
生长季封育和氮添加对退化高寒沼泽湿地土壤水分状况的影响
2021-12-09 09:05李宏林胡竞格梁德飞周华坤
草地学报 2021年11期
李宏林, 胡竞格, 肖 锋, 梁德飞, 周华坤
(1.青海大学省部共建三江源生态与高原农牧业国家重点实验室, 青海 西宁 810016; 2. 青海省寒区恢复生态学重点实验室,中国 科学院西北高原生物研究所, 青海 西宁 810008; 3. 青海省林草局, 青海 西宁 810002)
青藏高原高寒湿地是高寒草地生态系统的重要组成,既是我国重要的湿地分布区,也是全球气候变化的敏感区[1-3]。高寒湿地具有水源涵养和补给,碳储存,优良牧草供给等重要的生态和经济意义,是我国乃至亚洲重要生态安全屏障的主要组成部分[4]。然而,近几十年来在气候变化和人类活动影响日趋加剧的情形下,高寒湿地正在面临着快速退化和生态功能丧失的威胁,表现为湿地面积大幅度萎缩,土壤含水量和地表水持续时间明显下降,土壤理化性质和微生物组成显著变化,植物群落组成和结构明显改变[5-9]。
土壤水分作为高寒湿地最重要的环境要素和控制因子,是决定湿地生态系统结构功能和生态过程的关键[10-11]。研究表明,高寒湿地在退化过程中,土壤含水量持续减少,导致地上植物群落结构组成和土壤理化和微生物特征发生明显变化,进而影响到生态系统的服务功能。因此对于高寒湿地的恢复,最关键的问题是恢复土壤水分[12],通过不同的恢复措施,增加湿地土壤水分,从而促进湿地原生植被的重建,进而恢复高寒湿地的生态系统服务功能。
一般而言,湿地生态系统的恢复包括主动恢复(以人工辅助手段为主)和被动恢复(自然恢复为主)两种方法[13-14]。例如通过构筑水坝拦截水流,增加蓄水并扩大退化湿地过水面积,或者通过添加工业修复材料,改变退化湿地土壤物理性质,增加土壤水分,促进退化湿地土壤和植被重建的主动恢复[15-16];再如通过引进栽培牧草,在退化湿地中构建人工或半人工草地,迅速提高地表覆盖度,进而实现退化湿地水文条件改善和功能提升[17],或者利用土壤种子库作为退化湿地植物自然恢复演替的种子来源[18],通过植被恢复提高土壤水分状况,进而实现受损湿地生态系统的修复等被动恢复。虽然主动恢复具有恢复速度快、可控性高等特点,但是其对自然景观和生态系统的完整性的破坏也较大,因而许多研究更加关注通过适度的人工辅助,主要依靠自然恢复为主的被动修复方法,如围栏封育和营养元素添加等方法[19]。
近年来,以适度围栏封育和营养元素添加单独/协同来恢复受损草地生态系统的方法在退化高寒草地中被广泛应用,并取得了较好的恢复效果[20-24],例如返青期休牧促进了高寒草甸植物群落的生产力和多样性[25]。然而目前有关围栏封育和施肥的研究对高寒湿地恢复影响的研究则相对较少[16]。本研究通过在青藏高原退化高寒湿地生态系统进行生长季封育和施肥试验,评价围栏封育和氮添加对退化高寒湿地土壤水分状况的影响,以探究围栏封育和氮添加在退化高寒湿地恢复中的作用及其机制,以期为气候变化背景下高寒湿地的保护、管理和恢复提供科学依据。
