
川滇高山栎光合特性对不同海拔梯度的响应
2021-12-11 04:36程小毛黄晓霞
西南林业大学学报 2021年6期
宁 朋 王 菲 程小毛 黄晓霞
(西南林业大学园林园艺学院,国家林业与草原局西南风景园林工程技术研究中心,云南 昆明 650233)
植物的光合特性不仅由植物自身的生理状况决定,也与光照、温度、水分条件等外界环境因子具有密切的相关性[1-3]。海拔的变化对环境因子的影响十分显著,气温、大气压以及CO2浓度均随着海拔高度的增加而降低;光照强度、紫外线辐射均随海拔高度的升高而增加[3-4]。有研究认为,高海拔地区低温、强光、低气压的环境限制了植物的光合作用;随着海拔的升高,植物叶片的光合氮利用效率(PNUE)和光合系统的氮(N)元素比例均表现出先增加后降低的变化趋势,这是由于植物为了避免随着海拔的升高而不断增强的光照对光系统的损伤而降低了叶绿素浓度和捕光组分的N元素分布比例[5-6]。因此,通过对不同海拔植物的光合特性进行比较研究,对于研究植物对于不同生态环境下的生态适应性具有重要的意义。
川滇高山栎(Quercus aquifolioide)是山毛榉科(Fagaceae)栎属(Castanea)的常绿乔木,是我国四川、云南以及西藏等省区所特有的植物,在横断山地区海拔2 400~3 900 m范围内广泛分布[7]。目前,虽然已有关于川滇高山栎的恢复能力、凋落物分解、群落结构、遗传多样性、生理生化特性、胁迫耐受性等方面的研究[8-13],但对于其光合特性随海拔梯度的变化还少见报导。因此,本研究以滇西北玉龙雪山自然保护区内的川滇高山栎为研究对象,通过比较研究其不同海拔种群的光合特性,以期理解该树种对垂直梯度小生境的生态适应,也可为该树种的天然林保护提供参考依据。
1 研究区域概况
研究区域位于云南省玉龙雪山自然保护区(东经100°10′~100°20′,北纬27°10′~27°40′)内,玉龙雪山坐落于云南省丽江市境内,其主峰位于金沙江和澜沧江之间,高达5 596 m,南面与丽江盆地接壤,西北临金沙江虎跳峡,东麓是干海子高位山间盆地,显著的海拔梯度变化和丰富的气候类型使该地区成为了当今世界上植物区系最丰富的区域[14-15]。川滇高山栎广泛分布于海拔2 750 m以上的区域内,在海拔2 900 m以上的区域内,有着东亚地区保存最完整的川滇高山栎林带,在3 500 m以上的海拔区域内,由于高山地区复杂的气候因素,川滇高山栎以斑块状分布,并逐渐向上扩展。
