
冷等离子体对作物种子处理效应的研究进展
2021-12-17 07:20岑慧芳刘华玥张蕴薇
作物杂志 2021年6期
宋 璟 岑慧芳,2 刘华玥 王 慧 张蕴薇
(1中国农业大学草业科学与技术学院,100193,北京;2山西农业大学草业学院,030801,山西晋中)
1 冷等离子体概述
在物理学中,等离子体是除固、液、气体外的物质第四态[1],根据系统温度可分为高温等离子体和低温等离子体。低温等离子体又分为热等离子体和冷等离子体,其中热和冷表征的是粒子的热力学平衡状态。热等离子体中的所有粒子温度都很高,处于局部热力学平衡状态(LTE)。而冷等离子体中电子温度相对较高,重粒子温度相对较低,处于非局部热力学平衡状态(NLTE)[2]。冷等离子体主要由离子、电子、激发态的原子分子、自由基等活性粒子及可见光、紫外线等组成,各组分比例随冷等离子体产生方式、放电电压和气源种类不同有所差异。冷等离子体可通过介质阻挡放电、辉光放电、射频放电和电晕放电等方式产生。由于冷等离子体可在不同压强下产生,据此又将其分为非常压(低气压/真空)冷等离子体和常压冷等离子体[3]。清华大学团队研发了一种广泛用于生物诱变育种的新型大气压射频辉光放电冷等离子体发生器,并将其产生的冷等离子体命名为常压室温等离子体(atmospheric and room temperature plasma,ARTP)[4]。
2 冷等离子体在种子处理上的研究进展
冷等离子体在植物上的研究应用主要集中在作物栽培领域。冷等离子体种子处理技术最早由俄罗斯科学家提出并在蔬菜种子上应用,随后各国科学家也陆续开展相关研究,但均未深入机理层面。截至目前,研究仍多集中于粮食作物、经济作物、饲料作物等种子播前处理,以及处理后种子萌发、植株生长及生理生化观测层面。处理效应与样品种类、冷等离子体的激发方式、气体源和处理时间等多个变量相关[1]。数分钟内的冷等离子体处理对植物起到更多的是激活作用,即在不改变基因序列的背景下诱导植物产生表观遗传变异。作用效应主要体现在改善种子发芽率和生长势等萌发特性,影响幼苗的生长状态、成株的表型及生理生化指标,提高产量并改善作物品质等[5]。而长时间的处理效果更倾向于物理诱变,造成基因层面的变化。由于冷等离子体源的多样性和物理效应的复杂性,对其具体效应机理仍有待进一步深入研究。
2.1 非常压冷等离子体对作物种子的处理效应
较多研究都表明非常压冷等离子体种子处理在种子萌发、作物生长及生产性能提升等方面具有正向作用。归纳后发现,较适合农业生产应用的非常压冷等离子放电方式主要为射频放电、辉光放电和介质阻挡放电 3种。射频放电冷等离子体是通过高频高压电离气体产生的,其放电能量高、范围大[6]。辉光放电属于低气压放电的一种,当粒子由激发态回到基态时,会以光的形式释放能量。非常压辉光放电冷等离子体会通过与种子表面发生互作,改变种子表面形态[7],促进种子萌发及植株生长。而介质阻挡放电则是通过击穿放电电极间的工作气体(绝缘介质)达到产生冷等离子体的目的[6]。但不同放电方式对同种种子处理效应的横向比较还未见报道。