1 材料与方法
1.1 研究区概况
本研究在兰州大学高寒草甸与生态系统定位研究站(玛曲)高寒湿地实验样地内进行。试验地点位于甘肃省玛曲县境内,地处青藏高原东缘,平均海拔3 650 m,地理坐标为33°39′ N,101°52′ E,年平均气温为2.2℃年降水量为672 mm,属于半湿润半干旱气候,年日照时数约2 580 h,年均无霜期少于100 d[6,26]。
1.2 试验设计
于2018年5月,在高寒湿地试验区内选择退化程度较为一致的样地,该样地位于典型湿地区外围,属于轻度退化的高寒湿地,在暴雨或连续降水后有短暂的地面积水,其物种组成和水文特征都与未退化的典型湿地有显著差异。有关试验区湿地植被及其退化状况可参考Ma[27]和李宏林等[6]的描述。
首先进行裂区样方设计,在湿地中相隔20 m设置两个不同的放牧方式样区:生长季禁牧(Winter grazing,WG)和全年放牧(Year-round grazing,RG)。在每种放牧样区内,采用随机区组设计原则,设置不同的施肥梯度样方:CK(0 gN·m-2),LN(5 gN·m-2)和HN(10 gN·m-2)。每个施肥处理设置6个重复,两个不同放牧方式样区内各18个样方,试验总共36个样方。生长季禁牧样区采用普通铁丝围栏进行围封(高1.5 m,立柱间隔10 m,顶端置刺丝),从2018年开始(连续3年),在每年10月至第二年4月进行放牧,5月初封闭,全年放牧区整体处于中重度放牧状态,牧草利用率约为50%~70%。施肥于每年5月中下旬的阴雨天进行,所用肥料为尿素[CO(NH2)2]。
1.3 数据采集
于每年8月中旬开展样方调查,具体方法为:在每个处理的每个重复样区内随机选择一个50 cm×50 cm的样方,记录样方内出现的每个物种,得到群落物种丰富度(Species richnes,SR);然后采用网格法估计样方中每个物种的盖度(Coverage,%);最后剪取样方内所有植物的地上部分,装袋带回实验室烘干至恒重并称重,获取群落净初级生产力(Net primary productivity,NPP,g·m-2)。
在群落调查结束之后,在每个样方内用土钻(直径为3.5 cm)钻取0~20 cm土壤样品2份,装自封袋带回实验室,用于土壤理化分析。具体分析方法如下[28]:带回的土壤混合后用2 M的KCl溶液提取后在流动注射分析仪(AA3,Seal,Germany)上测定土壤速效氮含量(Available nitrogen,AN,mg·kg-1);剩余的土样风干,研磨并过0.2 mm土壤筛后,先使用硫酸高温消煮,然后在流动注射分析仪上测定土壤全磷含量(Total phosphorus,TP,mg·g-1)。土壤含水量于每年生长季使用TDR土壤水分速测仪按照3~4 d一次的间隔进行测定,对每个样区中心位置随机选择3个点进行测量,获取土壤含水量(Soil water content,SWC,%)。
