古代埃迪卡拉纪末期—寒武纪早期(第二世)短短4千万年的时间里,地球上突发性地出现了包括脊椎动物在内的几乎所有现生动物的早期化石代表[29-30],依次诞生了基础动物、原口动物和后口动物亚界,导致地球动物演化树的快速成型,这就是寒武纪大爆发(Cambrian Explosion)[29,31,36]。从生态系统生物群落和非生物因素两方面具体来讲,寒武纪大爆发生态上可以进一步理解为地球由前寒武纪微生物席为主体的原始进化生态系统(2极食物链2类生活界面:生产者和分解者/陆表和海水)向以动物为消费者的现代海洋动物生态系统(3极食物链2类生活界面:生产者、消费者和分解者/陆表和海水)演化。地球早期海洋中主动取食者——动物的出现,代表着生态系统进入了生产者、消费者和分解者组成的3层次食物链结构框架的初级进化生态系统阶段(见图1)。寒武纪生物高级分类阶元类型虽然很多,主要包括最早的脊椎动物和现代海洋中广泛分布的软体动物、环节动物、腕足动物等分类多样的底栖动物,但缺乏现生类型的中高级消费者。在初级生产者方面,现代初级生产力极强的植物界(专指具光合作用的陆地多细胞真核生物)远未出现,初级生产者仍以自养型菌藻类为主。与现代生态系统相比,最大的特点是所有的生物都离不开水,只能在海洋水体中生活。三层次食物链系统虽开始发育,生态结构也趋于复杂,但缺乏陆地生态因素,整个地球生态系统仍处于以海洋为底质,以海水为媒体/介质的单一海洋生态系统,因此属于地球生态系统演化的初级阶段。生物的生活环境也只包括液相(海水)和固相(海底岩石)两种界面。相对陆地生态系统的3相界面(液相、固相和气相)(见图1),在陆地和低空的气相界面还没有出现生物,因此生态环境相对局限和简单。
寒武纪时期,地球海洋处于陆表浅海环境,海洋底质由微生物席型底质向混合型海洋底质演化过渡,海底多泥多沙,缺乏后生动物建造,海洋硬底质环境尚未形成。海洋原始生产力仍以菌藻类为主,初级消费者以底栖无脊椎动物为主,包括固着型、游移型、游泳型和少量浅表内栖型等,已开始形成了多层次的海洋动物群落结构。但是,到了奥陶纪和志留纪,生态分层加速、多样性加快,动物低级分类阶元(科以下)进一步丰富(见图1),显现了地球最早初级进化生态系统的丰富和成熟化过程。泥盆纪是古生代生态系统重要的转折时期,古特提斯构造域开始形成,地球生态系统由早期单一的海洋生态系统初步演化出海、陆不同的生态系统,部分生物逐渐摆脱水域的影响,开始在陆地表面呼吸、生殖和运动,形成了液相、固相和气相三种界面的多样化生态系统(见图1)。从奥陶纪、志留纪到早泥盆,古无脊椎动物多样性空前繁盛,底栖生态、浮游生态、礁体生态和大型游泳型生态已高度发达。底栖群落以腕足动物为代表,礁体开始繁盛、以珊瑚、层孔虫和苔藓虫为主,浮游生态以笔石动物为代表,游泳型无脊椎动物以角石类头足动物为代表[38],脊椎动物开始出现以大量的有颌鱼类为代表,大型游泳生态出现并空前发育[39]。这样,现代海洋动物生态类型已全部出现并繁盛,代表着地球现代海洋生态系统的形成(见图1)。之后,地史时期海洋生物类型虽不断变化,但在生态系统演化中应只属于群落演替范畴(详见下文)。
本文主要以生态系统中的生物因素为主线,结合地质演化和阶段性(见图1),以寒武纪大爆发期间动物的生活方式(营养方式、运动方式和居住方式)、群落生物相互关系和生态工程建造(Ecological engineering)为焦点,重点探讨5.3亿年前后海洋动物生态系统的形成和复杂化过程。以期探讨地球最早动物生态系统的形成,生物圈和岩石圈相互影响、促进和发展,以期理解地球生态系统的形成发展和宜居性演化过程和阶段。
1 生态系统的划分及其主要地史演化阶段
1.1 生态系统的概念和内涵
什么是生态系统(ecosystem)?要了解生态系统的发展,首先必须掌握生态系统的组成。它是一定时间和空间范围内由生物群落与环境之间不断进行物质循环和能量流动而形成的统一整体。基于此,生态系统的组成主要包括生物本身和非生物环境两个部分(见图2)。非生物环境可以理解为生命的支持系统,包括大陆和海洋环境在内的不同生态因素(也称为生态因子ecological factors),根据地球科学的关注内容,本文建议可分为生长底质(泥岩、砂岩、台地、土壤等)、环境成分(如光、氧气、温度、深度以及海水的咸度、盐度和酸碱度PH值等等),生态介质(海水、空气、底质和土壤等)和代谢因子(C、N、P、S、 Ca、 无机盐等)四大方面。生态系统中的生物有机体包括所有的生物群落,根据它们获得营养和能量的方式以及它们在能量循环中发挥的作用,可以分为生产者、分解者和消费者(见图2)。
生产者包括所有的自养型生物,有光能自养型和化能自养型的细菌和深海中的小型生物。 消费者包括所有的动物和腐生、寄生生活的菌类或分解者,所以有大型消费者和微型消费者之分。分解者又称为“小型消费者”,也属于异养型生物,如细菌、真菌和罅隙型动物,它们将复杂的动植物尸体或遗体分解为简单化合物或者无机物质,可以再被生产者或者消费者利用。在生态系统中,生物与环境相互影响、相互制约,并在一定时期内处于相对稳定的平衡状态(见图2)。生态系统的范围可大可小,相互交错,生物圈当然是地球上最大的生态系统,也是地球上所有生态系统的综合整体,通常包括地球海平面之上和之下1万米的空间圈层,包括大气圈底部所有飞鸟、昆虫和细菌可以触及的范围,涉及一切生物立足的岩石圈表面以及全部有生命赋存的水圈。根据生物生存环境不同,地球生态系统可被分为水生生态系统(淡水和海洋)和陆地生态系统。根据研究的需要,水生生态系统进一步可以细分为淡水生态和海洋生态,而陆地生态系统可以细分为森林生态系统和草原生态系统等,这都是现代生态系统的基本概念和分类。
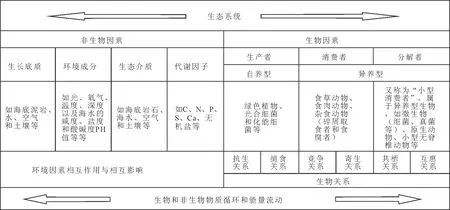
图2 由生物因素和非生物因素构成的生态系统和物质循环Fig.2 The composition and material cycles of an ecosystem consisting of inorganic and organic factors
1.2 地球生物分类和基本框架
生命是能够自我繁殖、自我复制的有机体,生物多样性是生命最尤为突出的特征。对于纷繁复杂的生物界,我们可以根据其大小形态和运动方式进行分类,有微观生命和肉眼可见的生命世界。随着当代生命科学的发展,尤其是分子生物学、基因组学等学科的发展,极大改变了人们对生物的传统分类和系统认识。根据Knoll[40]的核糖体RNA分析研究,地球生物可以分为细菌(bacteria)、古细菌(archaea)和真核生物(eucarya)三大生命超界/域 (见图3A)。然而,根据生物的结构形态和营养方式,生物五界分类系统(原核生物界、原生生物界、真菌界、植物界和动物界)通常广为多数学者所接受 (见图3B)。需要说明的是,古细菌和真细菌都是原核生物,但两者明显不同。古细菌往往生活在厌氧的沼泽、盐湖和酸性温泉等极端环境中,而真细菌在现实生活中分布十分广泛,其细胞结构往往具有特化的鞭毛或纤毛,以利于适应潮湿环境或者黏附在一些特殊有机体表面。蓝细菌又称为蓝藻,也属于一类具有光合作用的原核生物,常常在现实生活中大量繁殖在富营养化的水体中,从而形成水华(algal bloom)。因此,蓝藻不是藻,是一种特殊的具光合作用的原核生物(又称蓝细菌),它在叠层石地质记录的形成中发挥着十分重要的作用,是叠层石形成的生物贡献者(见图4)。与蓝细菌(蓝藻)不同,藻类是一类具有纤维素细胞壁的植物状的真核原生生物,有浮游生活的,也有底栖固着的藻类。我们食用的海带个体可以达到百米以上,属于藻类,是一种底栖生活的原生生物(见图3B)。藻类虽然有许多与植物相同的特点,但与植物明显不同。植物是适应于陆地生活的,是具有光合作用能力的多细胞真核生物,除苔藓植物外,都具有根茎叶的分化(见图3B),最早出现在志留纪末—泥盆纪早期[41-42]。但藻类不同,虽呈植物状,但只能在水中生活,最早可以追溯到18~20亿年前的元古宙早期。因此,谈及地球早期生命,都是菌藻类和早期动物相关的海洋生态事件[43],与早期植物无关。但植物的起源与陆地生态系统的起源与演化息息相关,是现代进化生态系统的建设者和初级生产力标志。

A 基于分子生物学的三大超界/域生物系统发生树(细菌、古细菌和真核生物),修改自文献[40];B 根据生物的形态结构和营养方式划分的地球生物五界系统分类方案,包括原核生物界、原生生物界、真菌界、植物界和动物界图3 地球生物三分和五分系统分类方案Fig.3 The taxonomic classification of organisms on Earth into three or five kingdoms
真菌的细胞内不含有光合色素,也无质体,在生活中只能寄生或者腐生在其他生物上,是典型的异养生物。在现实生活中,真菌和藻类(蓝藻)可以共生在一起,形成共生体,称为地衣。在极端环境中,藻类和蓝细菌通过光合作用为真菌提供养料,真菌可为蓝藻或者藻类提供生活必须的水分。在地质历史中,地衣可能出现在前寒武纪末期[44]。

A~H 南非太古宇化石;A~C,野外考察照片,图示Ventersdorp超群Ongers River组内部27亿年前的雨痕;A 雨痕化石碎片斜插在沙屑中;B 放大显示片状保存的雨痕碎片化石;C 不同化石的细节放大;D~E 条带状磁铁矿BIF层样品光面光学照片和微区X-射线荧光光谱仪(Micro-XRF)铁元素分析照片;F~H Pongola超群(约29~31亿年)和Transvaal超群叠层石化石(约25亿年);F 纵向剖面照片;G~H 横断面光学照片;I 陕南汉中梁山大南沟剖面灯影组顶部叠层石野外照片;J~L 河南宜阳地区洛峪口组顶部穹窿状大型叠层石礁体和纹层状结构图4 前寒武纪雨痕和叠层石及其块状礁体Fig.4 Field photos of raindrops and stromatolites from the Precambrian of South Africa and China
动物不同于植物和真菌类等其他生物,通常都具有运动能力,异养生活,营体内消化。虽然有学者或文献将单细胞原生动物也包括在动物内,但根据生物五界分类系统(原核生物界、原生生物界、真菌界、植物界和动物界),动物界通常只包括原生动物以外的所有多细胞真核动物门类(见图3B)。原生动物是单细胞真核生物,由单个细胞或多个细胞群构成。鞭毛虫纲是原生动物中最原始的类群,其中地质上最重要的原生动物化石有放射虫和有孔虫等[45]。后生动物起源于原始的单细胞真核生物祖先,经历了简单到复杂,低等到高等的系统发育过程,根据胚胎的发育和体腔的有无,动物进一步发育并可以分为二胚层、三胚层动物,或无体腔、假体腔和真体腔动物等(见图5)。然而,根据最新的分子系统学和基因组学系统分析,多细胞后生动物包括辐射对称的基础动物和我们熟知的两侧动物。两侧动物主要包括包括脊椎动物在内的后口动物,以及包括冠轮动物和蜕皮动物两大分支的原口动物(见图6)。这里只介绍基本分类框架,详细的形态、解剖等门类特点和鉴定特征不再赘述。因此,总体而言,多细胞动物可以四分为基础动物、后口动物、原口动物蜕皮类和冠轮类,三大亚界,四大超门(见图6),其化石最早记录显示多为“草根”型基础动物和冠轮类超门的不同类型(见图6)。