2.1.1 对种子萌发及幼苗生长的影响 有研究采用非常压冷等离子体在粮食、蔬菜、油料及其他经济作物种子上进行了多年室内和田间试验,发现冷等离子体处理能增加小麦胚根和胚芽长度[8];提高蔬菜种子出苗率、幼苗整体均一性和强壮度;促进春播大豆出苗等。张娜等[9]发现,真空氦气介质冷等离子体处理在促进小麦种子萌发和幼苗生长方面作用明显,经处理,小麦种子发芽率、成苗率、出苗整齐度显著提高。用真空辉光放电冷等离子体处理大豆、油菜、花生和芝麻4种油料作物种子后,种子活力指数、发芽指数、幼苗茎长和根长显著增加[10]。处理紫花苜蓿种子后,种子活力指数、发芽势和发芽指数明显提高[11]。
冷等离子体还可以改善种子表面形态及湿润性。Nalwa等[12]采用低压辉光放电氧气冷等离子体处理甜椒种子后,种子表面形态发生改变,播种后幼苗特性更优。用低真空射频冷等离子体处理小扁豆、菜豆和小麦种子后含氧官能团会富集到种子表面,显著改善种子表面吸水性及湿润性,提高种子发芽速度[13]。研究[8]还发现,冷等离子体源(真空射频冷等离子体辉光放电)处理对恢复老化种子活力有一定的正向作用。老化种子初始发芽率相对越低,处理后种子活力提高幅度越大。且单、双子叶作物老化种子对处理的敏感程度不同。
2.1.2 对作物生长及生理的影响 一定剂量的非常压冷等离子体种子播前处理还会影响作物的生长、生理及产量。采用冷等离子体处理小麦种子,小麦的株高、分蘖数、穗粒数及籽粒产量显著提高[8]。短时间内随处理时间延长,小麦光合相关色素含量增加,光合速率提高,淀粉和籽粒蛋白质含量增加[14]。春播大豆种子经处理后,株高、侧根比和干重也均增加[8]。一定剂量的射频真空冷等离子体处理松果菊种子后,植株叶片中维生素C、酚酸含量及自由基清除活性大幅提高[15]。冷等离子体处理对向日葵种子参与光合途径或其调节的蛋白质表达具有一定刺激作用,处理后种子赤霉素含量提高,蛋白质出现差异性表达[16]。冷等离子体处理还能打破葡萄芽的休眠。将葡萄芽暴露于氦气介质阻挡放电冷等离子体中能打破其休眠,使芽裂更加均匀迅速,营养生长更加旺盛。对葡萄芽组织进行生化分析后发现,冷等离子体处理触发了瞬时氧化应激,进而提高了游离脯氨酸、丙二醛和过氧化氢含量,其中脯氨酸含量的提高可能是细胞结构受自由基损害减少和打破葡萄芽休眠的主要原因[17]。
综上,从非常压冷等离子体对作物种子的处理效应来看,该处理能改变种子表面的粗糙度及湿润性,影响种子萌发、植株生理生化及生产性能。非常压冷等离子体对种子产生的积极生物学作用:处理后种皮表面形成的活性氧自由基会改变种皮结构,增加种皮亲水性和通透性,使外部的水和氧迅速进入种子,促进萌发;同时,冷等离子体与细胞内带电离子互作后被吸收部分的电子振动并转化为热能,使种子内生物大分子由基态跃迁到激发态,提高种子内酶活性,改变种子萌发及生长时期的代谢水平[18]。