1.4 数据分析方法
采用SPSS 16.0软件应用方差分析(ANOVA)对群落净初级生产力、物种丰富度、盖度、速效氮含量、全磷含量,以及土壤含水量等群落植被和土壤理化指标对不同放牧方式和氮添加水平的响应进行分析,并使用Origin 8.0软件对结果进行绘图。
2 结果与分析
2.1 土壤水分状况和营养元素对生长季节禁牧和氮添加的响应
生长季禁牧(WG)显著提高了退化高寒湿地土壤含水量,土壤中的速效氮和土壤全磷含量(图1a,b,c,P<0.05)。在禁牧处理下,低氮(LN)和高氮(HN)添加处理下的土壤含水量都显著高于对照(CK)(图1a,P<0.05);无论在WG还是全年放牧处理(YG)中,随着氮添加浓度的升高,土壤中的速效氮含量升高,全磷含量降低,但是土壤速效氮含量在不同氮添加处理之间差异不显著(图1b),而土壤全磷含量则在HN处理下显著低于CK处理(图1c)。
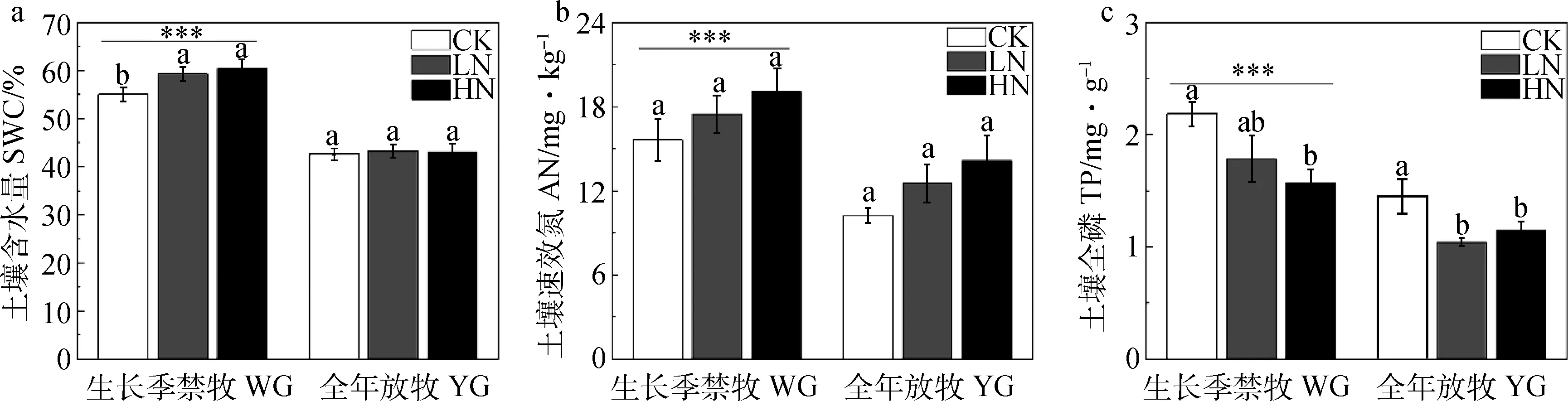
图1 放牧方式和氮添加对退化高寒湿地土壤含水量(a)、土壤速效氮(b)和土壤全磷(c)含量的影响Fig.1 Effects of grazing regime and nitrogen addition on soil water content (SWC,a),soil available nitrogen (AN,b), and soil total phosphorus (TP,c)注:CK表示不施肥;LN为施肥5 g N·m-2;HN为施肥 10 g N·m-2。星号表示不同放牧方式之间差异显著(***:P < 0.01;**:P<0.05);不同小写字母表示在同一放牧方式下不同施肥处理之间差异显著(P < 0.05)。下同Note:CK represents no nitrogen addition,LN represents nitrogen addition of 5 g N·m-2,HN represents nitrogen addition of 10 g N·m-2. Asterisk indicates significant difference between different grazing regime. The different lowercase letters indicate significant differences between nitrogen treatments under same grazing regime. The same as below
2.2 植物群落对生长季禁牧和氮添加的响应
生长季禁牧显著提高了植物群落地上植被净初级生产(NPP),同时在WG处理下,两种浓度的氮添加也显著提高了NPP(图2a,P<0.05),但是在YG处理下,氮添加并没有显著提高NPP。
