
水稻‘越光’品种的“前世今生”
2021-12-26 06:43王伍梅王辉张效忠杜士云
农学学报 2021年6期
王伍梅 王辉 张效忠 杜士云
摘要:‘越光是一种短园粒水稻品种,自1953年在日本一开始推广种植,就是日本人最喜欢的大米。即使大面积推广种植50多年之后的现在,由于其米质优良和长期形成的消费口碑,也依然是日本水稻最大种植推广品种。为全面了解‘越光品种育种过程和其特有的农艺性状以及控制有利性状基因组信息,我们以“Koshihikari: a premium short-grain ricecultivar - its expansion and breeding in Japan”文章为最主要参考文献并加以摘译,对其历史进行梳理,同时对当前中国优质稻育种提出了个人的思考。
关键词:水稻;‘越光;米质;QTL;分子育种;优质品种
中图分类号:S511文献标志码:A论文编号:cjas2020-0261
“Past and Present”of‘KoshihikariRice Variety
Wang Wumei, Wang Hui, Zhang Xiaozhong, Du Shiyun
(Institute of Rice Research, Anhui Academy of Agricultural Sciences, Hefei 230001, Anhui, China)
Abstract:‘Koshihikariis a short-grain rice variety. It has been promoted in Japan since 1953 and is the favorite rice of Japanese. Even after more than 50 years of widespread planting, it is still the largest variety of rice planted and promoted in Japan due to its excellent rice quality and long-term consumer reputation. To fully understand the breeding process of‘Koshihikarivariety and its unique agronomic traits, and control the genomic information of favorable traits, we used the article“Koshihikari: a premium short-grain rice cultivar -its expansion and breeding in Japan”as the main reference for excerpts and translates, sorted out the history of the variety, and put forward thoughts on the current breeding of high-quality rice in China.