图5 基于形态特征建立的传统动物界系统分类方案,如对称性和内部体腔的有无等特征Fig.5 Traditional animal phylogeny based on morphological characters, such as symmetry and presence/absence of internal body cavities etc

图6 基于分子生物学的现代动物系统分类框架,图示寒武纪大爆发导致基础动物、原口动物和后口动物三大亚界依次成型。基础动物毫无疑问可以追溯到埃迪卡拉纪晚期,在寒武纪纽芬兰世幸运期其骨骼类祖先大量辐射;冠轮类原口动物大量出现在寒武纪幸运期,而蜕皮类原口动物和后口动物化石记录出现较晚,主要发现于寒武系第三阶Fig.6 Modern animal phylogenetic framework based on molecular data, and the schematic pattern of Cambrian explosion of metazoans, with explosive radiation of basal metazoans, protostomes and deuterostomes in successions. The basal metazoans (1) can be traced no doubt back to the late Ediacaran with their explosive radiation of their skeletonized ancestors in the earliest Cambrian (Terreneuvian, Fortunian Stage); the mass appearances of lophotrochozoans (2) in Fortunian were essentially earlier than the occurrence of most ecdysozoans protostomes and deuterostomes (3) in Cambrian Stage 3
1.3 地球生态系统的划分和阶段
如上所述,生态系统是生物群落和环境构成的整体。因此,讨论生态系统的演化必须讨论地球生物群落和环境两个方面的演化(演替)。地史上,尽管生物群落和主要生物类群明确具有鲜明的阶段性和地史性特点,但要证明详细的生物演替也伴随着与之相吻合的环境阶段性却不是十分容易的事情。纵观地球大陆演化与重大生命演替和重要功能生物类群起源,本文提出,地球生态系统(宏观上即生物圈)根据大陆演化与生物群落的演变建议粗分为原始进化生态系统、初级进化生态系统和现代进化生态系统三大演化阶段(见图1)。原始进化生态系统以陆核形成、成熟,早期超大陆聚散,以及早期板块构造启动为地质背景[46-52],生物类群主要以菌藻类为生产者(见图4),以细菌微生物为分解者的2极食物链结构,生活底质仅以海洋底质(岩石)和沉积物为界面,生活介质仅以海水为介质的2相(液相和固相)环境,时间包括太古宙和元古宙为主的大部分地史时间,代表着以微生物(席)为主体的微生物生态系统的形成和演化阶段。在地球原始进化生态系统中,叠层石通过蓝藻的生长不断捕获和固结海底沉积物(见图7A),海底有机质被海洋微生物分解降解后进入岩石圈和地球内部,代表着生物圈与岩石圈物质初步循环的开始和发育(见图1)。

A 菌藻类初级生产者诱导的地球原始进化生态系统,图示华北大约10亿年前南芬组发现的多细胞藻类化石群落[18];B 埃迪卡拉纪晚期—早古生代以无脊椎动物消费者驱动的地球初级进化生态系统,动物群落由底栖固着型、游移型到高级消费者游泳型的演化过程,代表地球海洋生态系统的形成 [181];C 海陆空交互的现代进化生态系统,陆地植物代表地球最为高效的初级生产力,大型捕食者和高级消费者海陆分布,生态位特化分异巨大(2017年摄于南非好望角)图7 图示本文提出的地球三大演化生态系统:原始进化生态系统、初级进化生态系统和现代进化生态系统Fig.7 The proposed three-divisions of Earth′s ecosystem into Preliminary, Primary and Modern Evolutionary Ecosystems (PREE, PEE and MEE) as illustrated
初级进化生态系统以现代板块构造起始[53-55],冈瓦纳大陆聚合、潘基亚大陆形成的早期阶段为地质背景[52,56-58],以真后生动物(两侧动物)的出现为标志,代表着生产者、消费者和分解者3极食物链结构的形成。期间原始进化生态系统中出现的生产者与分解者的主要组成不变,生物生活底质也仅以海洋岩石和沉积物为界面,介质仍以海水为介质的2相(液相和固相)环境。但是,因为动物的出现导致关键生态功能群——消费者诞生,使有机和无机物质循环加速,营养方式由原始进化生态系统被动方式(光合作用和吸收)演化出动物积极、主动的摄食方式。海底生物群落生态分层不断提高,生物群落复杂度明显增强,出现了捕食、底栖固着、共栖、包壳和寄生等复杂生态现象的生态群落结构。生物组成上主要以海洋无脊椎动物为主,出现底栖、游移、浮游和游泳等现代海洋生态成分(见图7B,8),代表着现代海洋动物生态类型的形成。时间上主要以埃迪卡拉纪末和早古生代为主体,代表复杂现代海洋生态系统的形成。期间礁体动物出现,生物礁繁盛,刻蚀(污损)生物开始大量增殖,海洋动物生态系统工程加速(见图7B,9),包壳和污损动物的刻蚀和相互作用导致海底岩石生物风化速率急速加快,生物圈和岩石圈物质循环迅速加强,地球海洋生态系统循环和能量交换速度不断提高(见图7B)。
现代进化生态系统以古新特提斯洋的演化和潘基亚大陆形成和解体为地质背景,包括该大陆形成晚期和解体的不同地质阶段[52,59-60],以陆地动植物的出现、生物登陆和发展为标志。期间陆地生态系统逐步形成、发展和繁盛,3极食物链结构进一步丰富完善。初级生产者以植物界和菌藻类,多样性和分布范围更加宽泛,部分生物已开始跨越陆地和海洋两种生态系统(见图7C),生产者与分解者多样性大大提高,生物生活底质以大陆、海洋和大气为界面(液相、气相和固相),以海水、大陆和空气为交流介质,部分动物海陆交替生活。不同地史时期不同类型的动物形成“上山下海”生态现象,生态群落和营养结构复杂多变,生态类型繁多,高级生产者和消费者空前繁盛,海陆空生态系统交错复杂,相互影响(见图7C)。时间上横跨晚古生代、中生代和新生代,直至现在,代表海陆交互的复杂地球现代进化生态系统的形成、发展、演替和繁盛。生物圈、岩石圈、水圈和大气圈以生物为纽带,物质交流和能量流动速度加快,成为现代地球生态系统的最大特征。
2 地球初级生产者起源演化与原始进化生态系统形成
纷繁的生命是地球的灵魂,是地球区别于其他星球的主要特征之一。迄今,还没有确凿的证据足以证明除地球之外其他星球还有生命存在[61]。地球因为生命的出现和繁盛,才泽育了生态系统的演化和发展。有地质证据表明,地球的生命是从无到有、从简单到复杂演化而来。其中原核生物起源、真核生物起源和多细胞后生动物起源(寒武纪大爆发)是地球早期生命演化的三大里程碑事件,而后生动物(动物门类)的起源是最为宏伟的生命创生事件,诞生了地球上几乎所有的现生动物门类的早期代表,自此地球才真正地踏上了蓝色星球之旅。海洋动物生态系统出现之后,逐步建立、发育并繁盛,最终部分动植物率先登陆,摆动了陆地重力束缚、海水限制和呼吸困难,逐步开始在陆地上繁殖和生活,陆地生态系统最终出现,从而形成了海、陆、空生态系统并存,相互作用、相互影响的现代生态系统 (见图1,7C)。
地球最古老的大陆岩石可以追溯到44亿年前[1,62]。最新研究表明,地球上出现最早的生命记录(化石)大约38亿年或更早,是由最原始的叠层石微生物形成的[6,14]。叠层石代表最原始的生命形式,是地球初级生产力的开始,一切生命和营养有机质的开始,从太古宙早期一直延续到现代。澳大利亚鲨鱼湾(Shark Bay)大量繁盛的大堡礁就是很好的例证。早期的叠层石形态有对称和不对称之分,形态多样(柱状、穹状、褶皱状、圆锥状)(见图10),沉积于浅海-近地表受热液活动影响的环境[63]。但是,最近Nutman等人[64]重新研究了来自格陵兰Isua Supracrustal带内部约37~38亿年前的叠层石标本,最终认为其成因于浅水环境。 北美和南非地区也发现有大量的叠层石记录[65-66]。 南非的太古宇Pongola超群Chobeni组(距今29~31亿年)和Ventersdrop超群Omdraaivlei组(距今27.4亿年)内部保存有大量叠层石化石,是该地区较古老化石记录。大量繁盛的叠层石可形成穹窿状巨大的礁体,成为最早的地球生态工程/建造(ecological engineering)(见图4)。这些大量繁盛的微生物无疑成为改造地球表面的最早生命类型。在环境方面,南非Omdraaivlei地区还保存着地球上最早的雨痕化石(见图4A~C),这些27.4亿年前的雨痕被保存在Ventersdorp超群Ongers River组凝灰岩内。有学者根据这些雨痕的分布和密度预测大约27亿年前地表大气密度比现今大气密度的一半还低[67],雨痕的存在也说明地球当时已经形成了稳定大气圈[68](见图4A~C),大陆地壳已形成、生长、克拉通化、开始稳定并成熟,地史上出现了哥伦比亚超大陆和罗迪尼亚超大陆等不同阶段,属于地质演化的前加里东时期(见图1)。但是,前寒武纪太古宙沉积记录中的化石主要是微生物形式,因此前寒武纪的生态研究无疑主要属于微生物古生态(microbial paleoecology)范畴[33]。其中叠层石(stromatolites)和微生物诱导的沉积结构(MISS)成为前寒武纪古生物学、岩石学和沉积学关注的主要对象[69]。这些微生物结构不仅在碳酸盐岩环境中大量繁殖,还可以大量出现在碎屑岩岩相环境中(见图10)。这些化石证据表明,早期的生命席状生活在海底,进而形成微生物碳酸盐岩。其中的有机质固结、吸附海底沉积物,最后腐烂进而推进地球早期的有机和无机化学循环[6]。
叠层石-微生物席支撑的原生生物海洋生态系统持续到新元古代晚期(6.35亿年前)[72],地球环境发生巨大变化,全球变暖,氧气含量增加,真核多细胞复杂生物大量出现,包括多细胞藻类和大型的多细胞生物或真后生动物(Eumetazoa)。埃迪卡拉时期,虽然仍存在大量的叠层石碳酸盐岩沉积(见图4),但生物遗迹化石明显陡然剧增,形态多样,复杂化增强,海底沉积界面附近生物扰动明显加强[23,26,70]。地球生物群落发生宏观质的改变,生物从单细胞生物演化出多细胞生物类群。代表性的生物就是大型的埃迪卡拉生物群,它们形态多样,报道发现化石类型100余种,分布时间5.7~5.4亿年前[71,73]。目前,全球已发现产地超过30多处,我国最为典型和最具代表性的是湖北三峡的石板滩生物群。埃迪卡拉生物群的出现代表着地球自生命出现以后海洋生态系统发生了重大的群落演替,自此持续30多亿年的微生物席生态群落被宏体真核多细胞生物群落所主导,仍有学者质疑它们到底代表最早的多细胞动物(metazoans)还是多细胞植物(metaphytes),或者兼之,仍存有争议,在此不赘讨论。
3 地球海洋动物起源与初级进化生态系统形成
如上所述,初级生态系统以冈瓦纳大陆聚合[52,56-58],原特提斯海洋形成演化[74]为地质背景。当时大陆碰撞融合,现代板块构造建立、全球构造联动,风化加强[55],海平面降低,大陆风化矿物质元素源源不断汇入海洋,氧气增加,生物栖息环境多样性出现[75-76]。 地球发生里程碑式生物事件,首次出现初级消费者——两侧对称的无脊椎动物,地球开始进入了以多细胞动物(后生动物)和两侧对称动物演化和需求驱动的初级进化生态系统(见图11)。自此,我们的地球才真正踏上了宜居蓝色之旅,动物、微生物、环境协同演化,生物圈、水圈和岩石圈层协同发展。海底沉积物为后生动物提供发展平台,后生动物通过行为和生态系统工程建造影响和改造海洋底质,有机和无机环境相互制约、共同促进和发展,并开始驱动地球宜居性发展。真后生动物尤其是两侧对称动物经过寒武纪大爆发、奥陶纪大辐射,在志留纪末期地球动物海洋生态系统成型,各种海洋生态类型、营养方式的生物(动物)包括大型游泳型动物均已形成[32,77-79,181](见图7B, 12)。生态统计发现,早古生代之后的动物门类和营养方式等再无新变化和新类型增加,泥盆纪之后海洋生态系统的演化因此属于群落演替的范畴(见图11,12)。
在一定的地质环境背景下,生态系统的生物因素就是生物群落构成和结构。与原始进化生态系统的不同,埃迪卡拉—寒武纪界限附近地球首次出现了大量的初级消费者,食物链开始形成三层次营养循环结构,构成了完整的生产者、消费者和分解者闭环循环。期间,埃迪卡拉生物化石代表地球动物的首次辐射,再现地球基础动物的最早成型过程。之后,两侧对称动物快速出现,以寒武系底部的小壳化石及其上的澄江动物群为代表(见图13)。寒武纪早期地球处于陆表浅海环境,海水息息相通,初级生产者分异低,全球环境营养背景相似。但动物一旦出现,不同的生物相互作用,相互影响,就自然形成了最早的动物消费者驱动的生物群落。生物群落之间的相互关系,如捕食、共栖、对抗、寄生、包壳、幼体底质选择等,形成了早期动物之间的复杂生态关系(见图8, 9, 12)。生态关系反映了生态系统的复杂度、生态动力和运行效率。生物群落生态中除了生物本身的多样性,另一个重要的生态指标就是生活方式。生活方式是指生物为适应环境条件而具有的行为和生活习性,包括运动方式和居住类型(合称生态类型)及其摄食营养方式,也就是反映在动物吃、住、行3个方面,构成了生态多样性的重要指标和研究对象(见图8, 12)。
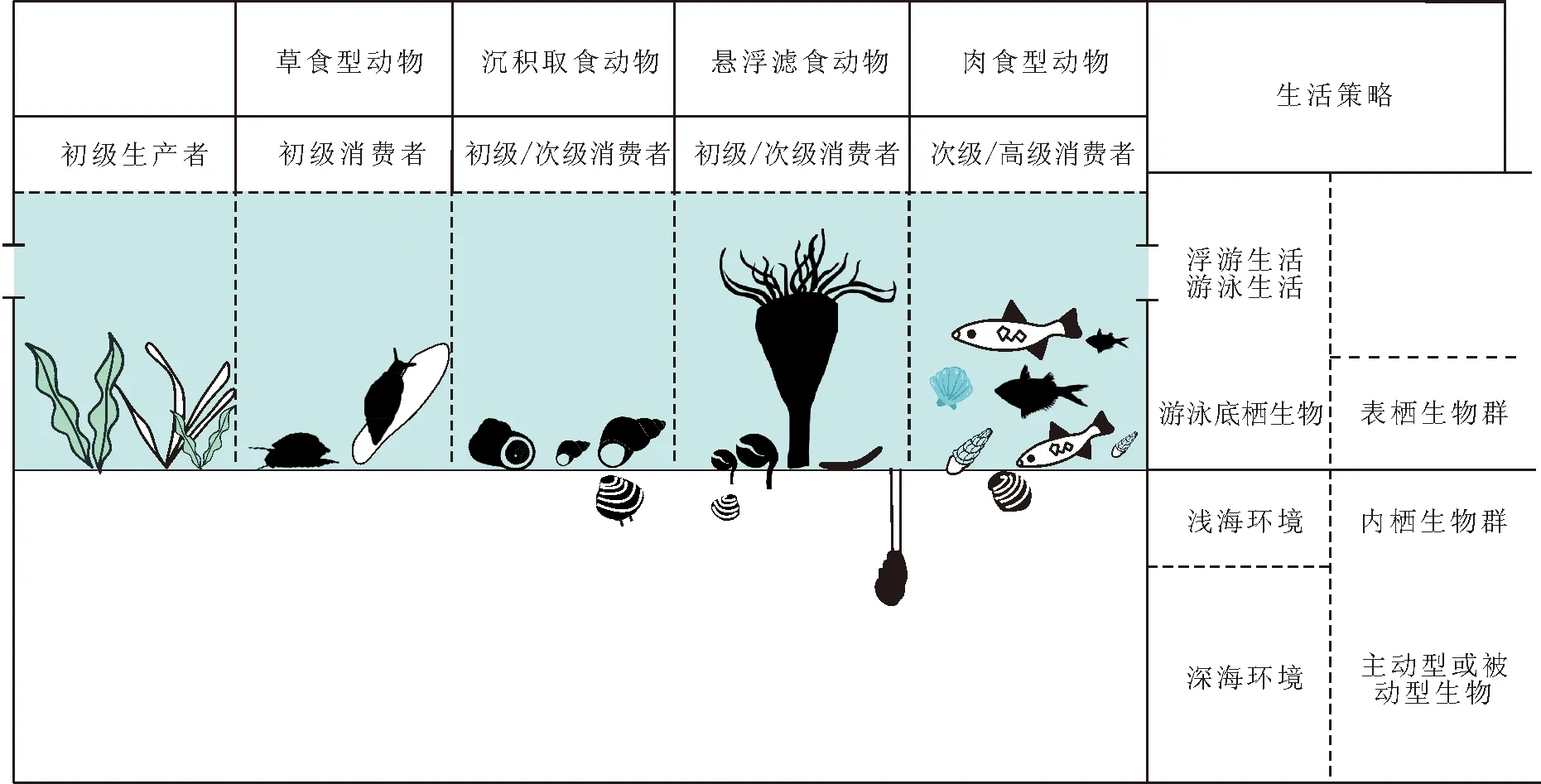
图8 海洋动物生活方式、营养结构和生态位(修改自文献[298])Fig.8 Diversified lifestyles, trophic structures and living niches of marine animals