群落物种丰富度(SR)在两种放牧方式之间没有显著差异(图2b),且在两种放牧方式下不同氮添加处理之间也没有显著差异(图2b)。
虽然不同放牧方式和氮添加对群落SR没有显著影响,但是禁牧显著提高了禾本科植物的盖度,降低了杂类草植物的盖度(图3a,b)。同时禁牧和氮添加也对群落中优势物种的盖度有显著的影响(图4),例如,发草和乌拉薹草的盖度在禁牧施肥处理下显著高于放牧未施肥处理(图4a,b,P<0.05),而鹅绒委陵菜的盖度则表现出完全相反的趋势(图4c,P<0.01)。
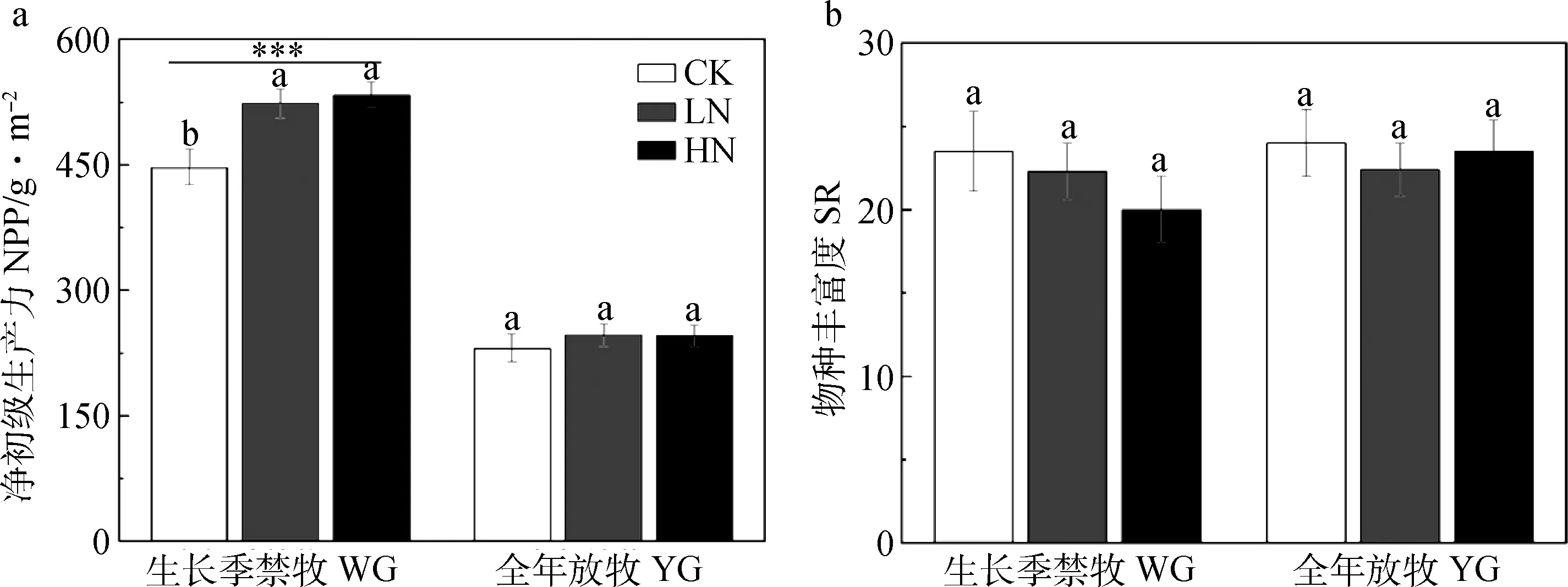
图2 放牧方式和氮添加对退化高寒湿地群落净初级生产力(a) 和物种丰富度(b)的影响Fig.2 Effects of grazing regime and nitrogen addition on community net primary productivity (NPP,a) and species richness (SR,b)

图3 放牧方式和氮添加对退化高寒湿地不同功能群物种丰富度的影响Fig.3 Effects of grazing regime and nitrogen addition on species richness of different functional group
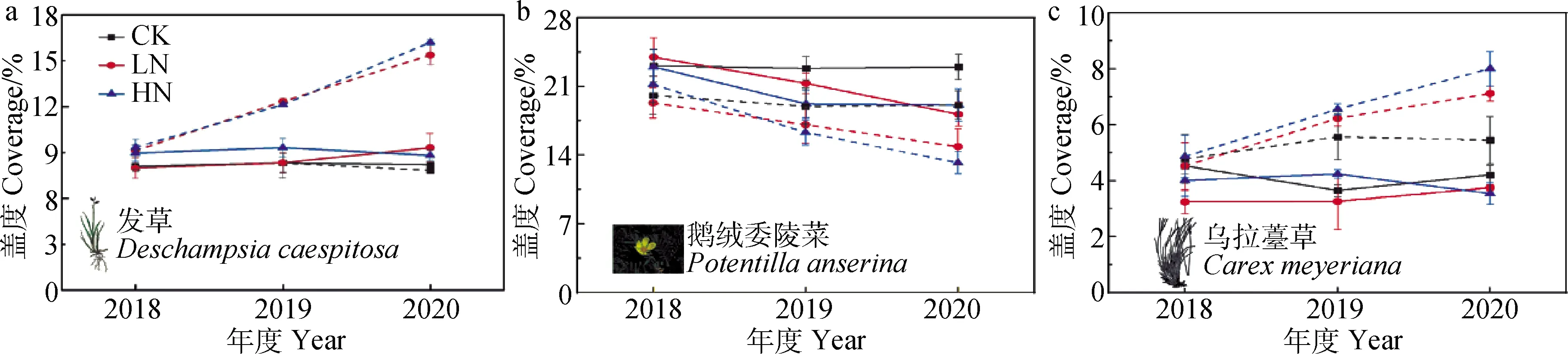
图4 放牧方式和氮添加对退化高寒湿地群落优势种盖度的影响Fig.4 Effects of grazing regime and nitrogen addition on coverage of dominant species注:实线为全年放牧处理,虚线为生长季禁牧处理Note:Solid lines represent year-around grazing treatment (YG),dashed lines represent growing season fencing treatment (WG)
2.3 生长季禁牧和氮添加对退化高寒湿地土壤水分状况影响的概念模型
基于前期的研究[6]和本试验的结果,我们构建了一个放牧方式和营养元素添加可能对退化高寒湿地土壤水分影响的简单概念模型(Framework model,图5)。由图可知,由生长季禁牧引起的退化高寒湿地植物群落结构(物种组成和不同物种的盖度)的改变是导致退化高寒湿地土壤含水量提高、土壤水分状况向好变化的关键。低浓度氮添加可能会通过抑制杂类草物种的丰富度和盖度进而促进土壤水分的增加,但是过高的氮添加则可能会由于极大地提高植物群落NPP而导致土壤水分的过度消耗。
3 讨论