2.2 常压冷等离子体对作物种子的处理效应
2.2.1 对作物生长及生理的效应 在种子处理中应用较多的常压冷等离子放电方式为大气压介质阻挡放电(atmospheric pressure dielectric barrier discharge,APDBD)。对APDBD冷等离子体开展生物学效应研究[19]发现,处理后细胞膜蛋白含量发生变化,细胞破裂,同时胞内流出物会进一步被冷等离子体降解为小分子。APDBD处理后作物种子发芽率、植株生长及品质均有不同程度的提高。此外,处理还促进了非健康种子生理机能的恢复,处理不同健康程度的大豆种子后,其发芽势、生长势和产量均有所提高[20],故将该技术用于种子播前处理可能有助于减少化肥农药的使用。王敏等[21-22]采用自制的APDBD冷等离子体装置对生菜和黄瓜种子进行处理,发现种子发芽势、发芽率以及幼苗株高、茎粗、品质等指标得到提高。用大气压冷等离子体处理茄子种子后,植株表型指标与产量得到了提高[23]。常压冷等离子体还能穿透番茄种子的荚膜,降低细胞膜的相对渗透率,提高番茄幼苗根系的活力和产量,影响植株的部分植物学特性[24]。处理后番茄单果重、果长、果粗也较对照有明显提高[25]。但强度大、时间久的处理也会对种子萌发及生长发育产生副作用。研究[26]发现,1.6kV、1min内的大气压氩气冷等离子体处理能有效促进芥菜种子萌发,而2.2kV、2min内的处理则表现出对芥菜种子萌发的抑制作用。
采用常压冷等离子体短时间处理4d苗龄拟南芥幼苗后,其生长受到明显促进,植株体内活性氧、丙二醛、脯氨酸积累量提高,抗氧化酶表达水平提高,表明抗氧化防御系统在冷等离子体处理促进拟南芥幼苗生长过程中起作用[27]。随常压冷等离子体处理时间增加,豌豆幼苗期黄酮醇糖苷谱提高[28]。长春花种子受氩气APDBD冷等离子体处理后,光合色素、可溶性酚和脯氨酸含量提高,同时次生代谢过程和生物碱合成相关基因DAT的表达受到影响[29-30]。
2.2.2 在作物抗逆境胁迫中的效应 关于常压冷等离子体种子处理技术在提高作物抗逆性方面也有一定研究。高浓度铜胁迫下,大气压辉光放电冷等离子体处理小麦种子后会促使幼苗将 Cu2+从根部转运到地上部分,以减轻其对根部的毒害和对幼苗生长的抑制作用,同时提高根、苗的活力指数。但低浓度铜胁迫下,冷等离子体会促进根和苗对铜的吸收,不利于幼苗生长[31]。此外,常压冷等离子体处理对降低盐胁迫效应、提高水稻的耐盐性具有促进作用。常压冷等离子体处理能提高盐胁迫下水稻幼苗光合气体交换和叶绿素荧光程度,提高光合色素含量、抗氧化酶及次生代谢酶活性,降低细胞中Na+积累,增加K+和Ca2+含量,使细胞膨压得到恢复[32]。较低剂量的常压冷等离子体处理会对穿心莲种子蛋白质积累、抗氧化酶类活性及其同工酶的表达起到促进作用,减轻逆境胁迫下过氧化物对细胞造成的损伤,提高其应对非生物胁迫的能力[33]。
