
20个新选玉米自交系产量和穗部性状配合力及其相关性研究
2021-12-14 04:48罗黎明蒋辅燕高连彰和永昌汪燕芬吴海兰毕亚琪尹兴福何永健包改丽姚文华徐春霞陈洪梅
西南农业学报 2021年10期
罗黎明,蒋辅燕,高连彰,和永昌,汪燕芬,吴海兰,毕亚琪, 尹兴福,何永健,包改丽,姚文华,徐春霞,王 晶,陈洪梅*
(1.云南省农业科学院粮食作物研究所,云南 昆明 650205;2.宣威市农业技术推广中心,云南 宣威 655400;3.云南田瑞种业有限公司,云南 昆明 650205)
【研究意义】玉米是我国第一大粮食作物,因其广适、高产和粮饲兼用等特性,在我国国民经济发展和现代农业生产中居于非常重要的地位[1]。然而,近年来生产上玉米品种趋同化现象严重,突破性品种少,品种综合抗性和适应性差,究其原因主要为玉米种质基础狭窄、种质遗传基础趋同化、所选自交系的各农艺性状没有达到育种目标所致[2]。因此,扩增玉米种质,选育优良的玉米自交系是解决上述问题的关键,也是提高玉米生产稳定性的重要途径之一。【前人研究进展】育种实践和相关研究表明,在玉米自交系的选育中利用地理远缘种质互导和热带、亚热带优良种质的引入可有效扩增玉米遗传基础,提高关键性状优良基因频率,更有利于选配出优良自交系[3-6]。而配合力的测定在优良自交系的选育中起着关键作用,评价一个自交系的优劣,除了看其产量、抗性、适应性和品质等方面外,还应特别关注其配合力的高低,通过自交系配合力的测定可在育种中有效预测优良自交系的应用潜力[7-8]。番兴明等在系统研究热带亚热带玉米种质与温带种质间的配合力与杂种优势的基础上,结合育种实践,提出Suwan、Reid、非Reid的“三角形”杂种优势利用模式,并进行了大量验证[9-11]。【本研究切入点】本研究选取20个新选玉米自交系为研究对象,以分别来源于Suwan、Reid和非Reid三大类群的5个玉米自交系为测验种,采用NC-Ⅱ 遗传交配设计,对其产量及穗部性状的一般配合力(GCA)和特殊配合力(GCA)及其相关性进行分析。【拟解决的关键问题】探究20个新选玉米自交系的配合力、杂种优势及其优势组配模式,评估新选自交系在玉米育种中的应用潜力。