研究发现[6,29],随着高寒湿地退化,湿地中原有优势种(包括莎草科和禾本科物种)的盖度和生物量在群落中的占比下降,而杂类草的盖度和生物量占比上升,造成湿地的水源涵养能力、初级生产力和生物多样性维持等生态系统功能出现明显下降。在退化湿地的恢复过程中,土壤含水量的增加、植被覆盖度的提高、原生优势物种比例的上升,以及土壤可利用营养元素的增多都是非常重要的指示性因子[17],而在这中间,土壤水分含量的提高尤其重要[6,10-11,17]。在本研究中,通过在一个轻中度退化的高寒湿地中开展为期3年的围栏封育和施肥试验,监测了生长季禁牧和氮元素添加在较短时间尺度上对湿地土壤理化性质和植物群落功能结构的影响,进而基于土壤含水量的变化评价了这些处理措施对湿地恢复的作用,构建了一个用于解释其可能机制的概念模型。
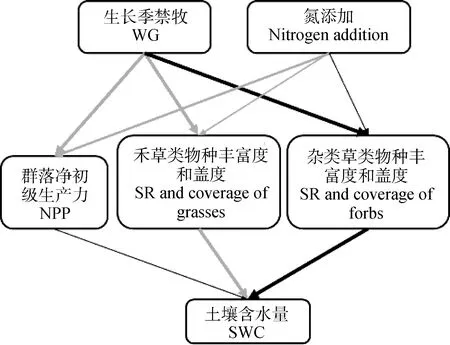
图5 生长季禁牧和氮添加对退化高寒湿地土壤 水分含量影响的简单概念模型Fig.5 Simple frame model explained the effects of grazing regime and nitrogen addition on soil water content of degraded alpine wetland注:灰色线表示具有正效应,黑色线表示具有负效应;线的粗细表示影响的强弱Note:Gray lines indicate positive effect;dark lines indicate negative effect;the thickness of lines indicates the degree to which the effect is
本研究发现生长季禁牧显著提高了退化湿地土壤水分含量(P<0.05)。虽然不同施肥处理之间土壤水分含量差异不显著,但是本研究发现土壤含水量在施肥处理下也具有增加的趋势(图1)。说明生长季禁牧和施肥的交互作用可以有效提高退化高寒湿地土壤含水量,改善退化湿地的土壤水分状况,从而为湿地的进一步恢复提供基础[30]。此外生长季禁牧和氮添加还提高了土壤中AN的含量,这一方面是由于无机氮的直接添加引起的,另一方面可能是由于土壤含水量的提高促进了湿地中固氮效应[31-32],从而提高了禁牧湿地中的土壤AN含量。研究发现,施氮肥能够显著降低土壤TP含量,而且这种抑制作用还可能会随着氮添加浓度的升高而加强[33-34],本研究的结果与之相似,说明在高寒湿地群落中氮添加也会降低土壤全磷含量。本研究中禁牧处理下TP含量显著高于完全放牧条件则说明土壤水分以及植物生长状况可能会对土壤TP的含量产生影响[35]。
在植物群落特征方面,与其它在高寒退化草地开展的研究的结果较为一致[19,22,25,36-37],例如生长季禁牧和氮添加均明显提高了退化高湿地群落NPP,增加了禾本科和莎草科植物在群落中的占比,降低了杂类草植物的比例。总体上,禁牧处理和氮添加对退化湿地植物群落影响较为一致,但生长季禁牧的效果都更加明显[16-17],这表明过度放牧是高寒湿地退化最普遍和最重要的影响因素,而适度禁牧可以作为湿地恢复的一种有效方式,同时这也预示着在湿地退化背景下继续放牧可能会加剧这种退化趋势。以往的研究更多强调湿地生态系统演变和退化中气候变化的重要作用[1,12,38],本研究结果则暗示合理放牧对保护高寒湿地生态系统可能具有同样重要的意义[21,39]。