3 新型常压非平衡放电等离子体(ARTP)处理技术
3.1 ARTP概述
由表1可知,不同冷等离子体有不同特点,相较于非常压冷等离子体,常压冷等离子体处理无需创造真空环境,产生的紫外线、强电场、带电粒子等复杂效应的可控性更强。而ARTP是综合了已有常压冷等离子体相关研究背景提出的一种较优产生方式组合的常压室温冷等离子体源,它结合了常压和射频辉光放电的优点,使高纯度氦气在高频电场中放电,是在常压状态下产生的温度在 25℃~40℃之间的一种高能冷等离子体[34]。
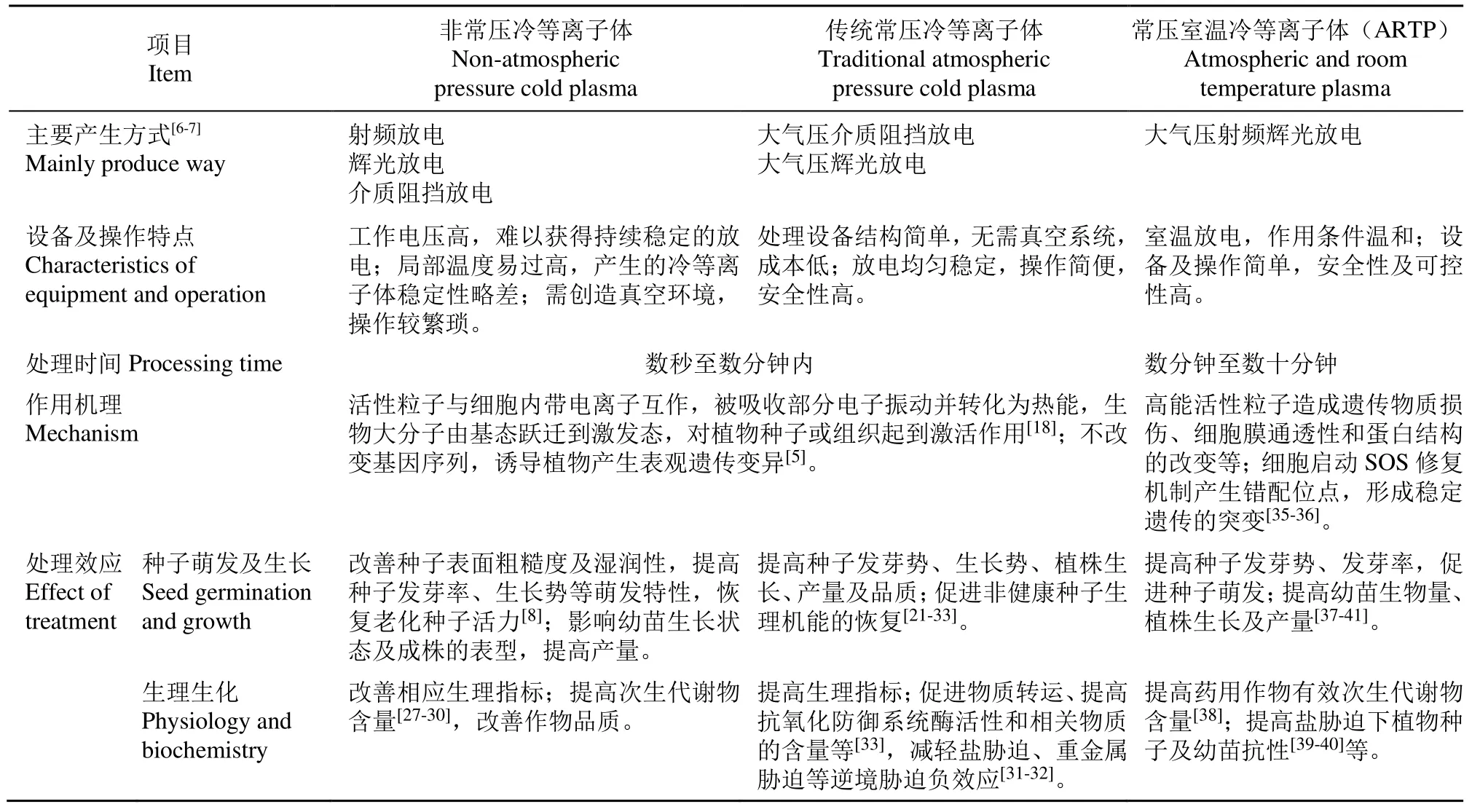
表1 不同类型冷等离子体性质及处理效应的比较Table 1 Generalization and comparison of properties and treatment effects of different cold plasma
不同于短时间非常压冷等离子体和传统常压冷等离子体处理对生物体造成的表观遗传效应,ARTP自产生以来,主要作为一种优良的物理诱变剂应用于微生物及植物诱变领域。其产生的活性粒子可透过细胞壁引起菌株或植物细胞膜通透性和蛋白结构的改变,造成遗传物质的损伤等。同时细胞启动 SOS修复机制产生种类丰富的错配位点,进一步形成能稳定遗传的基因突变[42]。故ARTP具有基因损伤强度高、诱变速度快、突变范围广等优点。其拓宽了冷等离子体的应用领域,在生物诱变育种中具有较好的应用前景。
3.2 ARTP技术在作物种子处理上的应用
近年来,ARTP正逐步被应用在原核、真核生物的诱变育种中,但已有研究大多都是围绕微生物展开的。ARTP最早应用于阿维菌素高产菌株的筛选[43],此后逐渐在多种细菌和真菌选育中推广。以促进细胞生长、增强各种次生代谢物产量、提高酶活性及增强微生物对极端条件耐受性为目的的微生物ARTP诱变已成功在生物酶、氨基酸和有机酸等药物制备业、食品制造业和能源加工业中得到应用[42]。
目前 ARTP应用在植物上的研究处于起步阶段,已有研究初步表明其会对植物种子萌发、生理生化和抗逆性等产生影响。ARTP处理玉米种子5~30min对出苗率无显著影响,但随处理时间延长,幼苗干重增加;处理30min后幼苗长度显著增加[37]。对药用植物怀牛膝和昆仑雪菊种子进行适宜剂量的ARTP处理,怀牛膝的种子发芽率、株高、最大冠幅、最大叶长、叶宽及产量有所提高,处理后昆仑雪菊种子的发芽势和幼苗生物量也有所提高,且处理后两者的次生代谢物含量也发生了变化,其中怀牛膝有效药用成分β-蜕皮甾酮含量及昆仑雪菊花朵总黄酮、木犀草苷和绿原酸含量提高[38],一定程度上表明ARTP技术具有选育高含量有效次生代谢物植株的应用潜力。