2 材料与方法
2.1 野外采样
川滇高山栎成片分布主要集中于海拔2 750~3 500 m,在海拔2 750 m左右由于遭受人为的砍伐而以较低矮的灌木形式存在,2 900 m为川滇高山栎、云杉(Piceaspp.)林和冷杉(Abiesspp.)林的混交林,海拔超过2 900 m达到3 050 m左右,形成了基本完整的川滇高山栎纯林林带,平均胸径可达5~10 cm,平均树高在2~8 m之间,郁闭度为0.5~0.8。川滇高山栎林在玉龙雪山分布的最高限海拔为3 500 m左右,与冷杉混生。当海拔超过3 500 m时,偶见川滇高山栎以灌木的形式零星分布。根据川滇高山栎在玉龙雪山自然保护区内的分布情况,在玉龙雪山保护区内(东经100°10′~100°20′,北纬27°10′~27°40′),沿同一坡向共设置6个海拔梯度(2 750、2 900、3 050、3 200、3 350、3 500 m)采样样地:每个样地内随机选取3个采样点,每个采样点内选择1株生长健壮、无病虫害的成年树。对树冠中部1年生枝上的健康成熟的功能叶进行光合测定,每株测不同方位的5片叶并取其平均值。待光合测定完成后再取其相应叶片用于叶面积及元素含量的测定,碳同位素组分(δ13C)的测定选取相同样株的树冠下部老叶。
2.2 指标测定
2.2.1 叶光合参数的测定
于2013年5月测定川滇高山栎光合参数。在每个样地内各选取5棵生长健壮、长势相当的高山栎植株,选取其健康、无病虫害的叶片,然后用LI−6400光合作用仪(LI-COR,美国)测定其光合作用,测定的光合参数包括胞间二氧化碳浓度(Ci)、最大净光合速率(Pn)、气孔导度(Gs)和蒸腾速率(Tr),测定时采用Led 2×3叶室,叶室内温度为25 ℃,相对湿度为36%~55%,设置CO2浓度为400 μmol/mol,光强为1 600 μmol/(m2·s)[16]。所有的指标测定都选择在晴朗天气的早上9:00—11:00完成,每株测定5片叶,每片叶重复测定3~5次,取平均值。
2.2.2 叶面积和叶干质量测定
用CI-202激光叶面积仪(CID,美国)对采摘好的植物样品叶片进行叶面积的测定,然后依次用自来水、去离子水依次洗涤,于105 ℃下杀青10 min后在65 ℃下烘干至恒质量,用电子天平(精确到0.01 g)称量叶片干质量,比叶面积用叶片的表面积与叶片干质量之比得出[17]。
2.2.3 叶片营养元素含量测定