1 材料与方法
1.1 供试材料
20个被测玉米自交系是云南省农业科学院粮食作物研究所近年来选育的新玉米自交系;5个测验种是分别来源于Suwan、Reid和非Reid三大优势类群的自交系。自交系种质来源及主要性状见表1~2。
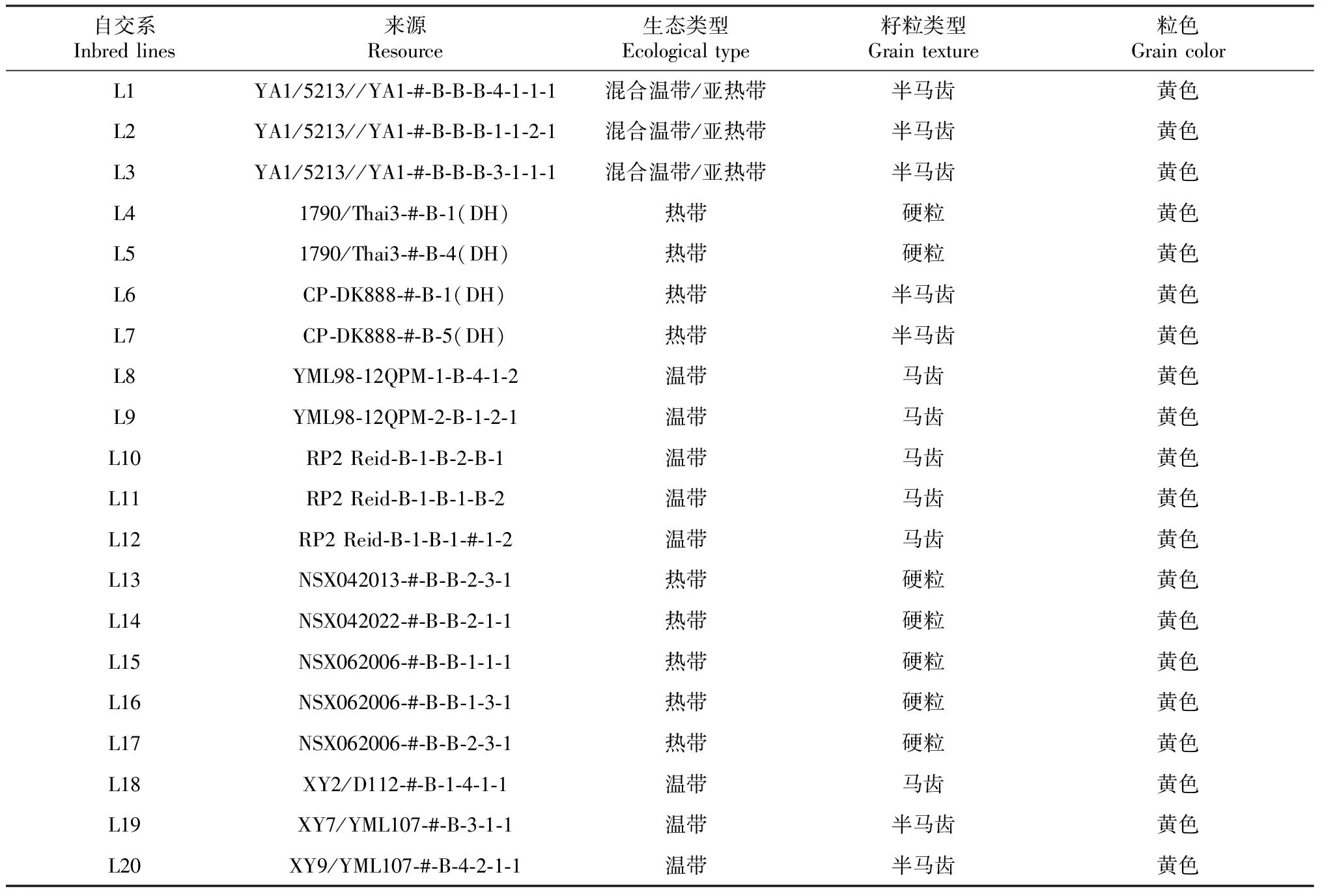
表1 20个玉米自交系的种质来源和主要性状
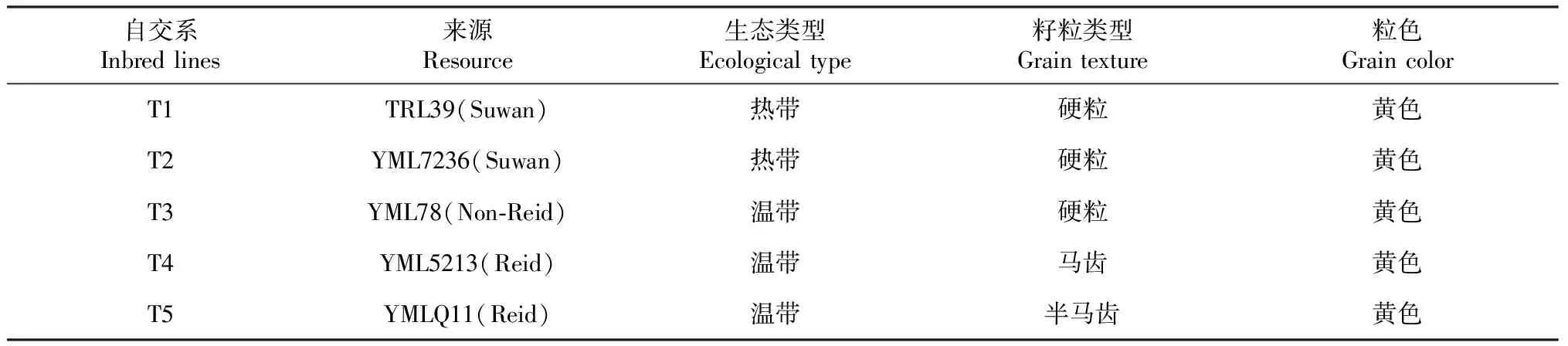
表2 5个测验种的种质来源和主要特征
1.2 试验设计
试验采用NC-Ⅱ遗传交配设计,于2017年在云南省昆明市以20个新选玉米自交系为母本、5个测验种自交系为父本,组配得到100个测交组合。2018年将这100个测交组合分别种植在云南省宣威市(海拔1980 m,北纬26°06′,东经104°03′)和砚山县江娜镇狮子山村(海拔1509 m,东经104°36′,北纬23°60′)两种不同生态环境下进行鉴定,以大面积推广品种海禾2号为对照。试验采用完全随机区组设计,2次重复,单行区,行长5 m,行距0.7 m,株距0.24 m,单株留苗,种植密度约60 000株/hm2。成熟后每小区连续收获中间10株果穗进行考种,测量穗长、穗粗、穗行数、行粒数,风干至恒重称取百粒重、单株产量,并计算出籽率。
1.3 数据统计与分析
应用SAS 9.2数据处理软件对穗长、穗粗、穗行数、行粒数、百粒重、单株产量和出籽率7个性状进行方差分析。当方差分析显著时,再用张玉东等编写的SAS line×tester程序对7个性状采用NC-Ⅱ遗传交配设计分析方法进行配合力方差分析,然后分别计算GCA和SCA的相对效应值[9]。采用SigmaPlot 12.5软件进行作图和数据相关性分析。