Keywords: Rice;‘Koshihikari; Rice Quality; QTL; Molecular Breeding; Good Quality Variety
0引言
1944年,在日本的新澙县农业试验场,育种家用当时北陸地区的一个抽穗早,产量高,但抗性差,极易感染稻瘟病的‘Norin1(‘农林1号),与当时拥有最强抗稻瘟病的‘Norin22杂交。在新澙县长冈国家农业实验站种植3000个F2群体,从中选择65个优良单株。1948年,20个F3株系种在福井农业试验站,然而1948年6月28日,福井地区发生7.1级大地震,完全摧毁了这座城市,并且大部分育种材料都丢失了。幸运的是,长岗的F3株系幸存下来了,他们从中选择5个优良单株,又从F4株系中选择了最优良的株系逐年加代纯合,直至1953年,选择到一个F8的优良株系,被命名为‘Etsunan17,进而在14个县种植,以考验它在不同地区的适应性。‘Etsunan17在1956年成为了新澙县的主推品种,并被定名为越光(图1)。鉴于越光品种良好表现,越光被农业部、林业部、渔业部(福井县政府)注册为优质品种[1]。
1越光水稻推广种植情况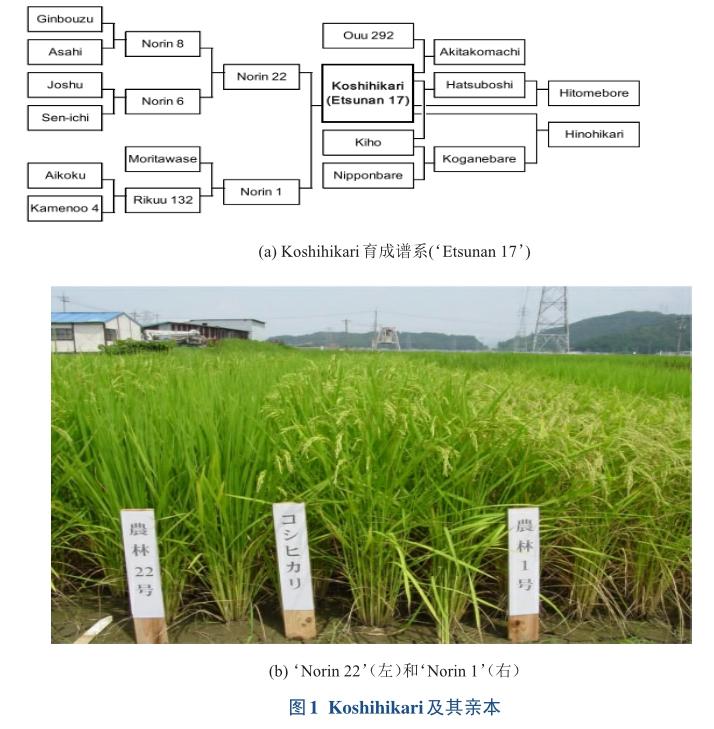
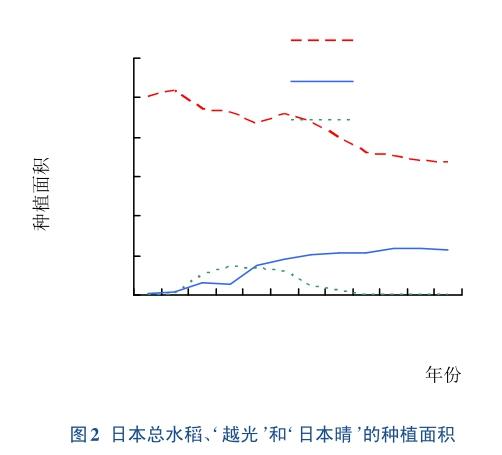
越光在新澙县推广种植之后,分别于1956、1957、1959年在东京附近的3个县——千叶县,栃木县和茨城县也相继被推广,并作为他们主要的栽培品种之一。1960—1961年,越光又在日本南部地区的鹿儿岛县和宫崎县推广种植。由于这2个县处于日本较温暖的地区,相对其他各地提前成熟,每年新米上市早。虽然越光产量表现一般,但市场对该大米需求却很大。1966—1969年,日本水稻大丰收,政府的储备中就持有55300万t旧稻米,供大于求。同时,经济发展,人们对水稻需求从追求数量到追求质量。恰巧,‘越光大米品质优良,满足了这一需求。20世纪70年代,日本西部和北部地区均采用‘越光作为主要栽培品种。自此越光在日本推广种植更加普遍。1979年,‘越光的栽培面积达到了297000 hm2,并自那时以来,一直保持着最高的位置[2]。截止2006年,越光的栽培面积达到了最大,约为640000 hm2(图2)。在目前日本排名十大推广品种中,越光的种植面积占日本全国水稻种植面积的36.2%,远远超过其他品种(表1)。