使用FW80高速万能粉碎机(TAISITE,中国)将称量干质量后的样品叶片粉碎磨成粉末状。样品中的碳(C)、N元素含量用C、N元素分析仪(Vario macro,德国)测量。将样品用浓HNO3和H2O2在MARS Xpress微波消解仪(CEM,美国)中进行微波消解后,用ICP-OES(Perkin Elmer,美国)测量样品磷(P)元素含量[18]。最后分别计算单位叶面积的C、N、P元素含量和单位叶质量的C、N、P元素含量。
2.2.4 叶片δ13C测定
将洗净后的植物叶片置于70 ℃烘箱中48 h以上烘干至恒质量,使用FW80高速万能粉碎机粉碎样品,过200目筛后取7~8 mg封入真空的燃烧管,并加入催化剂和氧化剂,550 ℃燃烧1 h。经结晶纯化后用DeltaPlusXP质谱仪(Thermo Fisher Scientific Inc.,美国)测定植物δ13并按公式(1)计算。
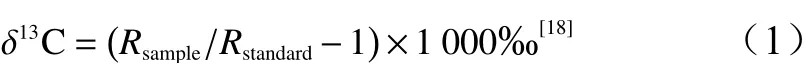
式中:Rsample和Rstandard分别为样品和标准的碳同位素比值。
2.2.5 PNUE和长期氮利用效率(NUE)计算
按公式(2)~(3)计算NUE和PNUE[19]。

2.3 数据分析
采用SPSS19.0和Excel表格进行数据的计算和统计分析,用Oringin9.0进行图表绘制,采用Duncan法进行不同数据组间的差异分析。
3 结果与分析
3.1 不同海拔川滇高山栎叶片的形态指标
由表1可知,不同海拔梯度上,川滇高山栎的叶长、叶宽、叶面积和比叶面积均存在显著差异(P<0.05),随着海拔的升高均呈现出先上升后下降的变化趋势,在海拔3 050 m处达到最大值;相比海拔3 050 m处,川滇高山栎的单叶面积在海拔3 500 m处下降了66.31%,比叶面积下降了9.47%;叶长/宽比随着海拔的升高呈现出升高—降低—升高的趋势,在海拔2 750 m和海拔3 050 m处显著低于其他海拔(P<0.05),在海拔3 500 m处达到最大值。表明随着海拔的升高,川滇高山栎叶片更加小而厚,叶形更为狭小。
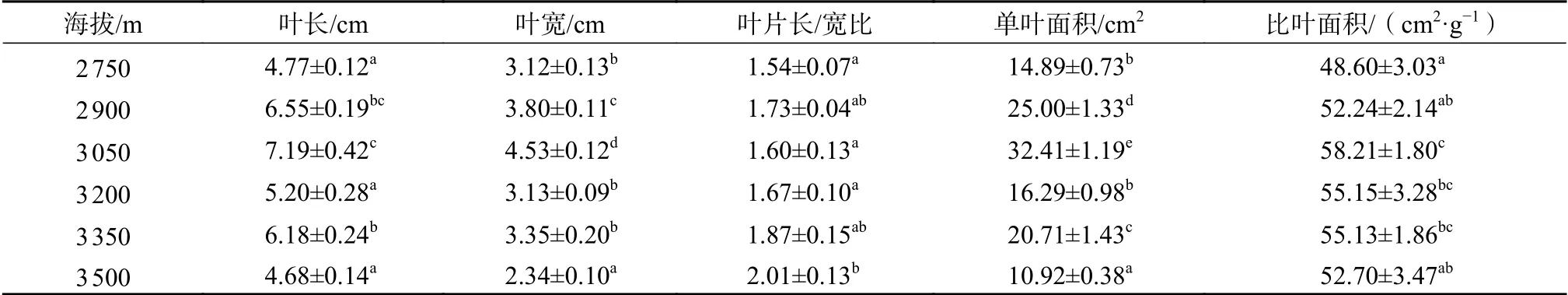
表1 不同海拔川滇高山栎的叶形态指标Table 1 Morphological indexes of Q. aquifolioides leaf at different altitudes