2 结果与分析
2.1 方差分析
对2个试验点田间试验单株产量和穗部性状进行的联合方差分析结果表明(表3),组合、被测系、测验种、被测系×测验种间各性状均达到极显著水平(P<0.01),说明各性状的组合间存在差异,可以进行下一步的配合力分析。除穗长、穗粗、穗行数和出籽率外,其余性状在不同地点间的差异达极显著水平(P<0.01),说明地点对各组合性状的表现有显著影响。

表3 100个测交组合2个试验点产量及穗部性状方差分析
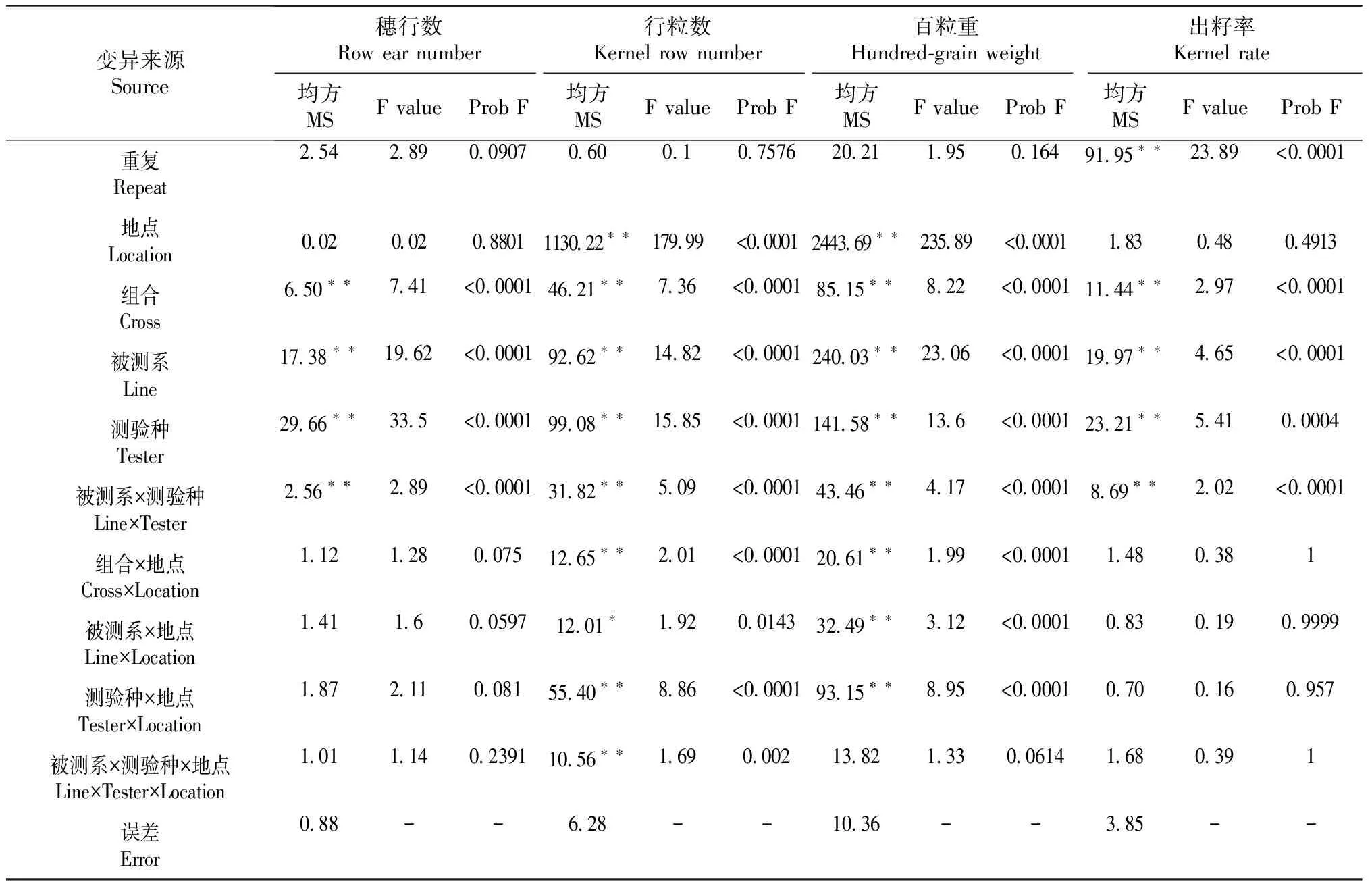
续表3 Continued table 3
2.2 配合力效应分析
2.2.1 一般配合力(GCA)分析 由图1可知,各参试自交系的产量一般配合力GCA效应值存在较大差异。其中,被测自交系L1、L2、L3、L4、L6、L7和L17为极显著或显著正值;L11、L12、L13、L16、L18和L20为极显著或显著负值;L8、L15为正值,但不显著;L5、L9、L10、L14和L19为负值,但不显著。
由表4可知,不同自交系同一性状和同一自交系不同性状的GCA效应值有很大差异。进一步分析产量GCA为极显著或显著的自交系的各穗部性状GCA效应值,自交系L1穗长、穗行数和行粒数GCA效应值为极显著正值,百粒重GCA效应值为极显著负值,穗粗和出籽率GCA效应值不显著;自交系L2除百粒重GCA效应值为极显著负值外,其余各性状均为正值,且穗粗、穗行数、行粒数和出籽率均为极显著正值;自交系L3穗行数GCA效应值为极显著正值,百粒重GCA效应值为显著负值,其余各性状均不显著;自交系L4穗长和穗行数GCA效应值为极显著正值,其余各性状均不显著;自交系L6除行数和出籽率GCA效应值为极显著负值外,其余性状均为极显著正值;自交系L7穗长、行粒数和百粒重GCA效应值为极显著或显著正值,穗粗和穗行数为极显著或显著负值;自交系L17穗行数和穗粗GCA效应值为极显著正值,穗长为极显著负值。