图9 寒武纪大爆发重要生态创新和生态系统工程、生态多样性变化和复杂化过程Fig.9 Marine ecological engineering and community complexities and successions during the Cambrian explosion of metazoans

A 古元古代叠层石,富藻纹层宽度远小于富屑纹层;B 中元古代叠层石,富藻纹层宽度接近于富屑纹层;C 新元古代叠层石,富藻纹层宽度远大于富屑纹层图10 太古宙、元古宙时期叠层石的多样性及总体形态[33]Fig.10 The diversity and configuration of stromatolites from Archean and Proterozoic[33]

A 寒武纪大爆发时期门类、干群纲类和纲类的多样性辐射模式;B 寒武纪大爆发相关假说:寒武纪信息革命、农垦革命以及寒武纪底质革命图11 寒武纪大爆发模式及假说Fig.11 The radiation pattern and hypotheses on Cambrian explosion of metazoans (Phyla)

图12 早古生代生态多样性演变模式和地球初级进化生态系统的建立Fig. 12 Marine animal ecological diversification during the Early Paleozoic and the establishment of the Primary Evolutionary Ecosystem (PEE) on Earth

图13 早古生代海洋不同动物门类属种首现及多样性变化模式Fig.13 The generic diversities and evolutionary patterns of different animal phyla during the Early Paleozoic
为详细了解地球多细胞动物(两侧动物)的辐射和生态演化,不同的学者提出了不同的假说,力图从不同的视角解析地球动物生态系统的形成过程和环境制约(见图11B)。前寒武纪微生物席和藻类主导的表栖生态系统一直持续到寒武纪早中期[80],但不同于前寒武纪,寒武纪移动生物(vagile epibenthos)增加,海洋沉积物不断受到动物的扰动和影响,大量动物在海底表面开始爬行,甚至钻孔或者形成潜穴,海底表面被动物作用、翻腾扰乱。德国遗迹学者Seilacher Dolf将这种沉积物和动物关系理解为农垦革命(agronomic revolution, AR)(见图11,14)[72,81],形如农业生产中的翻耕和犁地,导致“田地”疏松,表面有机物和沉积物/土壤充分混合,促进“作物生长和农业生产”。该假说主要针对前寒武纪的微生物席底质的打破和显生宙混合底质的形成提出的,适合于硅质碎屑沉积背景的生物群落。 农垦革命不仅直接加速了海洋有机和无机的相互作用和循环,而且直接影响寒武纪海底生活的动物形态、分布和种类,通过对海底表面的作用和影响,一种动物通过“农垦革命”开始影响和作用另一种动物的生活和生长,形成生态系统工程(ecosystem engineering)[82-83]。与农垦革命有异曲同工之妙的理论是寒武纪底质革命(Cambrian substrate revolution,CSR)[84-85](见图11,13)。随着前寒武纪微生物席生态类型生物的迅速减少直至消失,在软的沉积物表面自由或者固着生活的动物出现,沉积物中潜穴、内栖或者罅隙生活的生物也开始繁殖。这种生物活动和生活反复影响和扰动海底表面,使海洋原本安静的沉积过程中断。因此,寒武纪底质革命导致前寒武纪只受物理和微生物过程影响的海洋环境和沉积物,在寒武纪不仅有物理和微生物的持续作用,还主要受到后生动物的作用和改造,通过生物扰动(bioturbation)致使海洋环境的物理作用和微生物过程发生间断和影响,从而最终导致沉积过程的物理过程和后生动物的扰动作用成为环境沉积结构(sedimentary fabric)的主控因素[84,86-87](见图14)。寒武纪底质革命强调的是表栖动物对混合底质的反馈和主动性改造或适应改变,重点观察底质改变对早期底栖后生动物进化和生态的影响。该假说也包含后生动物通过各种行为和生活形成生态工程和生物建造,包括潜穴、钻孔、介壳层(堆)和各种壳体礁和生物礁体等,从而导致海洋底质的硬化或者固化,部分可理解为农垦革命的影响和结果,但更多的是生物和底质环境的相互作用和影响。寒武纪信息革命(Cambrian information revolution,CIR)[88]强调后生动物创新性复杂取食策略的出现及其对生态空间利用效率的提高,表明了不同类型动物的起源和对不同环境的适应性策略。宏观感觉器官的出现和运动能力的加强,使动物对环境的选择性提高,不同的生物伴随着不同的生态和生活方式,不同生活方式的动物继而占领不同的生态位,加速提高生态空间和生态系统的利用和效率,还有生态系统复杂度的增加(见图9, 12~14),这是寒武纪信息革命的内涵。总体而言,三种假说从不同的角度解释了显生宙后生动物(尤其是两侧动物)的出现、辐射和生态扩张。三种假说有一定的统一性和逻辑性。寒武纪信息革命(CIR)解释了不同类型的有组织、有器官动物的出现,运动发生,生态功能利用加强,进而导致农垦革命(AR)发生。农垦革命(AR)使得海洋底质不稳定性发生和异质性增强,致使前寒武纪微生物席底质衰竭,进而促使寒武纪底质演化(CSR),从而完美解释了有机生物对无机环境的影响和作用(见图14)。