3.2 不同海拔川滇高山栎叶的光合参数
由图1可知,川滇高山栎的光合特性受到海拔变化的显著影响(P<0.05),随着海拔的升高,川滇高山栎的Pn、Gs、Ci、和Tr均呈现出先增加后降低的趋势。其中,Gs和Ci在海拔2 900 m处达到最大值,并且随着海拔的进一步升高呈现出下降—上升—下降的趋势,在海拔3 350 m处再次达到峰值;Pn和Tr在海拔3 050 m处达到最大值,且极显著高于其他海拔(P<0.05),随着海拔的进一步升高显著下降(P<0.05),在海拔3 500 m处分别下降了32.19%和54.82%。表明在海拔3 050 m处川滇高山栎的光合能力最强,随着海拔的进一步升高,川滇高山栎的光合能力显著下降(P<0.05)。
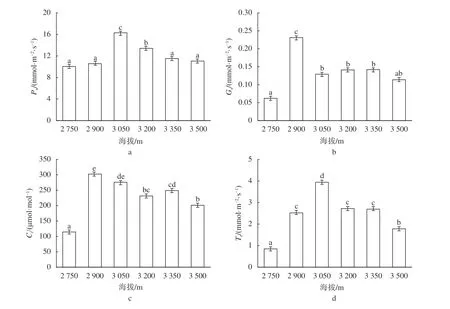
图1 不同海拔川滇高山栎叶光合参数Fig. 1 Photosynthetic parameters of Q. aquifolioides at different altitudes
3.3 不同海拔川滇高山栎叶的营养元素
由表2可知,川滇高山栎单位叶面积C、N、P元素含量均受海拔梯度的显著影响(P<0.05),单位叶面积C元素含量和单位叶面积N元素含量随海拔的升高呈现出先下降后上升的变化趋势,在海拔3 050 m处降至最低,随着海拔的进一步升高显著增加(P<0.05),在海拔3 500 m处分别增加了38.10%和27.59%;单位叶面积P元素含量随着海拔的升高呈现出先上升后下降的趋势,在海拔3 050 m处达到最高水平,随着海拔的进一步升高显著下降(P<0.05),在海拔3 500 m处下降了9.80%。
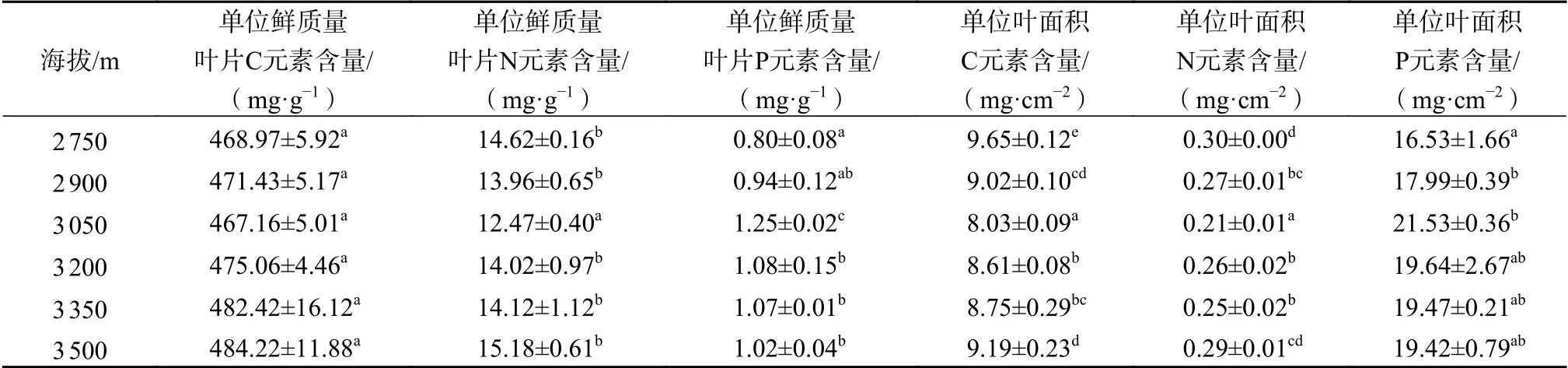
表2 不同海拔川滇高山栎叶片中营养元素含量Table 2 Contents of nutrient elements in leaves of Q. aquifolioides at different altitudes
3.4 不同海拔川滇高山栎叶片N的利用效率
由图2可知,川滇高山栎的NUE和PNUE均受到海拔梯度变化的显著影响(P<0.05),呈现出显著的非线性变化,在海拔3 050 m处达到最大值,随着海拔的进一步升高显著降低(P<0.05),在海拔3 500 m处分别下降了14.83%和49.53%,表明在海拔3 050 m区域内,川滇高山栎光合N利用水平最高效。