‘越光的种植面积和产量远远高于日本的其他大米归功于其优良的食用品质。在日本的112个品种中,其食用品质评为最高。‘越光米圆润饱满,色泽度高,直链淀粉和蛋白质含量低,黏性高,香甜度高。无论是在外观、气味、粘性和整体评价,煮熟的‘越光比‘日本晴更柔软,更香、更有粘性。然而,‘越光也存在其他农艺性状的缺点,例如:茎秆长,抗倒伏性差,叶片和穗部又易感染稻瘟病。尽管‘越光有一些抗性基因,包括Pish和Pik-s,但由于稻瘟病菌不断进化,‘越光的稻瘟病抗性比较差。‘越光对细菌性叶枯病表现出稍强的抗性,但易感染褐色条纹病。近些年,随着全球气候变暖,高温导致了其质量下降[3]。
2‘越光品種有利的性状QTL
‘越光品种在育种上的巨大价值,也引起人们对其控制有利性状的基因和整个基因组构成产生了极大兴趣。最早育种学家对为克服茎秆倒伏以及病虫害抗性差这些缺点,利用‘越光进行诱变育种。目前,已经研究出29个品种[4]。像‘Hokuriku 100,是越光的半矮化突变体,其茎秆长度比‘越光短30%[4];‘Jikei58是‘越光原生质体的衍生后代,其茎秆长度比‘越光短20%[5];‘Milky Queen是一个胚乳暗白色突变体,胚乳中的直链淀粉含量(9%~12%)低于‘越光(17.5%)[6];‘Yumegokochi[7]也是越光原生质体的衍生后代,其直链淀粉含量比越光低1.5%;除此之外育种者已经培育出一个缺乏α-球蛋白和一个强大茎秆的品种以及胚乳中具有低镉积累的新品种等等[8]。
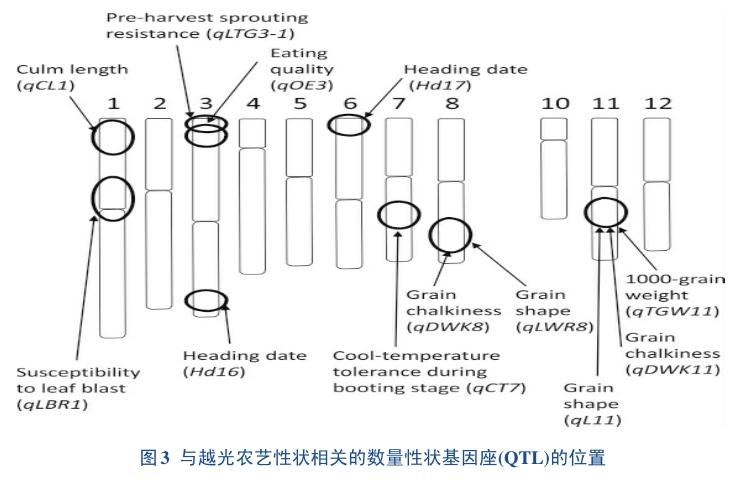
当然,对‘越光这种优异的遗传资源的研究,离不开QTL的定位分析。为了鉴定控制‘越光和‘日本晴重要农艺性状的QTL,研究人员构建回交自交系(BILs)和染色体片段替换系(CSSLs)2种类型的定位群体[9-10]。通过利用BILs和CSSLs,检测出关于越光的几个农艺性状的主效QTL(图3),主要包括品质、谷粒形状、垩白度、千粒重,茎秆长度以及对收获前发芽的强耐受性和孕穗期对温度的耐受性等等;特别一提的是,通过回交群体图位克隆到Hd16和Hd17[11-13],这2个基因不仅参与了‘越光对光周期敏感性的控制,而且也与米质相关,Hd17在越光中具有突变,也确保其在北方地区相对早熟,保持正常生长和收获[14]。另外,研究人员在1号染色体上发现了一个新的QTL与‘越光的茎秆长度和抗倒伏性有关[15];在3号染色体上发现了与‘越光食用品质有关的QTL以及与‘越光发芽收获前具有强烈的耐受性的qLTG3-1[16];在7号染色体上检测到了‘越光在孕穗期具有强耐寒性的QTL等等[17];这些QTLs对‘越光的优良品质和广适性具有很大的贡献。
3‘越光大米在当前日本生产和消费效应
自20世纪50年代以来,‘越光大米一直深受日本人的喜爱。然而,20世纪70年代以来,日本出生率下降和人口老龄化导致日本人口减少,对大米的消费量也在逐渐减少。随着面包等小麦产品的兴起,日本米饭的食用品质也日益多样化。年轻的日本人倾向于选择更硬的米饭,而不是像‘越光这样相对偏软的米。为了满足人们对大米的不同需求,日本水稻育种者正在尝试研究不仅可以增加消费量并符合饮食偏好多样化的新品种。目前有2种解决方案,一种是利用‘越光大米开发不同食用品质的稻米[18],另一种解决方案是“健康”大米[19]。并且随着日本对食品安全健康的意识不断提高,研究人员试图利用‘越光优良的品质,同时引进新的与功能米相关的突变。同时,‘越光基因组序列的解析,在基因组编辑时代,也为基因组选择育种提供了物质基础。