在本区域开展的研究还发现植物水分利用效率在不同物种间差异巨大[6],因此植物群落物种组成的不同,或者同一物种在群落中比例的变化都会改变群落水平的水分利用效率,进而影响土壤水分状况。例如分别在中国[40]和美国[41]湿地中开展的研究均发现湿地植物群落由于退化或者生物入侵导致的物种组成改变,会造成土壤中水分状况的变化。而更进一步的研究还表明水分含量的减少可能会进一步对地上群落的结构和组成产生负反馈效应[6]。在本研究中,生长季禁牧和施肥条件下,由于水分利用效率更高的禾本科和莎草科植物占比增加,而同时水分利用效率较低的杂类草占比降低[6,42],使得封育施肥后群落通过植物蒸腾作用散失到空气中的水分相对减少,再加上由于封育施肥地植被较高的生物量和覆盖度能够使土壤蓄留和保存更多的水分[38],从而使土壤含水量显著提高。
基于上述研究结果和前期研究的结论[6,42],我们提出一个基于湿地恢复过程的概念模型,以初步解释生长季封育和氮添加提高退化草地土壤含水量的可能机制。该概念模型主要明确以下两点:1)生长季封育和施肥显著增加群落地上植被盖度,减少了裸地比例,使得土壤水分主要通过植物蒸腾作用进入到空气,减少了水分通过蒸发的方式耗散,从而提高了土壤含水量;2)生长季封育和施肥显著提高了群落中禾本科植物和莎草科植物的盖度,尤其是发草和乌拉苔草的盖度,这两种植物都属于水分利用效率高的湿地原生物种[6,42],这在一定程度上提高了群落水平的水分利用效率,减少了土壤水分的丧失。
4 结论
生长季禁牧和氮添加能够显著提高退化高寒湿地土壤水分含量,这主要是由于生长季禁牧和氮添加提高了退化湿地群落覆盖度、增加了群落中水分利用效率高的莎草科和禾本科物种占比,从而整体上减小了退化湿地土壤水分的散失。而退化湿地土壤水分含量的增加则可能有利于湿地原生物种的生长和繁殖,从而促进退化湿地的进一步恢复和原生植被的重建。本研究作为前期研究的延伸,后续还需要开展更长时间的观测和更加细致的野外和室内试验,以明确不同恢复措施的作用机制,为高寒湿地的保护和恢复提供理论和数据支撑。
猜你喜欢
天津农林科技(2022年2期)2022-04-19
河南畜牧兽医(2022年3期)2022-04-13
绿色科技(2021年10期)2021-06-23
黑龙江工程学院学报(2020年5期)2020-10-21
水土保持研究(2019年6期)2019-10-19
草业学报(2019年2期)2019-02-25
高原山地气象研究(2016年2期)2016-11-10
海南热带海洋学院学报(2015年5期)2015-03-14
江苏农业科学(2014年3期)2014-07-16
塔里木大学学报(2014年3期)2014-03-11