图14 寒武纪底质革命和后生动物主导的海底环境演变 (引自文献[84])Fig.14 The Cambrian Substrate Revolution hypothesis and sedimentary fabric evolution from a combination of physical and microbial processes to a combination of physical and metazoan processes (modified from [84])
下面,本文倾力生物多样性和生态群落形成及复杂性方面对寒武纪大爆发动物系统辐射(地球动物树成型)和生态扩张,尤其是早古生代动物的多样性、生态类型和摄食方式的起源和演化进行探讨,力图从群落生态的角度理解地球初级进化生态系统的起源、发展和形成过程。
3.1 地球动物树成型和寒武纪大爆发
3.1.1 寒武纪大爆发地球动物辐射和演化 如上所述,多细胞动物又称后生动物,根据体腔的有无、个体发育、组织器官的形成可以分为三大亚界、四大超门,即基础动物、后口动物、原口动物蜕皮类和冠轮类(见图6)。
根据最新的分子系统和基因组系统学研究,基础动物系统学存在的主要问题是海绵动物与栉水母动物的姊妹之争,也就是海绵动物相对栉水母类最为原始还是反之[89-94]?因此,化石记录和关键化石的解释就成了问题解决的关键,自然引起学者的极大关注,系列研究都发表在高影响力的期刊杂志上[95-97]。生态上,这些基础动物大量繁殖为两侧对称动物的繁殖提供了大量的养料和食物,并与冠轮动物生态上一起成为海洋生物礁等生态工程的建造者,化石上常见的有海绵动物、刺胞动物珊瑚类(四射珊瑚)和各种锥石类化石[98-100]。
后口动物在动物系统分支中相对稳定,在国际学者的研究中有很好的共识[89-90,101],主要包括棘皮动物、半索动物、头索动物、尾索动物和脊椎动物 (见图6)。它们的最早祖先类型都在澄江化石库发现并先后发表在Nature杂志[102-108],其中最受关注和影响深远的当属“天下第一鱼的发现”[103,105-106],Nature杂志同期做了“抓住天下第一鱼”的专题评论[109]。该研究成果目前大量入选百科全书和系列国内外高校教材。
蜕皮动物的系统分析和基因组系统研究也已经成熟,分支框架基本稳定(见图6)。在不同的研究中均分为三大单系分支:有吻突动物类、线形动物类和泛节肢动物类[89-90,101],在这个超门有大批国内学者先后做了大量工作,代表性文章较多[110-116]。有吻突类主要是底栖游移型,泛节肢动物主要表栖爬行和底栖游泳类生活 (见图12)[117-119],最近通过三维显微CT应用在节肢动物研究方面具有明显的技术引导性[120-122]。
门类最多、形态差异最大、支系演化关系最不稳定的超门类群,当属冠轮动物(又称螺旋动物)[89,101,123],与蜕皮动物一起构成原口动物两大超门分支。其重要门类组成包括触手冠动物(腕足动物、帚虫动物、苔藓动物)、担轮幼虫类(软体动物、环节动物和纽形动物)以及大量奇异化石类型、罅隙型生物和体腔动物[124-125]。冠轮超门分支在两侧动物中虽然支序分析相对最不稳定[126-127],但基本涵盖现生的13个动物门类,各门类之间系统关系多变[89, 123,128-129],相对蜕皮动物和后口动物内部分支关系明显不稳定(见图6)。但是,总体而言可以分为3个支序分支,即扁形动物、群居动物和担轮动物三大类群[101,130]。海洋中化石最多的当属古生物学者熟知的腕足动物[131-151]、苔藓动物[151]、软体动物[152,167],但也有内肛动物[155, 168]、帚虫动物[168]、环节动物[153-154,168]以及星虫动物门[156,168]。最新的相关进展和评论也有报道[130],尤其是苔藓动物门的发现[172],改变了传统的地球历史时期寒武纪演化动物群和古生代动物群的地史演化框架。生态上,冠轮动物在海洋环境中基本都属于底栖生物,属于地地道道的“草根动物”,参与海底动物碎屑骨骼和礁体的形成,在海底硬化和改造发挥有重要的作用[157],其中如苔藓动物、软体动物砗磲、环节动物龙介虫、蛰龙介虫和圣诞树虫等有着“海底之花”的美誉。
3.1.2 寒武纪早期后生动物出现和地史演化 那么,寒武纪大爆发期间到底出现了多少个生物类型(门),它们的生活方式如何,即如何取食、运动和居住行为怎样?所有这些生态问题一直是进化生物学和古生物学以及分子进化生物学家关注的问题[123,128,158]。
虽然最早的多细胞后生动物很可能以埃迪卡拉生物为代表,但可靠化石证据表明,基础动物和两侧对称动物最早出现在寒武系底部的纽芬兰统。纽芬兰统也是我们常说的寒武系底部的前三叶虫化石地层,其化石丰富,在我国主要以富含在磷块岩或者灰岩地层中的大量小壳化石而著名[159-162]。这些小壳化石除了基础型海绵动物类、开腔骨和刺胞动物外,主要还包括两侧对称动物中的冠轮类原口动物,壳体或者壳体部分碎片[29,31,163], 包括原牙形刺、软舌螺和似软舌螺类、腹足类软体动物(见图13)。最近通过陕南宽川铺组磷块岩中微体壳体化石的研究表明,幸运阶的晚期可能还出现有大量的四射珊瑚、锥石类[98-100,165,166]和少数类型的有吻突动物(scalidophorans)等蜕皮类群[169-171]。总体而言,寒武纪纽芬兰统的壳体动物化石除了大量的基础动物外,数量最多的当属冠轮类动物,包括软体动物腹足类、软舌螺类和各种托莫特壳类。在地层上,腕足动物似乎也早于三叶虫等大量的节肢动物而出现[161-162,172-173](见图13)。
在三叶虫出现的前后,叶足动物微网虫、高肌虫和鳃曳动物似乎均快速开始大量辐射,但具体时间节点和前后顺序还没有得到明确的研究证实。但是,这些蜕皮类群的辐射主要以澄江动物群的研究类型为代表(见图13)[29,168,174-175]。
全面总结蜕皮类群和大量冠轮动物的化石记录[29,160,162,176],尤其需要注意的是在地史上化石十分丰富的苔藓动物在寒武纪大爆发期间一直缺乏化石记录[177],因此苔藓动物被认为是奥陶纪大辐射的产物。然而,最近我们在陕南镇巴地区小洋剖面西蒿坪段的系统研究取得了重大突破,发现了干群的次生磷酸盐化保存的苔藓动物化石,其规律性的虫室分布和模块化生长表明了苔藓动物的属性,其虫室形态稳定、整齐划一。该研究最近发表在英国Nature杂志[151],论文送审后2周意见全部返回,Nature杂志主编Henry Gee第一时间来信认为该文是他从事Nature主编34年以来同行评审最快、成果认可度最高的学术论文,因此加速了论文的发表[151]。Nature杂志同期以“苔藓动物最终发现在寒武纪”为题做了专题科普评论和肯定,认为寒武纪苔藓动物起源的最大谜团已经解开,指出苔藓动物以后工作的方向和科学问题主要将探索寒武纪早期到奥陶纪苔藓虫的地质演化的多样性历史和地质历程[78]。
基于早期的综合性研究[29-30],地球动物在寒武纪大爆发“主幕”时期出现至少21个动物门类,分属286个物种,包括了所有的触手冠动物(lophophorates)门(腕足动物、帚虫动物和苔藓动物门)、担轮动物(软体动物、环节动物多毛类和星虫动物)和所有的节肢动物和后口动物分支类群(见图6, 13)[171]。所有的发现和证据进一步支持了舒德干院士提出的三幕式寒武纪大爆发假说[29,180]。宏观而言,多细胞后生动物在前寒武纪—寒武纪界限附近历时大约4千万年,导致地球后生动物树三大亚界(基础动物、原口动物和后口动物)、四大超门类群(基础动物、后口动物、冠轮动物和蜕皮动物)全部成型(见图6)[29-31, 179]。无论相对漫长的46亿年的地球历史还是38亿地球生命演化史,地球动物树(界)成型历时四千万年具有明显的瞬时性和爆发性特点(见图11)。相反,与地球陆地生态系统中植物界的起源、发展和最终成型演化模式明显不同,演化速度慢,历时大约3亿年(见图1)。寒武纪大爆发期间后生动物的阶段性和突发性出现过程和模式(见图11)[160],已多有研究[29, 161-162, 176, 180,183],本文故不再详及。
3.2 寒武纪大爆发初级进化生态系统形成和海洋群落复杂性演变
生态上,最原始的多细胞真核动物消费者当属埃迪卡拉生物[43,184-185],主要出现在5.6~5.4亿年前的埃迪卡拉纪晚期。无论其亲缘关系如何,埃迪卡拉生物群在生态还是在生物多样性方面,对地球生态系统演化都十分重要,代表着地球异养型动物型消费者的诞生。埃迪卡拉型生物与寒武纪早期两侧对称动物一起构成了寒武纪大爆发的3个阶段(见图6, 15)[29,180],导致地球动物树三大亚界分阶段依次爆发成型。更重要的是,包括埃迪卡拉生物在内的寒武纪生命大爆发事件代表着地球生态系统开始从菌藻类初级生产者牵引的原始二极生态系统(生产者和消费者组成)(见图15),向以动物消费者为动力的三极生态系统(生产者、消费者和分解者)过渡。

图15 澳大利亚埃迪卡拉动物群野外考察和博物馆参考照片Fig.15 Field excursion and museum-visiting photos of Ediacaran fossils from South Australia