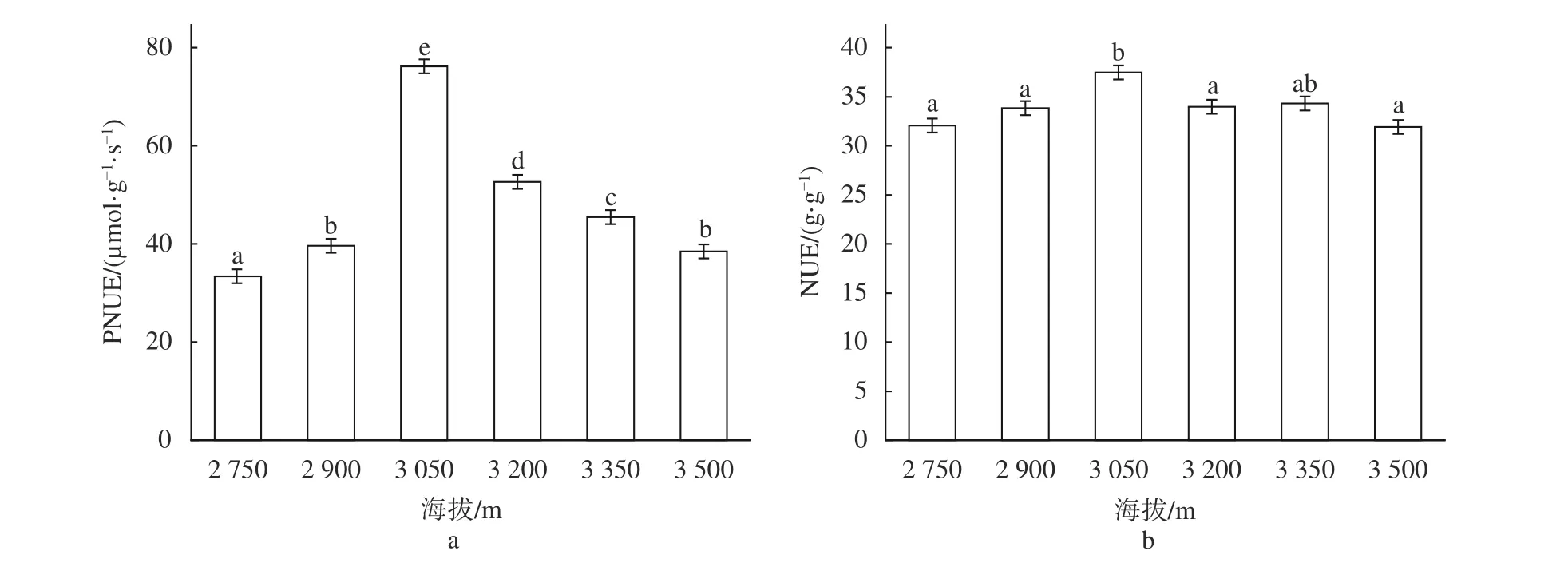
图2 不同海拔川滇高山栎的氮利用效率Fig. 2 The nitrogen utilization efficiency of Q. aquifolioides at different altitudes
3.5 不同海拔川滇高山栎叶片δ13C差异性
由图3可知,川滇高山栎的叶片δ13C值随着海拔的升高呈现出显著的非线性变化(P<0.05),在3 050 m和海拔3 200 m处显著低于其他海拔(P<0.05),然后随着海拔的进一步升高逐渐增加,在海拔3 500 m处上升了8.02%,显著高于其他海拔梯度(P<0.05),表明川滇高山栎的长期水分利用效率在海拔3 500 m处最高。
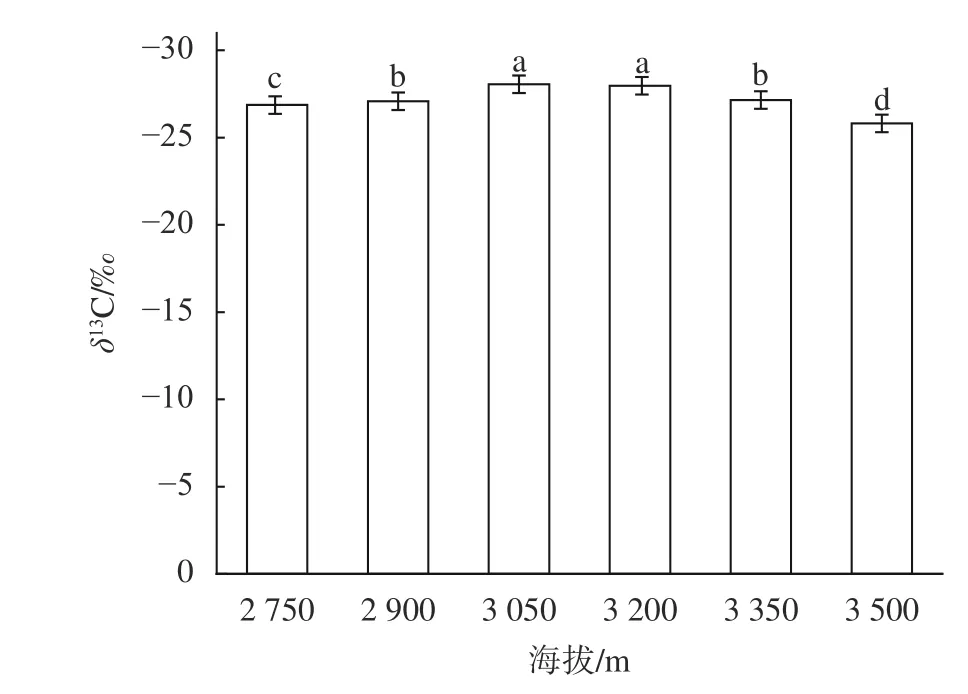
图3 不同海拔川滇高山栎叶片δ13C值Fig. 3 The δ13C value in leaves of Q. aquifolioides at different altitudes
4 结论与讨论
植物叶片的形态性状与生理特性密切相关,是植物适应环境的一种表现[20]。比叶面积是指植物叶片单面面积面积与其干质量的比值,植物在光照不足时通过提高其比叶面积以截获更多光能,在强光条件下通过减小比叶面积以增强叶片的自我保护[21],在水分亏缺的环境中通过减少比叶面积以降低Tr[22]。叶长宽比的增加有利于降低植物叶片水分蒸发和热量散失以适应低温环境[23]。有研究指出,随着海拔的升高,植物叶片的叶面积、比叶面积下降而长宽比上升,这是植物适应高海拔地区强光、低温环境的结果[23]。在本研究中,川滇高山栎的叶面积和比叶面积在海拔3 050 m处达到最大值,表明川滇高山栎在海拔3 050 m处具有更好的生态适应性;而随着海拔的继续升高,叶面积和比叶面积显著下降,而叶片长宽比显著增加,表明川滇高山栎不得不通过更加小而狭长的叶片来适应高海拔地区较为恶劣的生存环境。
高山植物的光合特性变化能够反映植物对高海拔地区环境条件的适应策略[5],Pn能有效反映植物的碳同化能力,海拔梯度的升高引起外界环境因子的变化,从而引起植物Pn的变化[5,24-25]。