4‘越光水稻品种对中国优质稻育种的启示
随着‘越光在日本的种植面积不断增加,为增强品种的适应性,一些良好的农艺性状不断地导入到‘越光背景之中,例如它对不同环境的良好适应性、对收获前发芽的耐受性以孕穗期的耐寒性等等,这些特性远远超过了其抗倒伏性差、抵抗力弱的缺点。‘越光也是第一个通过下一代测序技术对全基因组进行测序的粳稻品种,作为日本战后水稻育种最重要育种亲本,这也为其他衍生出来日本水稻品种遗传系谱提供很好参考基因组同时,也为基因编辑育种提供精确信息来源。在‘越光遗传背景中不断开发出一系列同基因和近等基因系等,这些新发现和材料将促进基因组学辅助水稻育成优质品种。
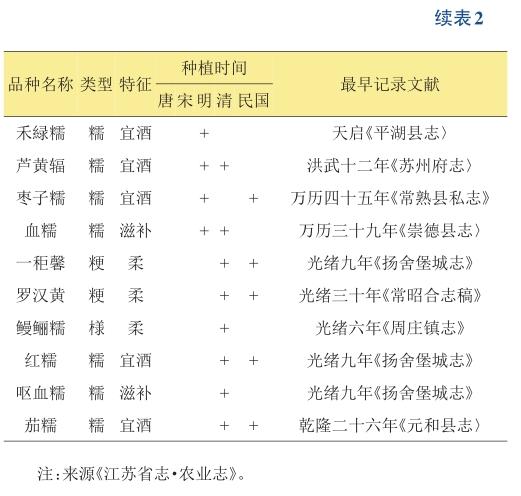
中国是水稻起源大国,水稻种质资源非常丰富[20]。不仅历史上有很多优质稻种的记载(表2),如今越来越多的优良品种也相继问世。利用这些现有的优质稻种资源,再适当参考其他国家优质的稻种资源,结合基因编辑和传统育种方法去挖掘有利的基因,逐渐改良优质稻不利的农艺性状。这不仅可以在优质米推广地区更高效率上创造出新品种,也可以提高优质稻生产的持续性和稳定性,促进优质稻的市场竞争力和相关产业的发展。此外,很多优质的籼稻品种通常都比较耐高温,因此关注这些品种,作为重要亲本资源去展开育种,对全球化气温升高,提高稻米生产安全也是十分必要的。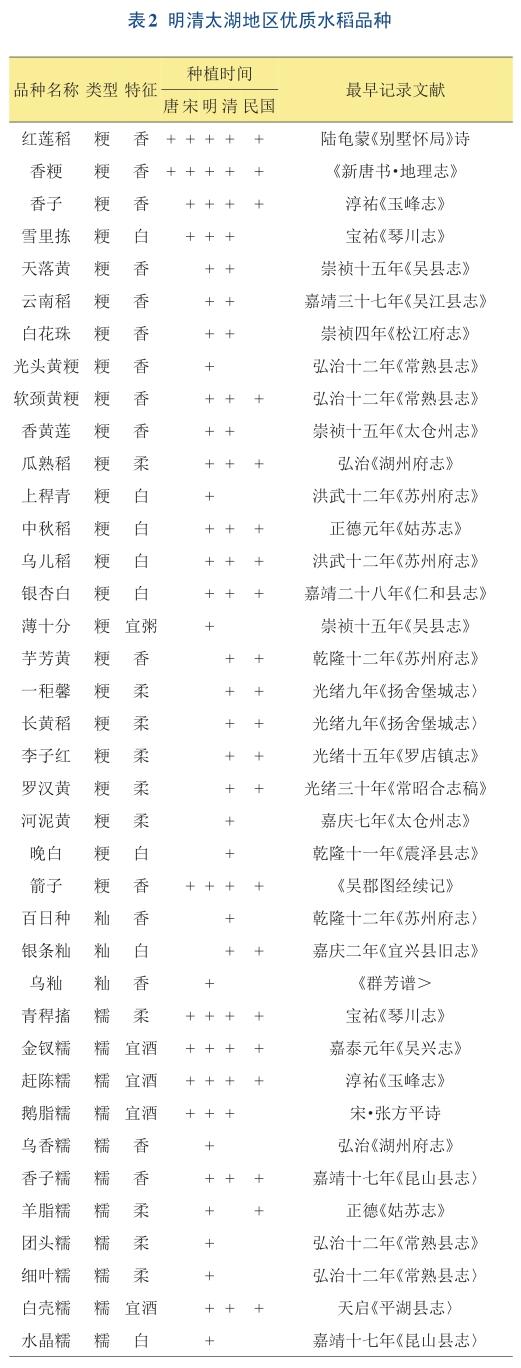
参考文献
[1]Fukui Prefectural Government. Brief descriptions of the Japanese leading rice variety,‘Koshihikari, and its lineage[C].Proceeding of‘2ndAsian Crop Science Conference,1995:2-3.
[2]Rice Stable Supply Support Retrieved from Organization Website[EB/OL].2017,http://www.komenet.jp/jishuchousa/144.htm.
[3]Takai T, Ikka T, Kondo K, et al. Genetic mechanisms underlying yield potential in the rice high-yielding cultivar Takanari, based on reciprocal chromosome segment substitution lines[J].BMC Plant Biology,2014,14(1):295.