A 埃迪卡拉阿瓦隆组合生态图,显示底栖固着生活及其垂直群落结构(修改自文献[25]);B 埃迪卡拉纪高家山克劳德管生活方式及其微生物席支撑的垂直群落结构,引自文献[197]图16 埃迪卡拉生物垂直群落分层Fig.16 Tiering community dioramas for the Ediacaran Biota
随着工作的不断深入,埃迪卡拉生物群的类型和产地也不断地扩大[186]。总体而言,埃迪卡拉生物群是一类非矿化、无眼睛、缺乏附肢和任何组织器官的多细胞生物或基础型动物,生态上属于海洋表栖的异养型生物(epifaunal organisms)(见图15,16)。但它们个体大小悬殊,形态分异大[25,187],不同于微生物席群落,埃迪卡拉生物生态上垂直分层加速,变化较大(tiering complexity),生态空间从单纯的微生物席占有的二维海底表面向海底表面上下的三维空间伸展(见图16)。有证据表明,埃迪卡拉生物在一定地质时期内,时间越早个体越大,这可能与埃迪卡拉末期出现运动和游移的两侧对称动物直接相关联,它们导致海底扰动加强[33],影响或破坏了埃迪卡拉纪微生物席表栖环境(见图16)。
在埃迪卡拉纪晚期和寒武纪早期,生物开始发生矿化[188-189],在前寒武系—寒武系界限附近出现了大量微小的亲缘关系不明的壳体化石,统称为小壳化石(small skeletal fossils,SSFs)(见图17),代表着地球最早发生的生物矿化事件[190-191]。这些化石宏观上应该包括埃迪卡拉晚期的明星化石克劳德管(Cloudina),甚至全球分布的陕西迹化石(Shaanxilithes)[192-193]。磷酸盐化的不同壳体化石目前已在全球不同大陆的传统前寒武系—寒武系界限之下发现[188-189],最新的文献详见文献[194],支持了前寒武系界限附近的连续性。但是,壳体化石的出现和生物矿化的发生,在前寒武纪—寒武纪界线附近或者上下仍是一种地质突发事件,明显受环境控制或制约,值得多学科进一步深入探讨[195-196]。根据目前的生态研究,这些壳体化石很可能属于最早的礁丘附着型生物,固着在微型的块状叠层石礁上营滤食生活(见图16B)[197],一定程度上代表着生物开始改造地球,并影响环境向适合生物的栖息环境发展。不同的丘状小礁导致微生物席生态环境异质性增加,这样微生物席主导的均一性被打破,生态环境出现不同的微环境和生态位(niches),导致出现了形如稀树草原样零星分布的有机质和食物资源,分布明显不均。基于此,瑞典乌普萨拉大学、国际著名进化古生物学家Graham Budd提出了稀树草原假说[199](Savannah hypothesis, SH)(见图16B),认为这些不均等的信息和食物资源为寒武纪后生动物和钙化骨骼的出现搭建了生态平台[198,200]。寒武纪生命大爆发的实质在系统演化方面是两侧对称动物的爆发性快速辐射事件[30,174,201],生态上是以后生动物为主导的海洋复杂生态系统的初次建立事件[30,34,202-203],也就是地球初级消费系统出现的生物事件,导致完整的食物链初次在地球上显现(见图12,13)。 大量动物门类出现的同时自然是不同躯体构型生命的创新,伴随着动物生活方式多样,运动能力和生物矿化等生态事件发生[204-205]。
新的生态现象的发生必然源于复杂行为和新躯体构型的产生[266],但并非所有的躯体构型的出现都会引起生态创新或者群落生态变革(见图9)。因此,生物物种多样性变化与群落生态系统的改变并不具有直接的耦合关系[34,206-207]。其中,只有生态类型不同并对其他生物和环境有生态贡献或影响的的动物先驱,不断侵占或探索不同的海洋生态空间,才能吸引和帮助大量不同的动物尾随其后,导致整个群落占领新的生态位(见图12,13)。生态位不同的动物先驱通过造礁、壳体富集等生态工程进而改造地球环境(见图9, 12)。大量动物的出现和不同生态位的占有,最终导致生物生态重构、海洋生态背景改造和环境条件改善,进而改变地球环境向动物宜居性和多样性更高层次方向发展(见图1),这也是盖亚假说强调的主要方面。
在早期的研究中,学者们多过分强调生物对环境的被动适应,而忽略了动物对环境的改造和主动影响甚至改变。需要指出的是,相对陆地生态系统的植物界,除原生生物外,动物尤其是无脊椎动物是探索和改造地球的生物先驱。因为在显生宙的早期,也就是早古生代地球上除了微生物和原生生物藻类外,生态类型和营养方式最丰富的当属无脊椎动物亚界,这也是我们在讨论地球早期历史和早期生命演化时,同学们提问“为什么不讨论植物”的原因。
寒武纪大爆发诞生了后生动物三大亚界,21个动物门类,这些动物相互依赖,相互影响,相互竞争,就形成了直接的群落生态关系。生态关系根据性质的不同,可以分为生死对抗关系和合作共生关系,如捕食关系、共栖关系、对抗关系、寄生关系、包壳关系、幼体底质选择等等[141,150,208-211,225]。为了理解生物作用的过程和影响,寒武纪海底生物群落的组成和结构就变得十分重要,尤其是底栖生物(benthos)。这些底栖生物包括大量的壳体化石,形成了具有显著特色的寒武纪演化生物群(Cambrian evolutionary fauna)[212-213],主要包括三叶虫、非铰合类腕足动物、软舌螺和棘皮动物等壳体动物化石。壳体化石的出现是寒武纪区别于前寒武纪生物的主要特征,也是寒武纪生物群落的重要特点。矿化结构的发生也是动物与动物相互作用(固着、共栖、捕食、包壳和竞争)的生理基础,也是无机与有机相互交流的桥梁和纽带。
海洋环境的变化经常会受到生物活动和生活过程的影响,如农垦革命和底质演化等,进而作用或影响到一定环境中生物群落的构成和属性 (见图11~13)[214-216]。根据分析[217-218],不同的生物通过不同的活动或生物建造,如生物扰动或钻孔等行为,生物礁、壳体富集或介壳层(shell beds)作用于海底环境,从而影响其他生物的生活方式和营养结构。前寒武纪—寒武纪过渡期间微型和宏体骨骼化石的大量出现为寒武纪群落结构的发展提供了物质保障。这些壳体化石包括大量的带壳动物(如三叶虫、软舌螺和腕足动物),也包含丰富的底栖固着生物,如海绵等[33,219],构成了寒武纪进化生物群的主体。他们通过大量聚集的壳体或分散的壳体碎屑作用于海洋底质,促进了海洋基质的演化[84,195]。海洋底栖生物的相互作用同时也是海洋生态结构复杂度(ecological complexity)的反映和直接证据[220]。一定程度上,生态建造或结构(ecological structure)的建立和复杂性直接取决于关键物种(keystone species)的出现、繁盛和消失[207,212](见图9),而并不在于群落生物多样性的增加和下降[221]。在寒武纪底栖固着的生物中,三叶虫、腕足动物等壳体化石非常关键,它们的化石在寒武系出现早、丰度大[222-223],大量分散出现或者富集出现的壳体(shell concentration)对寒武纪早期泥质海底硬化发挥重要的作用,同时还充当底质生物(basibionts)为其他生物的发展提供关键的重力支持[222, 224]。更有甚者,大量聚集或原位生长的腕足动物壳体聚集成介壳层或者富集壳层,在寒武纪泥质海洋环境中,发挥着铺路石的作用,为其他底栖动物的生活拦洪避沙,为其生长繁盛提供了平台 (见图19)[150,225],甚至改变了海洋或海岸浅水环境[226]。
化石证据显示,埃迪卡拉纪后生动物的类型主要以基础动物或者刺胞动物等级的生物为主,生态相对比较简单,生物之间缺乏竞争和捕食,基本处于基础生态位(primary tiering)[221],生活方式主要以底栖固着为主[203]。根据早期的研究,埃迪卡拉生物的固着方式与显生宙的底栖固着生物也明显不同,主要可以分为:① 浅水沉积贴附型(sticker);② 吸盘型(suctorial disk);③ 栖息型(rester)[230],这些生物多以底栖滤食生活为主。随着农垦革命(AR)的开始(见图14),前寒武纪底质开始向显生宙混合底质演化(寒武纪底质革命)[84],埃迪卡拉生物形态已经不再适应寒武纪底质的要求,后生动物的形态也随之遽然发生变化,开始出现显生宙型的附着器官或者组织[227-228],如腕足动物肉茎等不同形态的附着器官,同时也出现了大量的底栖移动型的软体动物,开始在沉积物表面啃食(牧食)藻类和微生物[167]。之后沉积取食的穴居生物和游移生物才大量出现(见图12)。在寒武纪第二世的澄江动物群中,已发现多门类的生物可以附着和固着在三叶虫、腕足类和软舌螺类等不同门类的壳体碎屑上生活(见图18,19),一种生物附着在其他生物体上生活,在生态上就形成了生态分层,生物群落分层加速,生态空间利用效率明显提高。如上所述,以澄江生物群为代表的寒武纪大爆发“主幕”时期出现至少21个动物门类,286个物种,分属基础动物亚界(海绵动物、古杯动物、刺胞动物、栉水母类)、蜕皮动物(节肢动物、鳃曳动物)、冠轮动物(腕足类、苔藓类、软体动物类、多毛类和)可归入23个生态功能群[222](见图12),生活方式多样,不同的生物占据了不同的生态位。但是,表栖类型在群落中占主导地位,对比早古生代海洋群落的生活方式和营养方式类型,寒武纪早期生态系统内栖型、掘穴型动物相对缺乏,潜穴或者掘穴能力有限(见图20)[168,222,224,229,246]。

A~B 寒武纪第四阶关山动物群腕足动物壳体及其表面窃食寄生的管状动物复原图;A 原位群体介壳层;B 显示管状生物与舌形贝腕足动物之间的存在的窃食寄生关系(引自文献[150]);C~D 寒武纪第三阶澄江生物群鳃曳动物体表附着的小型蠕形动物共栖关系;C 复原图;D 吸盘古宿虫(Inquicus fellatus)化石形态特征、微观形态和EDX图谱分析(引自文献[208])图18 寒武纪大爆发期间壳体动物和蠕形动物之间存在的群体附生关系Fig.18 The host-specific symbiotic relationships of early Cambrian shelly and vermiform animals