综合分析参试自交系单株产量和穗部性状GCA,单株产量GCA效应值为极显著或显著正值的自交系,有较多穗部性状GCA效应值为极显著或显著正值,自交系L1、L2、L3、L4、L6、L7和L17的产量和穗部性状配合力综合表现优良,具有较高的育种利用潜力,在杂交育种中较易组配出性状优良的杂交组合。
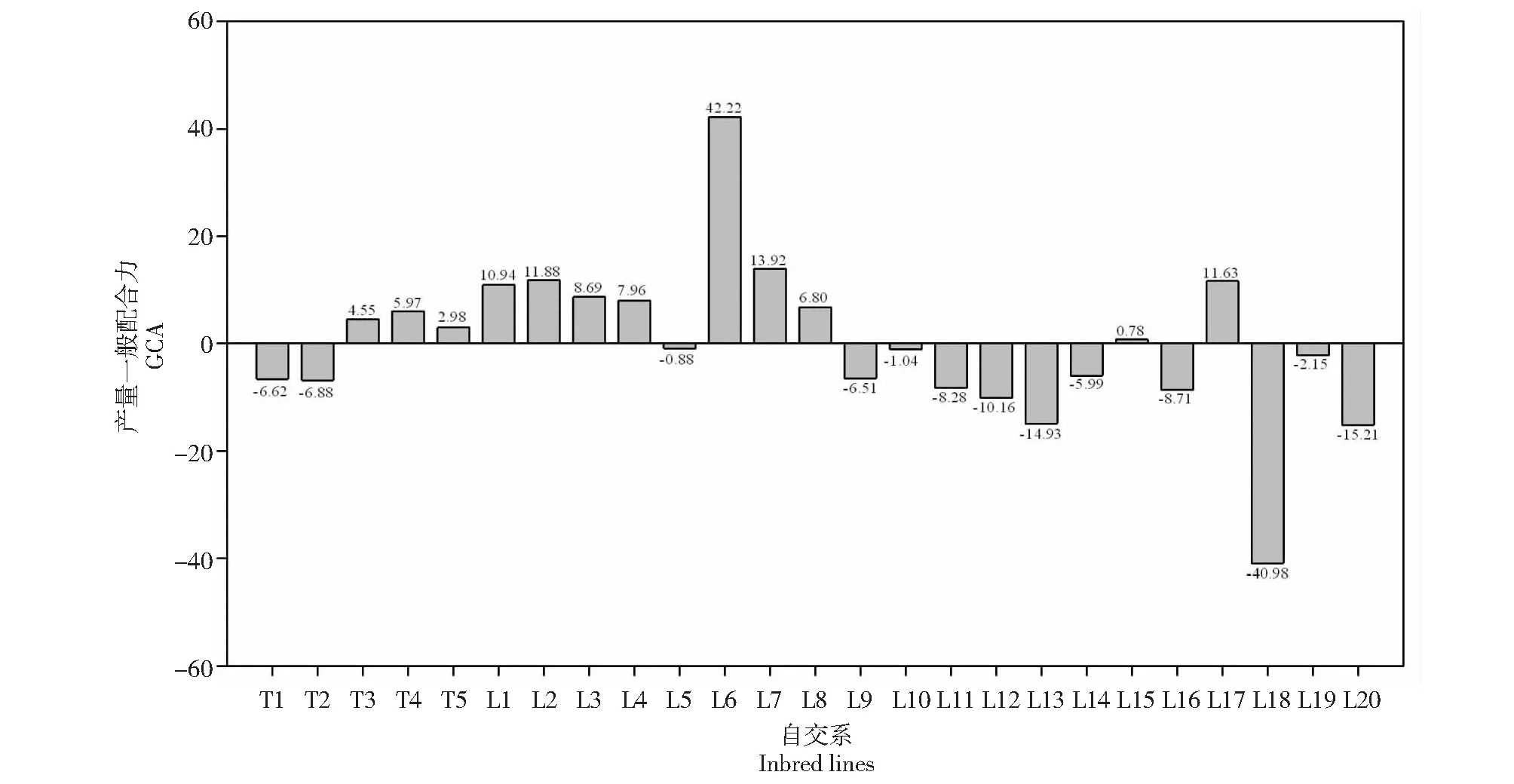
图1 20个自交系产量一般配合力(GCA)的效应值Fig.1 Estimates of GCA among 20 inbred lines for yield
2.2.2 特殊配合力(SCA)分析 由表5可知,单株产量SCA效应值为正值的组合共48个,其中有14个组合达极显著水平,6个组合达显著水平,排前10位的组合分别为:T1×L3、T5×L17、T1×L2、T3×L8、T4×L1、T4×L13、T4×L15、T1×L19、T5×L6、T5×L16,其效应值依次为83.20、48.82、48.80、46.54、45.35、42.83、40.59、37.19、34.85、32.84,这些组合大多是硬粒型×马齿型(半马齿型)或马齿型(半马齿型)×硬粒型;单株产量SCA效应值为负值的组合共52个,其中有14个组合达极显著水平,5个组合达显著水平,排后10位的组合分别为T1×L15、T1×L16、T1×L13、T4×L8、T1×L17、T4×L9、T3×L3、T3×L1、T5×L3、T4×L18,其效应值依次为-70.86、-67.05、-58.53、-57.95、-45.65、-41.64、-37.93、-37.90、-30.11、-25.11,这些组合大多是硬粒型×硬粒型或马齿型(半马齿型)×马齿型(半马齿型)。
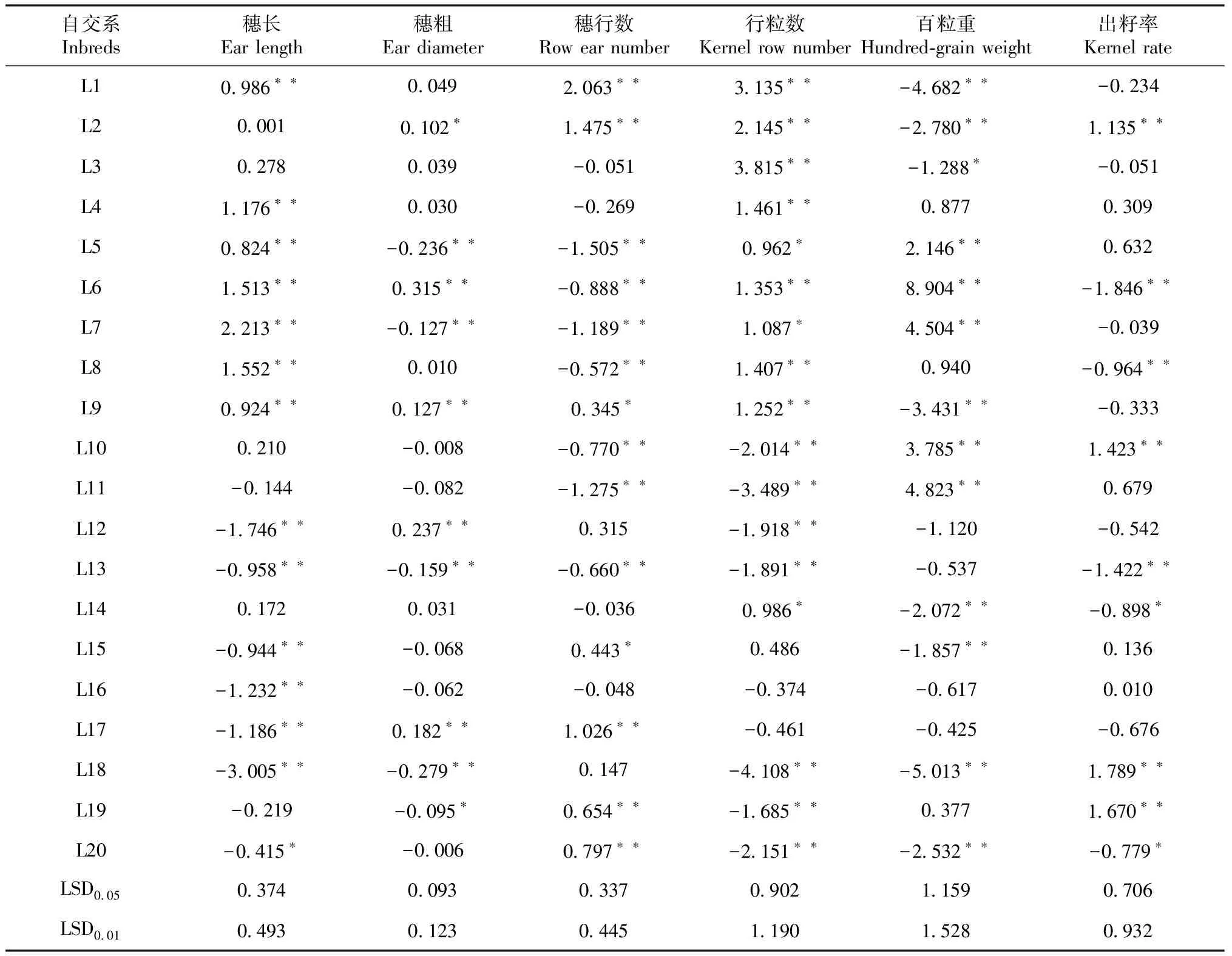