[4]Tanisaka T, Tomita M, Yamagata H. Gene Analysis for the Semidwarfism of Two Mutant Strains, Hokuriku 100 and Kanto 79, Induced from a Rice Variety Koshihikari: Studies on the Utility of Artificial Mutations in Plant Breeding XVIII[J].Japanese Journal of Breeding,2008,40(1):103-117.
[5]Mori M, Taniguchi S, Noda H, et al. Studies on breeding using protoplast- derived plants in rice (Oryza sativa L.), Koshihikari variety, 2: Somaclonal variation in the seed progeny of prtoplastderived rice plants[J].Bulletin of the Shiga Prefecture Agricultural Experiment Station,1997.
[6]Ise K, Akama Y, Horisue N, et al. "Milky Queen, a new highquality rice cultivar with low amylose content in endosperm."[J]. Bulletin of the National Institute of Crop ence (Japan),2001.
[7]Sukekiyo T, Hayashi Y, Yahiro Y. Production of a unique rice variety "Yumegokochi" by protoplast breeding method and its application for commercial use[J].農林水産技術研究ジャーナル, 2003,26:44-47.
[8]Samoto S, Kanai D. Studies on the Mutation Breeding in Rice.: I. Short Stiff Mutations induced by Gamma-ray Irradiation to the Rice Variety Koshihikari[J].Japanese Journal of Breeding,1975,25(1):1-7.
[9]Matsubara K, Kono I, Hori K, et al. Novel QTLs for photoperiodic flowering revealed by using reciprocal backcross inbred lines from crosses between japonica rice cultivars[J].Theoretical & Applied Genetics,2008,117(6):935-945.
[10]UJIIE, Kazuhiro, ISHIMARU K. Analysis of Chromosomal RegionsAffectingAgronomicTraitsUsingNipponbare/ Koshihikari//Koshihikari Chromosome Segment Substitution lines[C]//Abstracts of Meeting of the Cssj CROP SCIENCE SOCIETY OF JAPAN,2010.
[11]Yano M. Natural Variation in Hd17, a Homolog of Arabidopsis ELF3 That is Involved in Rice Photoperiodic Flowering[J].Plant and Cell Physiology, 2012, 53(4):709.
[12]Hori K, Ogiso- Tanaka E, Matsubara K, et al. Hd16, a gene for casein kinase I, is involved in the control of rice flowering time by modulating the day-length response.[J].Plant Journal,2013:36-46.
[13]Kiyosumi H, Toshio Y, Masahiro Y. Genetic dissection of agronomically important traits in closely related temperate japonica rice cultivars[J].Breeding ence,2017,67(5):427-434.
[14]Yano M. Natural Variation in Hd17, a Homolog of Arabidopsis ELF3 That is Involved in Rice Photoperiodic Flowering[J].Plant and Cell Physiology,2012,53(4):709.
[15]Hori K, Kataoka T, Miura K, et al. Variation in heading date conceals quantitative trait loci for other traits of importance in breeding selection of rice[J].Breeding Science,2012,62(3):223-234.
[16]Hori K, Sugimoto K, Nonoue Y, et al. Detection of quantitative trait locicontrollingpre- harvestsproutingresistancebyusing backcrossed populations of japonica rice cultivars[J].Theor. Appl. Genet,2010,120(8):1547-1557.
[17]Takeuchi Y, Hayasaka H, Chiba B, et al. Mapping Quantitative Trait Loci Controlling Cool-temperature Tolerance at Booting Stage in Temperate Japonica Rice[J].Breeding Science,2001,51(3):191-197.
[18]Kobayashi A, Tomita K, Hayashi T, et al. A new rice cultivar with high eating quality, Ichihomare[J].Breeding Research,2018,20(2).
[19]Idesawa Y, Tanimoto R, Oyamada M, et al Screening and evaluation of high dietary fiber mutants in rice seeds[J].Breed Res, 2017,19(Suppl 1):48 (in Japanese).
[20]胡忠孝.中國水稻品种资源数据库建设与应用研究[D].长沙:湖南农业大学,2012.
猜你喜欢
世界热带农业信息(2021年7期)2021-08-11
好孩子画报(2021年3期)2021-04-02
农民致富之友(2020年8期)2020-05-11
农民致富之友(2019年23期)2019-08-16
农产品市场周刊(2017年35期)2017-10-25
养生大世界(2016年12期)2016-12-06
科学之友(2016年4期)2016-04-15