叶宏达等[24]认为高海拔地区的低温和强光环境有利于马铃薯(Solanum tuberosum)Pn的增加,金高明[25]在对富士苹果(Malus domestica)的研究中也得出了类似的结论,而李惠梅等[5]则认为,高海拔地区的低温、强光、低气压等不利环境条件降低了植物的Pn。在本研究中,生长在海拔2 750~3 050 m的区域内川滇高山栎的Pn随着海拔的升高逐渐增加,可能是由于光照强度随着海拔的升高而增强,使叶片能吸收到更多的光能[24-25];而随着海拔的进一步升高,高海拔地区的环境更为恶劣,降低了川滇高山栎的Pn和Tr[5]。植物光合能力主要受气孔因素和非气孔因素两方面的限制[26]:气孔限制表现为Gs降低而使Ci下降[27];非气孔限制则是由光合器官破坏、光合酶活性下降、光合色素含量降低、活性氧代谢功能破坏等引起的Pn值下降,而Ci上升或不变[26,28]。本研究中,在海拔3 050 m处,川滇高山栎的Pn主要受到气孔限制,在海拔3 200~3 350 m的范围内,Pn下降而Ci上升,表明此时川滇高山栎的Pn主要受到非气孔因素的限制。这可能是由高海拔地区缺水、高UV-B辐射的自然环境引起的[28-29]。
叶片是植物代谢活动最活跃的器官,叶片营养特征可以反映植物的营养利用策略,而研究叶的营养含量对叶营养特征的研究具有重要的意义[30-31]。张志录等[32]认为,叶片C、N含量随海拔而升高可能与叶片的光保护有关。刘千里等[33]对川滇高山栎的研究中指出,川滇高山栎成年叶中N元素含量随着海拔的变化呈现出先降低后增加的趋势。而卢月娅[34]则认为随着海拔的升高,光照增强,植物不得不降低叶绿素的含量来缓解强光对光合系统的伤害,进而导致叶片中N元素含量减少。在本研究中,川滇高山栎单位叶面积C、N元素含量在海拔3 050 m处达到最低后又随海拔的升高而增加,这可能是川滇高山栎适应高海拔地区光照条件的方式。P元素是酶类的重要组成部分,其含量可以有效反映植物生长健康状况,而低温和缺水环境会降低土壤中的有效P含量,抑制植物对P的吸收[35]。本研究中结果表明,在海拔3 050 m生长的川滇高山栎能够更好地吸收利用土壤中的P元素,具有更好的生长健康状况,这可能是由于此处具有更好的温度和水分条件,而随着海拔的进一步升高,高海拔地区低温和缺水的恶劣环境使得川滇高山栎对P元素的吸收受到抑制,也降低了川滇高山栎的生长健康状况。