表4 20个温带被测系穗部性状的一般配合力效应值
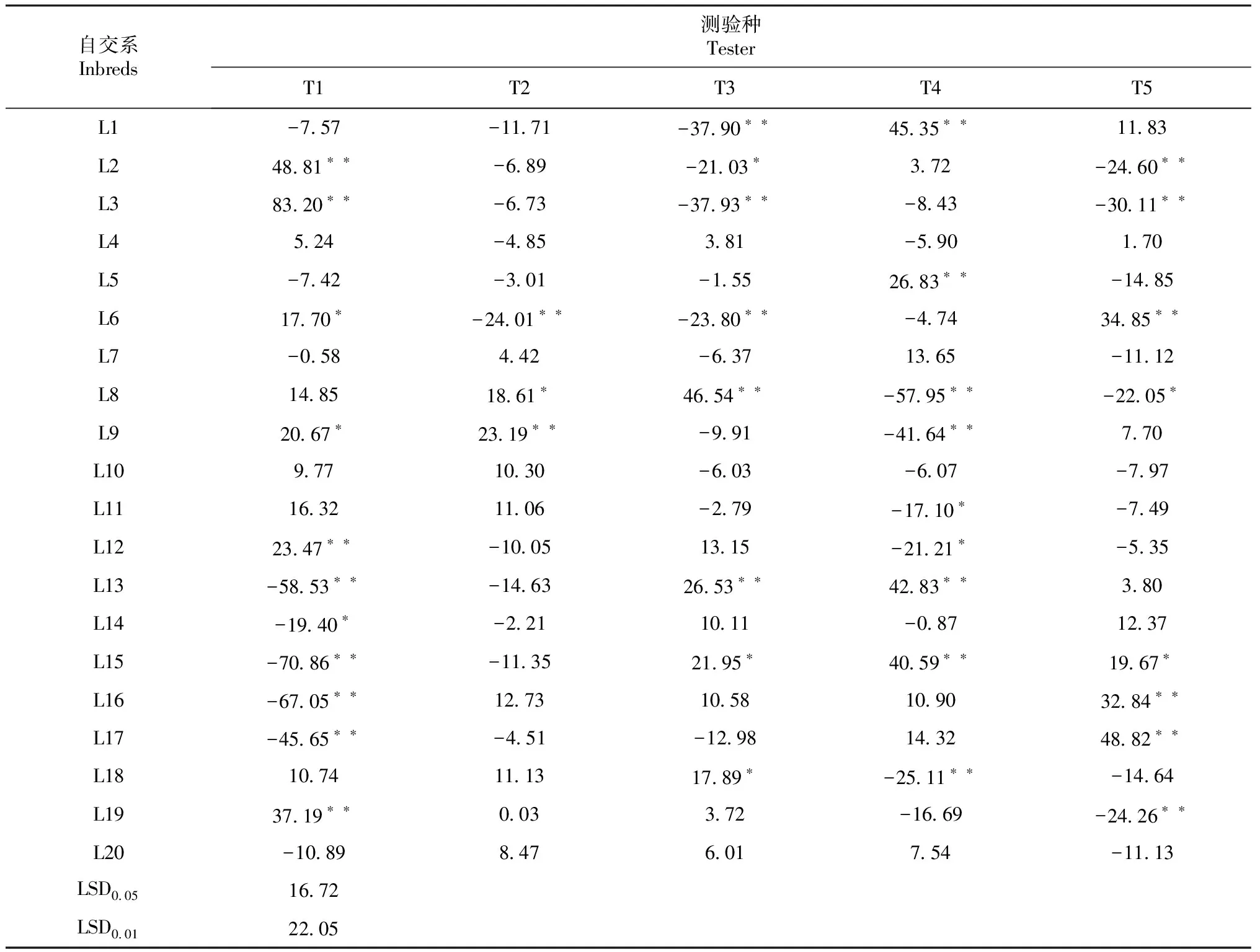
表5 100个温热杂交组合的单株产量SCA效应值
综合分析各穗部性状的SCA效应值可知(图2),同一性状不同组合以及同一组合不同性状的SCA效应值有较大差异。总体而言,单株产量SCA 效应为显著正值的组合,多数性状的SCA 效应为正值;单株产量SCA 效应为显著负值的组合,多数性状的SCA 效应为负值。
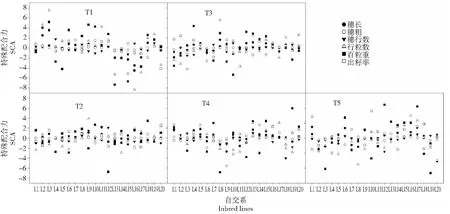
图2 20个温带被测系和5个测验种穗部性状的特殊配合力效应值Fig.2 SCA effects of ear characters among 20 temperate inbreds and 5 testers
2.2.3 对照优势分析 由表6可知,不同组合单株产量及对照优势有较大差异,单株产量变幅为87.9~246.9 g,对照优势变幅为-48.5%~44.4%。单株产量高于对照的组合有43个,其中对照优势H≥10%的组合有19个,依次为T5×L6、T1×L3、T4×L1、T3×L8、T1×L12、T1×L6、T4×L15、T4×L6、T5×L17、T4×L13、T1×L19、T3×L15、T4×L17、T4×L7、T4×L5、T3×L6、T5×L16、T5×L1、T4×L2;单株产量低于对照的组合有57个,其中对照优势H≤-20%的组合有16个,依次为T1×L16、T1×L13、T1×L15、T4×L18、T5×L18、T1×L17、T1×L18、T4×L12、T3×L18、T4×L8、T4×L9、T1×L14、T2×L13、T1×L5、T1×L20、T2×L18。