A 初级分层动物腕足动物澄江仡佬萨贝(Eoglossa Chengjiangensis, Wang et al., 2014),通过纤细肉茎锚定在海底表面生活;B 次级分层者澄江顾脱贝(Kutorgina Chengjiangensis, Zhang et al., 2007),通过粗大肉茎附着在三叶虫碎片上生活;C 中高级生态分层者澄江龙潭村贝(Longtancunella Chengjiangensis, Hou et al., 1999);D 变化多层分布的马龙矿山乳孔贝附着在大型藻类或可能的寒武纪树形笔石动物上,引自文献[164,199] 图19 寒武纪大爆发期间腕足动物生活方式和垂直群落结构, 图示腕足动物成为寒武纪大爆发期间生态分层最为成功的后生动物,包括初级分层、次级分层和中高级分层Fig.19 The lifestyles and tiering complexity of early Cambrian brachiopods as exemplified from the Chengjiang Fauna (Stage 3), with the note that brachiopods were the first benthic metazoans that achieved their success in ecological stratification and tiering complexity by Cambrian Stage 3, encompassing primary and secondary, and medium-high height of tiers
3.2.1 寒武纪发大爆发多细胞动物取食结构和营养方式 生物生活方式的研究关注3个重要指标:运动方式、居住类型和营养方式(见图8, 20)。生物的营养方式有光合作用、表面吸收和摄食(主动和被动)。光合作用是植物获取营养的主要方式。微生物(细菌和真菌)主要依靠表面吸收获取营养,它们吸收营养的关键是扩大个体表面积。真菌的菌丝体就是为了扩大吸收面积。动物以摄食为主,有被动取食也有主动取食,摄食的关键是不同程度提高器官的活动和运动以提高取食效率。藻类和高等植物能自身合成营养物,属于自养生物类型(autotroph),动物属于异养生物(heterotroph)(见图2)。有些细菌属于化能自养型,不是通过日光取得能量而是通过化学反应合成有机物,当然,蓝细菌(蓝藻)和藻类一样是光能自养生物类型。在埃迪卡拉纪—寒武纪期间,除了蓝藻和藻类这些自养生物外,所有的埃迪卡拉生物和寒武纪动物都属于异养生物(见图2)。这些动物摄食微小的有机质颗粒或微小生物个体(浮游生物)的为微食性生物,包括滤食生物(filter feeder),如腕足动物、珊瑚等具有触手的滤食动物;碎屑取食者(detritus feeder)如腹足类、双壳类和部分棘皮动物等;以及沉积取食者(deposit feeder)(见图8, 12, 20),它们吞食海底泥沙中的有机质,如环神经类鳃曳动物等[230],这些生物能产生大量的生物扰动和泥质碎屑粪便[231]。
据目前的研究,埃迪卡拉生物生活上很可能是一类表面渗透滤食生活的低级动物,代表着地球早期生态系统中异养型的初级消费者的出现。即使一些管状化石甚至存在可能的狭长消化道结构[232],也是滤食生物。但是,他们的营养方式复杂,可能包括表皮渗透取食(osmotrophs)、腐生取食(saprotrophs)、悬浮取食(suspension feeders)、沉积取食(deposit feeders)、刮啃取食(grazers)(见图12),还有报道的一些很难想象的捕食和寄生现象[233-234]。
澄江动物群和寒武纪特异型保存化石的研究为理解不同物种的个体生活方式和群落生态提供了大量的数据和化石资料[86,235-236],但不同类型生物的个体生态,诸如幼体固着[141]、群体行为[238]和节肢动物孵育行为[239-240]等还需要深入的生态分析。澄江化石的研究表明,澄江生物群动物的生活方式至少包括内栖型(infaunal)、表栖固着型(sessile epibenthos)、表栖游移型(vagile epibenthos)、管栖型[237,230]、游泳型、浮游和假浮游生活型等。从生态类型的统计和分析可以看出(见图12),寒武纪大爆发不久,几乎所有现代生物的营养结构和生活方式都很快出现并发生辐射(见图13)[79,82],这样所有的动物生活方式和营养结构在早古生代海洋中都已经出现。之后,海洋生物群落不断演替并发生变化,生态类型和营养方式只是在不同生物或者之间再次上演,反映了相同生态压力下不同时期生物组合的演替(见图1)。
3.2.2 寒武纪大爆发垂直群落结构和生态分层(Ecological Tierings) 分析海洋生物群落结构复杂性有3个重要的指标和参数,包括生态分层(tiering)、取食方式(feeding)和运动能力(motility)(见图20)。取食方式和运动能力大家都比较熟悉,也容易理解。取食方式,如我们熟悉的悬浮取食、沉积取食、啃食和肉食等(见图8);运动能力有快速和慢速以及固着和非固着之分。其中生态分层相对生疏,是研究底栖生物群落的重要指标参数[241],显示底栖生物在生态环境中的垂直分布模式和生态空间的利用效率。垂直高度是一个重要的参数,0~5 cm属于低水平高度,但高于20 cm高度就可以属于高水平分层高度,5~20 cm属于中高水平。除了高度之外,动物根据能否主动利用其他资源或者建造可以分为初级分层者(primary tierers)(见图16)和次级分层者(secondary tierers)(见图18~20)。次级分层者明显是能够利用其他动物的壳体或者洞穴和钻孔使得其能顺利保持在沉积物和海水表面之上或之下的生态空间。与次级分层者相比,初级分层者只能自己直接作用于海底,或固着或钻穴。基于此,从而分析一定地史时间生物的生态群落复杂度和演化[33,242]。
很明显,埃迪卡拉生物群的群落高度变化很大,从普遍存在的20cm到最高的120 cm,落差明显很大[25]。但是,所有的埃迪卡拉生物类型都属于初级分层者(见图16),只能直接作用于海底,期间的生物未能发现相互作用、相互附着和支持的生态现象。
腕足动物是古生代,尤其是早古生代底栖动物群落的重要化石代表,个体数量大、类型丰富,不同地质时期的生态类型化石易于收集,也便于比较(见图19,20)。澄江动物群的发现为研究腕足动物等寒武纪底栖动物群落和生态提供了重要依据[132-138]。根据早期化石研究[132-141,164],寒武纪腕足动物已发育有多种不同的生活方式:包括肉茎附着型(pedicle attachment)、肉茎锚定型(pedicle anchoring)、黏附型(cemented)、自由平躺型(free lying)、假浮游型(pseudoplanktic)和半内栖型(semi-infaunal)。它们已经初步适应了软质(soft substrate)与硬质(hard substrate)海底表面几乎所有生态空间[134-144,164,246](见图19,20)。寒武纪早期的舌形贝类腕足动物通常具有细长的肉茎,能将矿化较差的壳体锚定在海底,其生活方式好比海底的风筝,如玉案山舌孔贝(LingulellotretaYuanshanensis, Zhang et al., 2020)和澄江仡佬萨贝(Eoglossachengjiangensis, Wang et al., 2014)。此外,舌形贝型的腕足动物纯真滇东贝(Diandongiapista),其壳体厚度较大,矿化程度高,生活时通过纤长的肉茎直接将壳体锚定在沉积物和海水表面,壳体前端向上营滤食生活。然而,另一些腕足动物却长有粗大的肉茎,肉茎末端直接可以附着在其他动物壳体表面或者壳体碎屑上生活,如澄江龙潭村贝(LongtancunellaChengjiangensis)和澄江顾脱贝(KutorginaChengjiangensis)(见图19)。更为有意思的是,在澄江动物群的晚期,在滇东马龙地区玉案山组的顶部发现了一些小型的舌形贝型腕足动物——马龙矿山乳孔贝(KuangshanotretaMalungensis)[164,244],它们可以通过纤细的肉茎将壳体直接附着在大型的藻类或者半索动物笔石动物的笔石枝表面,形如挂在树枝的苹果(见图19D)。这种生态空间的扩张,为乳孔贝在寒武纪晚期—奥陶纪的辐射奠定了基础。这表明腕足动物可以通过其浮游幼虫利用其所有底栖生物的表面生活。因此,腕足动物是生态系统中最早建立生态分层(tierings)的底栖后生动物,不仅包括直接作用海底的初级分层(primary tiers),还爆发性出现了依靠其他生物碎屑生活的次级分层和中高级生态分层(medium-high levels of secondary tiering)[39,141-142,243,164,210,245](见图19)。对布尔吉斯化石库腕足动物的生态学研究发现,寒武纪腕足动物与其他动物如海绵动物,存在现代昆虫中广泛存在的拟态现象[210,245],它们壳体附着在海绵动物上,利用壳体边缘细长的刚毛模仿海绵动物的骨针伪装自己,以逃避其他捕食动物捕食的危险(见图21)。此外还有的个体可以附着在其他浮游动物的身体和壳体表面,形成假浮游生活现象[210,245-246],成为寒武纪动物相互利用共栖生活的典型范例。
底栖生物除了适应寒武纪混合底质外,大量壳体化石的出现对寒武纪基底的改造也发挥了重要作用[130,247]。根据 Erwin[217]和 Erwin and Tweedt[218]的分析,不同生物通过不同的行为活动,如生物礁、壳体富集或介壳层(shell beds)、生物扰动或钻孔等行为作用于海底环境,从而影响其他生物的生活方式和营养结构。大量出现的分散壳体碎屑或者聚集的壳体作用于海洋底质,促进了底栖固着悬浮滤食型生物群落分层,出现了大量的次级分层结构生物(secondary tierers),生态空间利用(Ecospace utilization)效率显著增强(见图18,19)[141,150,243],加速促进了海洋生态环境的演化和代谢[84,195],但海水/沉积物表面之下的穴居或内栖行为有限,舌形贝类腕足动物还未出现穴居内栖生态行为[134,164,246](见图20)。底栖海洋动物生态分层和垂直群落结构代表着早古生代甚至显生宙生物群落和生态系统复杂度的重要内容,因此,其复杂度可以最早追溯到寒武纪大爆发期间[235,249],无疑使早期海洋底栖生态群落分层的研究更加全面[139,141,241,243]。
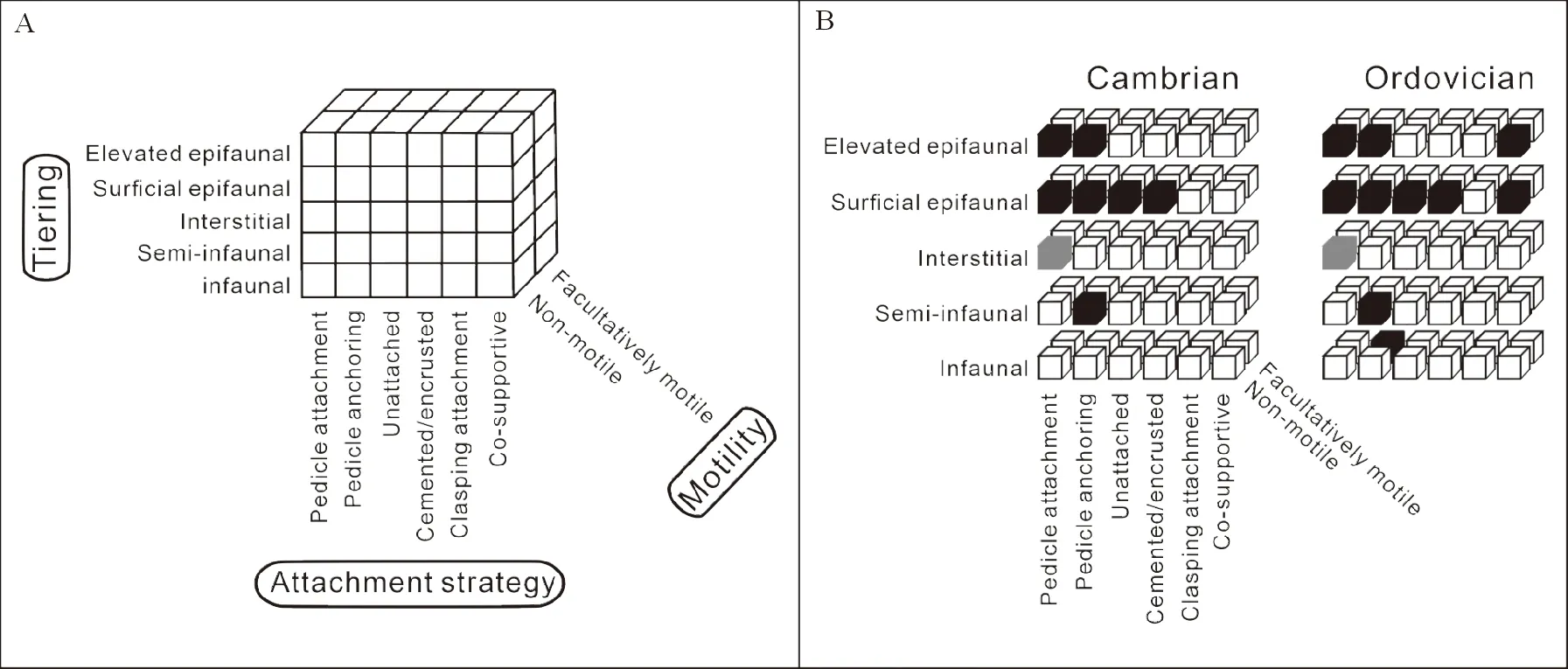
A 寒武纪腕足动物生活方式和生态空间分布;B 奥陶纪(早古生代)腕足动物生活方式和空间分布图20 寒武纪与早古生代腕足动物生活方式和生态分布对比图(引自文献[229])Fig.20 Comparative studies of the lifestyles and ecospaces utilized by Cambrian and Ordovician brachiopods (from [229])
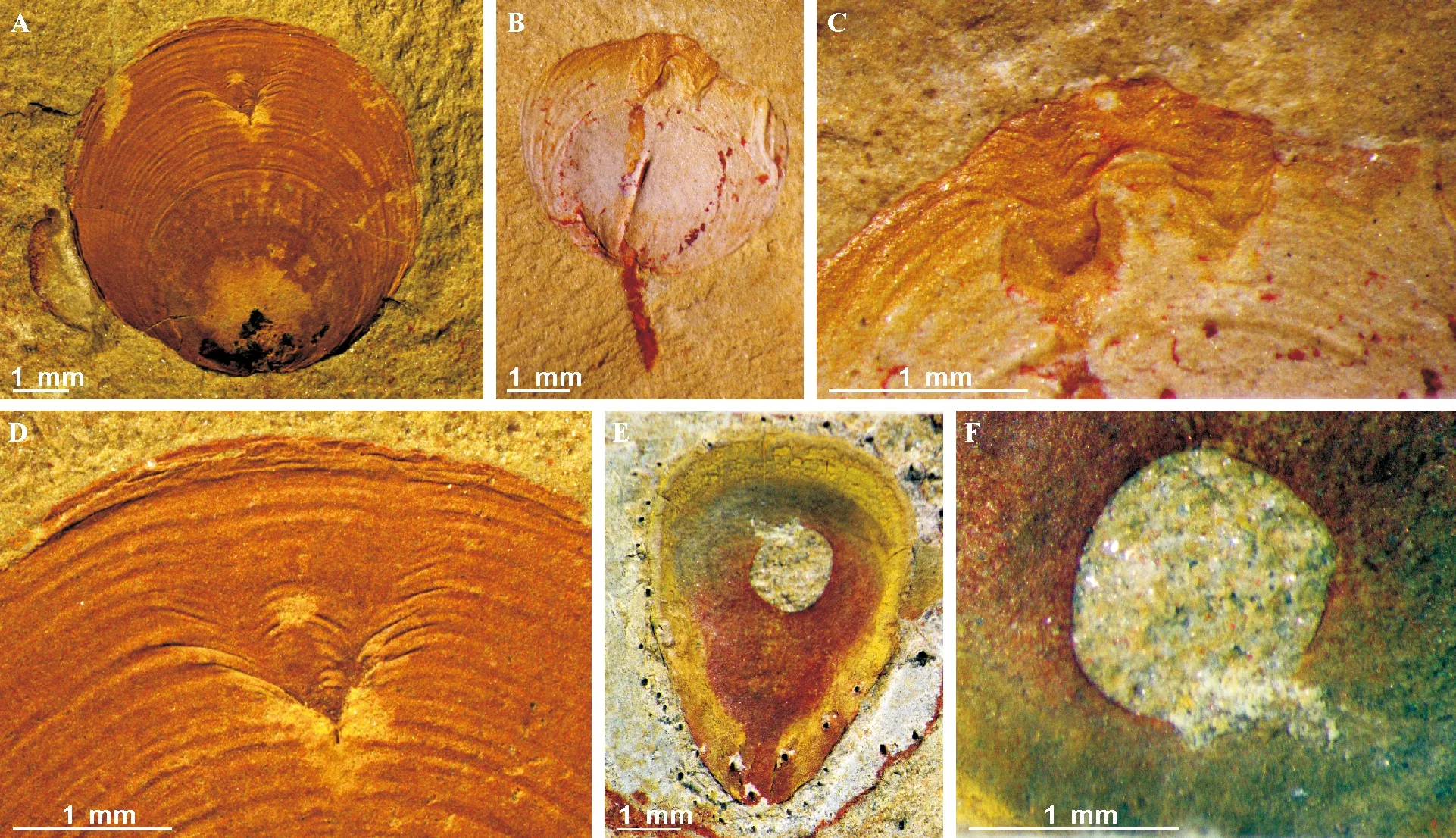
A~D 舌形贝型腕足动物乌龙箐新圆货贝(Neobolus Wulongqingensis, Zhang et al., 2020),显示壳体边缘修复的碎壳型捕食记录[144]; E~F 边缘圆润保存的圆形捕食孔保存在舌形贝型腕足动物马龙始圆货贝Eoobolus Malongensis(Rong, 1974)上, 系统修订详见文献[149]图21 寒武纪第四期关山动物群腕足动物壳体上保存的修复型捕食记录Fig.21 Repaired durophagous shell damages preserved in brachiopods recovered from the Guanshan Biota (Cambrian Stage 4), eastern Yunnan, Southwest China