N元素是组成光合器官的重要元素,因此叶片光合能力受到N元素含量的直接影响,叶片PNUE是叶片的单位面Pn值与单位叶面积N元素含量的比值,可以反映植物N元素投资于光合作用的水平[36]。有研究指出,高海拔地区的植物为缓解强光对光合器官造成的损伤,会减少N对于光合系统的投资,而将更多N投资于保护构造上,表现为植物PNUE随着海拔的升高而降低[6]。在本研究中,在海拔3 050 m处,川滇高山栎的PNUE达到最大值,说明在海拔3 050 m处,川滇高山栎将更多的N元素投资于光合作用;而随着海拔的进一步升高,环境胁迫愈加严重,迫使川滇高山栎将更多的N元素投资于保护组织,从而使PNUE降低,这也是川滇高山栎适应高海拔地区胁迫环境的一种表现。
叶片的δ13C值与植物的长期水分利用率存在正相关关系,可以有效反映植物的长期水分利用率。有研究指出,植物叶片的δ13C值与土壤温度和土壤水分含量呈负相关,高海拔地区的低温和缺水环境引起了植物叶片δ13C值的增加[37-38]。在本研究中,川滇高山栎的δ13C值在海拔3 050 m和3 200 m的海拔区域内降至最低值,说明海拔3 050 m和3 200 m处具有更好的水分、温度及光照条件,川滇高山栎光合碳同化能力较强,生长较快,而蒸腾耗水也较强,水分利用效率低下。而随着海拔的进一步升高,高海拔地区的低温、干旱以及紫外辐射的增加等都可能降低叶片的光合能力,使得叶片生长狭小,生长缓慢,但同时也可以提高其对水分的利用效率,如本研究中在高海拔3 500 m处,川滇高山栎的叶片δ13C值均高于其他海拔。
本研究表明,在海拔3 050 m处,川滇高山栎的单叶面积、比叶面积、Pn、N利用效率均达到最大值,水分利用效率最低,说明海拔3 050 m处是川滇高山栎的最适生长区;而随着海拔的进一步升高,川滇高山栎Pn、N利用效率、比叶面积显著下降,叶长/宽比显著增加,叶呈狭长型,水分利用效率显著增加,表明川滇高山栎可以通过积极改变其叶片生长形态及生理生化特性来适应高海拔的不良环境。
猜你喜欢
农学学报(2022年8期)2022-11-16
灌溉排水学报(2022年9期)2022-09-28
农业与技术(2022年16期)2022-09-01
军事文摘(2022年14期)2022-08-26
农业工程学报(2022年8期)2022-08-08
作物学报(2022年10期)2022-07-21
散文诗(2021年22期)2022-01-12
环球时报(2019-05-23)2019-05-23
物联网技术(2016年11期)2017-01-12
江苏农业科学(2016年9期)2016-11-28