综合分析参试自交系各组合单株产量对照优势、GCA和SCA效应值可知,产量对照优势H≥10%的19个杂交组合中,多数组合双亲或亲本之一的GCA效应值为极显著或显著正值,双亲SCA效应值为极显著或显著正值的组合有13个,占68.4%。产量对照优势H≤-20%的16个组合中,多数组合双亲或亲本之一的GCA效应值为极显著或显著负值,而双亲SCA效应值为极显著或显著正值的组合有8个,仅占50.0%。综上所述,进一步说明利用产量GCA 效应值较高的自交系作亲本,较易组配出高产杂交组合;产量SCA效应值高的组合,其产量杂种优势不一定高,产量杂种优势高的组合,其产量SCA效应值也不一定高。
2.3 一般配合力(GCA)效应值相关性分析
对参试自交系穗部性状GCA效应值和单株产量GCA效应值的相关分析结果表明(图3),穗长和行粒数GCA效应值与单株产量GCA效应值呈极显著正相关,相关系数(R2)分别为0.51和0.48;穗粗和百粒重GCA效应值与单株产量GCA效应值呈显著正相关,相关系数(R2)分别为0.38和0.29,说明提高穗长、行粒数、穗粗和百粒重GCA有利于单株产量GCA的提高。本试验中穗行数和出籽率GCA与单株产量GCA之间无显著相关性,相关系数(R2)分别为0.003和0.13,说明穗行数和出籽率GCA的变化对单株产量GCA的影响不显著。
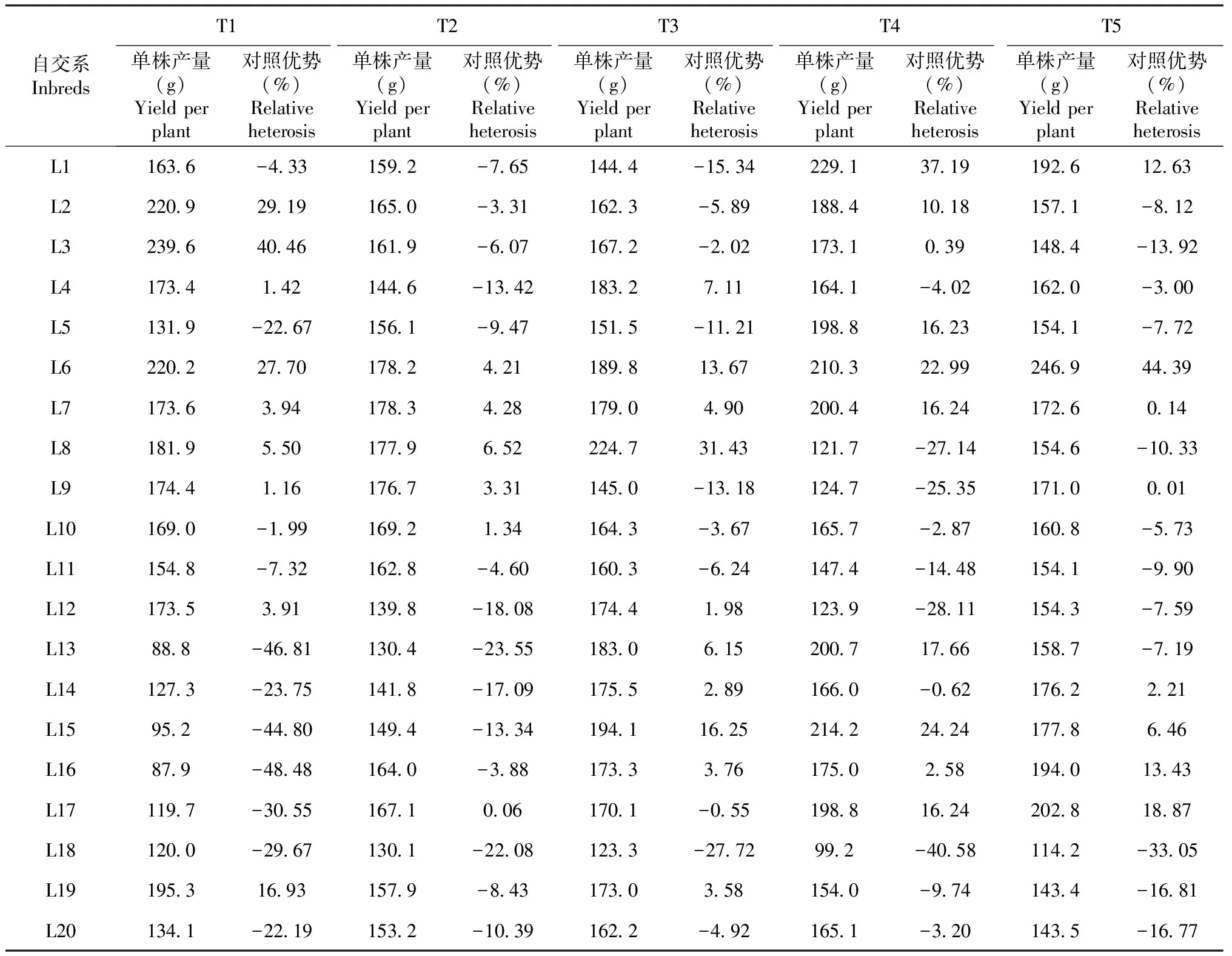
表6 100个温热杂交组合的单株产量及对照优势
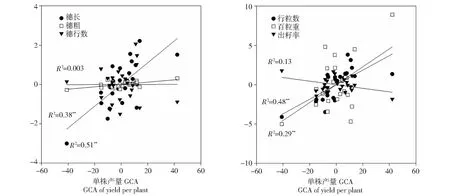
图3 20个温带被测系和5个测验种穗部性状与产量一般配合力之间的相关性Fig.3 Relationship between GCA effects of yield and ear characters among 20 temperate inbreds and 5 testers
3 讨 论
3.1 玉米自交系的配合力