ARTP处理也有助于提高盐胁迫下植物种子及幼苗的抗性。采用ARTP处理二月兰种子,可缓解盐胁迫对出苗率和根鲜重的胁迫作用,提高株高和地上部分鲜重,而未处理的对照出苗率、株高、地上和地下部分鲜重均降低[39]。有研究[40]还表明,ARTP处理通过提高多种氧化酶活性及可溶性蛋白含量、降低丙二醛含量及质膜相对透性,促进NaCl胁迫下胡麻种子发芽和幼苗生长,但对不同品种胡麻的促进效应存在差异。
ARTP的处理效应受处理剂量的直接影响,一般通过控制处理时间、功率、气流量、转速及样品距喷射口距离来调控[4]。ARTP处理在植物上的报道大多是通过控制处理时间造成处理效应的差异。如黄忠兴等[41]发现,不同 ARTP处理时间下的甘蔗胚性愈伤组织平均活愈率差异显著。此外,不同植物、同种植物不同部位的最适诱变剂量也不同,通常随诱变时间增加,处理效应呈先正向后负向的趋势。
4 展望
通过概述冷等离子体作物种子处理技术在国内外的研究成果及应用进展,发现已有研究大多围绕栽培作物展开,所处理的植物样品大多为种子。对处理效应的研究集中在表型和生理层面,对机理的探究仍有待深入。这可能是由于冷等离子体源处理具有非定向性,产生的现象是多种效应的综合结果,其机理难以从某一或某些角度单独阐述。此外,冷等离子体的产生方式、处理时间、设备及操作等具有不同的特点和倾向性,所处理的不同作物本身也存在特性差异,上述不同组合方式产生的效应难以统一化、规律化。因此,在未来冷等离子体作物处理技术发展过程中,还需结合具体研究条件和实际需要进行较多的配套应用技术研究,以期探索建立更加普适高效的冷等离子体处理体系。
相比而言,非常压和常压冷等离子体处理对作物种子萌发、生长及生理生化的整体效应无明显差异,但常压冷等离子体处理无需创造真空环境及反复添加样品,操作更为简便,在实际应用中更易推广。ARTP作为常压冷等离子体的一种,产生和作用条件更加温和,且对细胞内基因的损伤强度高于传统物理化学诱变方法。ARTP的突变位点分布广、类型多样、丰度高,是一种更加简单、高效、安全的新型冷等离子体处理技术,已经在工业微生物育种中得到了广泛应用并取得良好效果,相信未来也会在植物育种和栽培上得到更多应用。
猜你喜欢
今日农业(2021年10期)2021-11-27
中国特种设备安全(2021年5期)2021-11-06
空间科学学报(2021年6期)2021-03-09
现代塑料加工应用(2021年5期)2021-02-28
云南化工(2020年11期)2021-01-14
中国生物医学工程学报(2019年4期)2019-07-16
新农业(2017年2期)2017-11-06
材料科学与工程学报(2016年4期)2017-01-15
中国火炬(2015年12期)2015-07-31
华东理工大学学报(自然科学版)(2014年2期)2014-02-27