A~C 峡东地区寒武纪天河板组核形石古杯礁;D 峡东地区艾家河剖面水井沱组单体古杯化石[173]图22 我国寒武纪早期的古杯礁体和单体化石。古杯通常认为是地球生态系统工程建设中最早的造礁后生动物类群,但在我国的寒武纪古杯礁体发育较弱。同时其礁体形成规模相对其他后生动物建造,如腕足动物介壳层在时间和规模上均不占优势Fig.22 Cambrian Archaeocyaths from lower Cambrian of China. Archaeocyath is generally considered among the earliest-reef-building metazoans during onset of the Ecosystem Engineering on Earth. However, the Archaeocyath reefs and their associated organism-inducing buildups are less-developed than those resulted from other animals,for examples, aggregated shell beds of brachiopods, either in earliest occurrences or in geological scales of buildups
3.2.3 寒武纪大爆发壳体富集(介壳层)和及其生态关系 群体(colonial)、包壳(encrustations)和壳体富集的介壳层(shell beds)等重要的生态现象通常认为出现在寒武纪中晚期到奥陶纪,是奥陶纪大辐射的重要生态现象。在寒武纪,古杯动物通常被认为是最早造礁的多细胞后生动物,在西伯利亚和澳大利亚都有很好的保存和研究[250, 251]。在我国陕南福成和鄂西天河板组有很好的保存,但并不壮观(见图22)。在我国滇东澄江动物群和鄂西水井沱组都有单体古杯化石报道(见图22D)。与澄江化石库的其他化石发现相似,不同属种的化石通常单属、单种出现,多个属种的化石仅发现在受到水流搬运影响的背景层化石中。这些不同类型生物的分布主要是地形地貌的异质性决定的,呈现斑块状分布,因此有学者将其称为稀树草原(savannah)群落。然而根据滇东地区的研究,到了寒武纪沧浪铺期的红井哨组顶部,情况发生了很大的变化。在马龙地区红井哨组的顶部,以及昆明地区及其周围不同区域的乌龙箐组广泛存在大量的腕足动物壳体介壳层(shell beds)(见图18A)[151,225,247,253]。大量出现的腕足动物壳体,在海洋基质的改造和硬化方面发挥了重要的作用,发挥了铺路石的作用,使其他底栖生物的繁衍和生息成为可能。在寒武系第4阶的关山动物群中,腕足动物开始超越三叶虫成为海洋底栖生物群落中最重要的基质生物(basibionts)。其大量出现的壳体已经超过三叶虫等其他寒武纪演化动物类型[225,252],成为寒武纪生态系统建设的贡献者(ecosystem engineers),为加速其他表栖固着滤食生物(epibionts)的发展提供了更多的固着平台和生活空间[141]。大量富集的壳体(介壳层)加大了寒武纪软基质海洋基质环境(soft-substrate)的改造和硬化[226,254]。有趣的是,在沧浪铺期乌龙箐组不仅仅存在大量的腕足动物壳体富集层,还在大量的壳体化石上发现存在着数量较多的蠕虫状管状生物。统计分析发现这些管状生物沿着壳体边缘向外辐射状排列,主要集中在壳体两侧40°~70°的位置,与壳体内部纤毛环的分布和血管(脉管)的分布高度一致,管子前端可能长有触手(见图18B)。这些虫管簇状附着生活在腕足动物壳体表面,并对这些腕足动物的生长造成极大的负荷和负面影响,影响了其壳体的正常发育。相对未感染的腕足动物个体,被固着的腕足动物生物量减少至少26%,统计分析表明,这些壳体表面生活的虫管与其附着的腕足动物存在明显的寄生关系,属于窃食寄生关系类型(kleptoparasite),代表了地球上最原始的寄宿关系[150]。这个发现获得了中国古生物学会2020年度十大进展[254]。统计发现,这种虫管只附着在乌龙箐新圆货贝一种腕足动物壳体上生活,对其他同期相似的腕足动物和其他生物,如三叶虫不发生感染,因此属于专性寄生虫[150]。大量出现的腕足动物和寄生生物化石,表明在5.4亿年前的寒武纪大爆发后不久,动物便需要与寄生虫作斗争。相似的专性共生关系(commensalism)最近还发现存在于蠕形鳃曳动物化石中[208]。研究人员在环饰蠕虫和马房古蠕虫类两大类蠕虫动物的标本上发现了成群的保龄球状的附生生物——吸盘古宿虫(Inquicusfellatus)。很明显,这种关系不明的附生生物只共生在生活方式相似的这两种蠕虫身体表面。研究者认为,该蠕虫共生关系已经具备了宿主专一性和宿主转移性两重生态关系(见图18C~D)[208]。此外,澄江化石库方形古虫和楔形古虫前体两侧的鳃孔两侧也有椎管状的小型生物分布,具有现生海洋中现代污损生物管栖多毛类的属性[209]。因此,寒武纪大爆发之后很短时间内,动物群落生态关系已经相当复杂,与后期的早古生代生态压力或生态动力区别不大[255]。
3.2.4 寒武纪大爆发捕食压力和对抗关系 化石证据显示,捕食关系在地质历史中最早可能发生在前寒武纪—寒武纪界线附近的壳体化石表面[256],但有学者认为捕食关系有可能在几十亿年前的原核和真核生物之间已经存在[257]。毫无疑问,随着地球初级消费者的出现,尤其是动物的出现,生态系统中必然存在着不同的捕食关系。但是,这里讨论的主要是捕食压力和捕食关系的存在意义,即不是讨论初级捕食者与生产者(菌藻类)之间存在的营养关系,而是消费者与消费者,也就是一种动物捕食另一种动物的关系[258-259]。在前寒武纪—寒武纪界限附近的壳体化石上经常会发现微型的钻孔或者溶蚀型微孔,虽然可以解释为可能的捕食关系,但也有早期微生物(分解者)或者后期成岩微生物造成的可能性。然而,在澄江化石库中的节肢动物中发现有许多的动物受伤愈合的宏体疤痕,表明了壳体动物之间存在宏体捕食压力或者致命伤害的可能性[144,260-261]。但是,这些疤痕多发生在澄江化石中都发生在肉肥味美的纳罗虫和三叶虫等节肢动物中,在成千上万的“壳多肉少”的贝壳类如腕足动物化石中并未发现任何壳体损伤和修复的痕迹[144],表明在澄江动物群时期,捕食压力相对较弱。在澄江化石库之后的关山动物群(早寒武世沧浪铺期或寒武纪第四期),腕足动物的壳体上捕食痕和修复的疤痕随处可见(见图21),代表着从寒武纪早期到后期捕食压力增加的过程[144]。之后,在各种壳体化石上捕食痕迹在三叶虫和腕足动物发现较多[35,262-264]。
3.2.5 寒武纪大爆发遗迹化石其生态启示 前寒武纪—寒武纪界限附近的遗迹化石的研究,近些年取得了很大的进步[34,231,265-268]。如上所述,制造遗迹的动物能通过钻孔或者洞穴增加海底沉积物/海水表面,提高海底渗透率和通透性,从而为新的生物提供栖息环境。埃迪卡拉纪—寒武纪遗迹化石丰富,记录连续,似乎很好弥补了该时期体化石稀少和保存不佳的不足,有力支持了埃迪卡拉纪时期(560 Ma)动物的运动性和两侧动物活动的存在[23,26,268],其多样性和形态复杂度在埃迪卡拉末期和寒武纪幸运期明显增加,也很好反映了寒武纪早期动物发育体制构型的创新和行为的复杂性。在我国滇东寒武系石岩头组和中谊村段多有保存[269]。之后的澄江动物群研究发现海底表面已经开始出现了大量动物爬行、觅食和逃逸的痕迹,虫迹共存多有报道[270-271]。在滇东地区的乌龙箐组中遗迹化石更是随处可见[252,272]。近期,本课题组对关山生物群(寒武系第四阶)的系统发掘发现存在大量的遗迹化石,其中保存了颗粒清楚的粪便化石。粪便化石中的大量粪球粒还存在再次利用的现象,揭示出5.2亿年前海洋生态系统中复杂的营养策略和取食方式已经形成[231]。
3.3 早古生代动物初级进化生态系统的发展和完善
奥陶纪生物大辐射是继寒武纪生命大爆发之后,在生命历史舞台上演的又一幕重大生物演化事件,表现为寒武纪早期出现的后生动物门类的生物多样性在目、超科、科、属、种级别开始急速增加[213,273-275],是以三叶虫、软舌螺和磷酸钙质腕足动物等重要壳体生物为代表的寒武纪演化动物群演替为以钙质壳腕足动物、苔藓动物、海百合和四射珊瑚为主体的古生代演化动物群的地史过程[276]。奥陶纪大辐射的主要特点为底栖生物礁大量繁盛,生物多样性快速增加(见图11~13)。奥陶纪大辐射期间生物多样性的增加主要表现在3个方面:分类单元多样性(α-diversity)、群落生态多样性(β-diversity)和生物地理多样性(γ-diversity)[213,273,277],三者之间具有一定程度的因果关系。期间,生态系统主要以滤食生物、造礁生物和海洋底质的硬化为主要特征,脊椎动物鱼类化石变得更为普遍,但海生无脊椎动物空前繁盛,明显占据了主导地位,浮游生物和游泳生物迅速崛起,生态系统类型不断完善。浮游生态系统以笔石动物的大量出现为标志,游泳生态系统以鹦鹉螺型的头足类震旦角石和喇叭角石为代表。根据目前的数据统计和系统研究[79,278-279],早古生代海洋生物的演化是一个长期的、渐进的地质过程。最近奥陶纪早期摩洛哥Fezouata动物群的发现进一步解释了奥陶纪和寒武纪动物群的相似性和演化连续性(见图11~13)[255,280]。早古生代海洋生态系统经过寒武纪大爆发、奥陶纪浮游生态系统演化、奥陶纪底质演化和志留-泥盆纪游泳型大型生物革命等不同生物事件,底栖固着型、底栖游移型、浮游型和游泳型生态系统发展稳定,形成了典型的无脊椎动物为主的地史演化特色和生态优势(见图12,13)。其中,底栖固着型的动物有大量的腕足动物[225, 283-286]、苔藓动物[287-288]、棘皮动物、海绵动物[289-290]和层孔虫[281-282]等。底栖游移型生物主要为双壳类和腹足类[178-179]。造礁底栖滤食生物主要以苔藓动物、刺胞动物四射珊瑚类、海绵动物和层孔虫为主[79]。游泳的动物在出现大量的头足类震旦角石类之外,还出现了大型的高级捕食型脊椎动物有颌类[291](见图13)。从生态类型多样性考虑,寒武纪和早古生代动物生活方式和生态类型在很大方面也十分相似,生物建造和底栖生态工程都可以加以对比生态升级(见图20)[150,225,246,255,279]。在志留纪末期,以大型有颌类脊椎动物为代表的高级捕食者和高级消费者才大量出现[17,39],代表着地球现代海洋生态系统已经开始发育。志留纪之后,脊椎动物和高级消费者,如鱼类进一步发展,在晚古生代游泳型生态系统仍以无脊椎动物为主,菊石型头足类取代鹦鹉螺型角石类,在中生代后才让位于大型的海生爬行类和新生代的海生哺乳类,在此不予详及,详见下文。