玉米育种成功的关键在于选择优良的亲本自交系,但杂交后代的优劣不仅取决于自交系农艺性状的好坏,还取决于自交系的配合力高低。前人研究表明,GCA是受基因的加性效应决定且可遗传,而SCA是某一亲本与另一特定亲本组合后代的特殊表现,是由基因的非加性效应决定[12-13]。邵祺、樊丽生等[14-15]的研究指出GCA效应值在不同自交系和不同性状之间有较大差异,同一测验种与不同被测自交系产生的SCA效应值也存在较大差异,本试验对参试自交系配合力的分析也得出了同样结论。
前人研究表明GCA与SCA之间没有表现出较强的一致性,较高的GCA是自交系组合获得高产杂交后代的重要保障,同时SCA也可以为优良自交系的选育提高重要参考[16]。本研究中共试自交系L1、L2、L3、L4、L6、L7、L17具有较高的产量GCA效应值,用这些自交系组配的杂交组合大多具有较高的产量及对照优势;当两亲本的产量GCA效应值均较低或表现为负效应时,即使其SCA效应表现较好也难以获得高产后代,如本试验中自交系L12、L13、L16和L18等虽然表现出较高的产量特殊配合力SCA效应,但产量和对照优势表现均较差。而当杂交后代具有较高的产量SCA效应时,通常两个亲本中至少有一个会表现出较高的产量一般配合力GCA效应,本研究中产量SCA效应排前10的自交系组合除T1×L19外的9个组合的双亲均至少有一个具有较高的产量GCA效应,这与李明顺和张华等[17-18]的研究结果一致。
3.2 穗部性状与产量一般配合力(GCA)的关系
前人以不同自交系材料为基础,对玉米产量及其穗部性状的相关性进行了大量研究[19],指出玉米穗部各性状是形成产量的基本前提,各性状配合力的高低对产量有显著影响,产量GCA是各性状共同作用的结果[20]。本研究对玉米穗部性状与产量GCA相关性的分析表明,穗长、穗粗、行粒数和百粒重的GCA与产量GCA之间呈显著正相关关系,与祁志云和顾丽香等[21-22]的研究结果一致,而金黎明等[23]的研究则表明玉米穗粗GCA与其产量GCA之间的相关性不显著。在玉米穗部性状中穗行数GCA与产量GCA之间的相关性,前人研究结果有较大差异,阮培均等[24]研究表明穗行数GCA与产量GCA之间呈显著正相关关系,而祁志云等则得出相反的结论。本试验研究结果表明,穗行数GCA与产量GCA之间无显著相关关系,这与顾丽香等的研究结果一致。出籽率与产量GCA之间的关系在不同的研究中也得出了不同的结果,乐明艳[25]对10份白玉米自交系的研究表明出籽率与单株产量之间存在显著的正相关关系,而金黎明等的研究则显示出籽率GCA与产量GCA之间无显著相关关系。本研究结果则表明出籽率GCA与产量GCA之间无显著相关关系,与金黎明研究结果一致。综上所述,玉米不同穗部性状与产量之间的关系在不同研究中得出的结果并不完全一致,分析原因可能是所用种质材料不同造成的,说明不同种质特性的玉米自交系穗部各性状对产量的贡献及产量形成过程的调控存在较大差异。
4 结 论
新选玉米自交系L1、L2、L3、L4、L6、L7和L17的产量和穗部性状配合力综合表现优良,具有较高的育种利用潜力。供试自交系的穗长、穗粗、行粒数和百粒重与产量的GCA效应之间存在显著正相关关系,穗行数和出籽率与产量的GCA效应之间无显著相关关系,在育种中通过提高穗长、穗粗、行粒数和百粒重有利于提高产量,而穗行数和出籽率对产量的影响则需进一步探究。
猜你喜欢
中国种业(2022年8期)2022-08-19
安徽农学通报(2022年6期)2022-04-07
养殖与饲料(2021年11期)2021-11-15
种子(2021年8期)2021-09-23
园艺与种苗(2021年8期)2021-09-23
种子(2021年7期)2021-08-19
农民致富之友(2020年27期)2020-09-12
广西农学报(2019年1期)2019-07-18
农业科技与装备(2018年1期)2018-12-10
农民致富之友(2016年5期)2016-10-21