4 陆地生态系统形成与地球现代进化生态系统建立
如上所述,地球最早的初级生产力发生在海洋中,以具有光合作用的蓝细菌(蓝藻)及其后来的真核藻类为主。在演化中,微生物和藻类的种类或许变化,但不同时间生产者与分解者在高阶元分类和营养方式上不变,因此初级生产者在海洋生态系统几乎变化不大并一直维持到现在。但是,海洋生态系统中消费者却千变万化,在早古生代主要以海洋无脊椎动物为主。寒武纪大爆发期间大多数现生动物门类都已经出现[171],无脊椎动物如三叶虫、软舌螺、竹节虫、笔石、腕足类、珊瑚类、牙形刺和软体动物腹足类、双壳类和头足类等繁盛在古生代形成了地史古生物的主要内容,在海洋生态系统中整个古生代甚至三叠纪早期无脊椎动物一直是海洋中的霸主或者生态雄主[292-293]。
然而,在寒武纪最早出现的脊索动物门(脊椎动物亚门)在志留纪之后迅速演化成为地史演化的标志性事件(见图1)。其中鱼类的发展演化令人瞩目,在内陆河流、湖泊或河口地区都反映了淡水鱼类大繁盛和征服大陆的进程。到晚泥盆世,鱼类在征服大陆中又迈出了一大步,总鳍鱼类向两栖类演化,格陵兰东部晚泥盆世发现的鱼石螈化石成为最早的两栖类。两栖类最终演化出爬行类,甚至到哺乳类各种脊椎动物。所有这些应得益于于古生代重要的生态系统创新事件——陆地生态系统的形成和发展(见图1)。自从泥盆纪或者更早的志留纪晚期[294],地球初级生产者发生很大的变化,陆地上出现了初级生产力更强的一类生物——植物界(苔藓类、蕨类、裸子植物和被子植物),地球陆地首次披上了绿装。植物从原蕨植物到石松类逐渐摆脱了水的束缚,形成了根茎叶的分化。到了中生代裸子植物进一步发展,植物垂直分层加速,生态空间利用效率空前增长。白垩纪晚期被子植物开始繁盛,在新生代开始在陆地植物中占统治地位。植物的演化似乎与海陆的变迁、超大陆的聚散和特提斯洋的形成和演化有很大的相关性。随着初级生产力植被从原蕨植物、节蕨植物、裸子植物到被子植物的演化,脊椎动物也随着从两栖类、爬行类到哺乳类的演化。期间两栖类在生活中离不开水域环境,受精卵需要在水中发育。中生代是裸子植物的繁盛时代,同时也是爬行动物最为昌盛的时期。但爬行类在石炭纪晚期出现,在摆脱海水限制(重力、生殖和呼吸)后,中生代部分类型又返回海洋,与两栖类相反他们在陆地生殖、海洋生活。同样,哺乳动物在新生代演化中除了部分冲向蓝天外,也有陆地生殖、海洋生活的各种大型动物。陆地植物的出现,枯枝落叶及植物残片经过分解,与岩石风化相互作用,植物根系在岩石表面可以开展生物风化和溶蚀作用,地球开始出现土壤。所有这些生物和非生物的作用过程,形成了海陆空相互呼应、相互影响的现代海洋生态系统,能自我修复、自我完善,因而繁盛至今。
5 结论和展望
生态系统是一定空间和时间范围内所有生物群落及其非生物环境的综合。生物圈是地球上最大的生态系统。生态系统与自然界的生物一样,并不是固定不变的,而是一个不断变化的动态系统。生态系统的动态包括生态系统的进化和生态系统演替两个方面。根据生态系统的地质背景和生物组成(生产者、消费者和分解者)的不同,本文将地球生态系统宏观划分为三大演化阶段(见图1),原始进化生态系统、初级进化生态系统和现代进化生态系统三大演化阶段。原始进化生态系统是以菌藻类初级生产者演化为主线的2极生态系统。 初级进化生态系统在原始进化生态系统的基础上,诞生了动物消费者,地球进入了以消费者需求驱动的海洋动物生态系统,无脊椎动物繁盛,生态类型繁多,鱼型脊椎动物剧增,3极食物链生态结构完善,出现了现代海洋生态系统所有的营养方式和结构。现代进化生态系统是地球在海洋生态系统的基础上诞生了陆地生态系统,植物成为生产力最为高效的原始生产者,随之鱼类登陆,进而四足动物(两栖类、爬行类、鸟类和哺乳类)起源演化。因此,四足动物的发展和演化是陆地生态系统演化的产物,地球进入海陆空交互、高效循环的现代进化生态系统阶段。
研究表明,最早植物起源于志留纪晚期,最早的被子植物却在晚侏罗世出现,植物界成型历时至少大约3亿年之久。然而,早期动物的辐射主要发生在寒武纪大爆发期间,地球动物树(门)历时4千万年快速成型,这与植物界主要门类演化长时间、慢演化明显不同。其原因和演化动力明显与动物之间存在的捕食关系(军备竞赛)和海洋动物的生态系统工程的快速发生和演化密不可分。但与之相比,植物演化明显受到环境和生境的影响和限制更为明显,例如昆虫的辐射明显在加速被子植物的演化中发挥着重要作用[295]。古特提斯洋的关闭导致海岸线和陆表海面积减少,陆地面积相对增加,浅海海洋生态压力增大,导致动植物开始登陆。在沧海桑田、海陆变化中,经过古特提斯洋和中、新特提斯洋等海陆环境的变化,被子植物在新特提斯洋开始形成之后并最终快速演化(见图1)。
对比地球生态系统的三大演化阶段,相互交织,逐步提高。地球原始进化生态系统诞生了地球海洋生态系统的初级消费者和分解者,是无机界和有机界相互作用的开端。地球初级进化生态系统承袭了原始进化生态系统的生产者,但诞生了地球上最早的宏体消费者——动物,动物通过生态系统工程作用于海底,引领地球朝着动物宜居性的方向发展。在早古生代,现代海洋动物生态进化系统的生态类型和营养方式全部建立,泥盆纪之后海洋生态系统再无创新,生态类型和营养方式都已固化,之后只属于生态演替范畴。但陆地生态系统属于全新,并诞生了植物界,陆地原始生产者出现,生存空间空前扩展。理解这些过程需要我们对比研究地史关键生命事件和大陆聚合、环境背景关系及其相关性。过去更多地强调环境对生物演化的控制和影响,很少关注生物如何影响甚至改造环境的过程,就如现在的造林植树改善环境一样。所以应该有新的理论如盖亚假说一样,重点探讨生命如何影响和改造地球和生态环境,向适合自己的宜居方向发展。例如,最新的《地形学杂志》报道鲑鱼的交配正在无意间改变了地形地貌。研究发现鲑鱼是一种溯河洄游的鱼类,它们在淡水出生,海水中长大和生活,性成熟后会跨越数百公里溯游而上,返回故乡交配繁殖。雌性鲑鱼在河床上挖洞产卵,导致河床松散易动,最终导致河床基岩松动不断侵蚀。对比模拟发现,鲑鱼的存在对河道垂直的改变可达30%,在上百年的地质演化中鲑鱼对地形和地貌的改变无疑产生了巨大的影响[296]。同样,螃蟹也发挥着相似的环境影响,最近《美国科学院院刊》《PNAS》发文,发现在气候变暖海平面上升的背景下,螃蟹的掘穴异常活跃,其行为能将原本紧密相连的草坪区变成孤立的水塘[297]。 该研究发现, 螃蟹是一种关键物种, 在气候变化的情况下, 它能以远超自身重要性的方式改变其所处的生态系统。 同样在远古时期的寒武纪早期, 大量的舌形贝腕足动物壳体沿着格陵兰东北150 km的海岸线连续聚集, 富集了大量的介壳层[226], 这些壳体像铺路石一样硬化了浅海海面,改变了当时的氧化还原环境。 该类腕足动物壳体在我国滇东地区更早的地层也有发现[151,225,247,253]。在5.2亿年前的寒武纪早期,大量富集的腕足动物壳体在滇东地区马龙—昆明—武定—会泽地区能覆盖6 000 km2的滇东海域,硬化海底表面,为其他底栖生物的附着提供了平台,加速了其他底栖海洋动物的繁盛、群落生态的复杂化和生态空间的利用效率[151]。因此,在以后的研究中需要继续关注地球宜居性演化过程中的关键化学和物理条件,生命起源演化与重大地质过程的耦联,还要更多关注海洋生命如何通过生物地质建造(生态系统工程)影响地球早期的演化过程,而不是被动适应。
因此,在开展地球深时研究时,应重点开展3个对比研究:
1)地球海洋动物生态系统起源与陆地生态系统起源演化对比,寒武纪大爆发出现了地球最早的动物,历时4千万年动物界(三大亚界、四大超门)成型,无脊椎动物成为早古生代生态系统主要的消费者和生态驱动者,这些动物如何适应地球早期海洋环境,它们又有什么样的生态工程影响和制约地球早期海洋生态系统的发展和宜居性演化?相比之下,植物作为高级初级生产者对陆地生态系统发生的过程影响和环境无疑有重要的制约作用,但植物界为什么慢成型(约3亿年),是受地球大陆构造格局控制还是不同地质时期的环境制约?
2)泥盆纪动物登陆与中、新生代动物“下海”对比研究,这些生态现象与古特提斯洋演化、潘基亚大陆形成和裂解,与海陆转换、海洋面积和陆地面积相对大小,海岸线缩短和延伸,温度变化有何关系等等?为什么不同地质时期相同门类的动物生态趋向不同,全球地质背景和环境对比如何,对现代全球变暖、海平面上升,海岸线附近动物活动明显加强有何启示?
3)现代的海陆空、多圈层(生物圈、水圈、岩石圈和大气圈)生态系统与地球早期的单一海洋生态系统对比,现代地球的海陆空都有生物和动物分布,加速促进地球的物质循环和能量转换,但显生宙早期地球只有动物和菌藻类生活在海洋中,菌藻类作为地球系统唯一的生产者。
无脊椎动物是早期地球唯一的消费者,也是营养需求和物质循环的引领者。因此,从寒武纪大爆发开始,地球才真正踏上了她的宜居蓝色之旅(见图1)。与陆地植物的绿化工程相比,早期海洋动物在生态系统演化的意义应该予以重视,加强动物工程生态相关研究及其对地球宜居性的影响无疑更具深远的意义。
致谢: 感谢西北大学地质学系董云鹏教授的有益讨论! 在成文过程中地质学系舒德干教授、韩健研究员对部分内容作了修改并作了大量讨论和有益建议,感谢团队青年千人Tim Topper和Luke Strotz的工作和讨论。在此表示衷心的感谢!
猜你喜欢
减速顶与调速技术(2022年3期)2022-02-11 11:26:36
科学(2020年4期)2020-11-26 08:27:02
小哥白尼(趣味科学)(2019年10期)2020-01-18 09:16:24
制造技术与机床(2019年4期)2019-04-04 12:22:10
广东技术师范大学学报(2016年5期)2016-08-22 09:07:26
焊接(2016年2期)2016-02-27 13:01:22
小学科学(2015年2期)2015-03-11 21:52:45
小学科学(2015年1期)2015-03-11 14:17:16
生物进化(2014年3期)2014-04-16 04:36:42
科学大众·小诺贝尔(2009年9期)2009-10-23 05:26:54
