
亚热带针阔混交林优势种黄山松和马褂木的空间格局
——以浙江景宁草鱼塘为例
2021-12-07 13:35:30余丽慧姚良锦徐端妙夏丽敏陈孟广焦洁洁
江苏林业科技 2021年5期
余丽慧,姚良锦,徐端妙,夏丽敏,陈孟广,焦洁洁
(1.浙江景宁县林业总场,浙江 景宁民族自治县 323500; 2. 浙江省林业科学研究院,浙江 杭州 310023;3. 中国林业科学研究院森林生态环境与保护研究所,国家林业和草原局森林生态环境重点实验室,北京 100091)
基于不同种群的空间分布特征和格局动态研究,探究种群间的共存机制和种群对生境条件的适应程度[1-3],有助于对种群动态和外来物种的入侵和定居提供理论指导。物种的生长状况和对资源的利用情况及群落结构的影响均能通过空间分布格局展现出来[4-6],能够更好地了解物种独特的生境喜好和种内与种间相互关系。森林群落中的优势种作为群落的建群种,决定了群落结构和环境的形成,是控制森林群落外貌特征和群落类型的关键因素[7-9],对生境有着较大的影响和较大的资源利用率。对优势物种进行空间格局动态分析,有利于对群落结构的形成和多样性维持机制有更深入的认识。
森林群落中的多数物种在资源的利用、寄生或被捕食以及物种的繁殖特性等均能从空间分布的聚集程度上体现出来,而空间分布格局又受到尺度的影响,随着距离的变化,物种可能由聚集分布转变为随机分布或均匀分布[4]。物种间的相关性也在不同尺度上表现出差异性,受尺度影响由正相关性转变成负相关性或无明显相关性[10]。点格局分析法[11-13]通过以物种个体分布的空间坐标为基础,对任意距离尺度下的种群分布格局和种间相互关系进行合理分析。目前,多尺度分析比较集中在对热带森林群落的研究,一般采用空间点格局方法来探究群落中不同物种的空间分布格局和空间关联性[3,9,14-16]。虽然亚热带地区森林群落生物多样性更为丰富,但是受地形等限制,大面积的物种空间数据取样较难获取,因此,关于通过多尺度格局动态分析亚热带天然森林植被的研究相对较少。
植物的生物学特性、生境条件以及森林系统植物间的化感效应等显著影响了群落结构和物种分布格局[17-18]。植物间的化感作用在生态修复和重建过程中越来越受到重视[19-20],显著影响着森林群落的天然更新。植物化感作用是指供体植物(如松科植物)等通过挥发、分泌、淋溶和降解等方式释放次生化合物并进入环境(特别是土壤和空气)中,从而对邻近受体植物(如田间杂草等)生长发育形成抑制或相互促进关系的化学生态学现象[21-22]。Lodhi[23]通过研究不同优势树种间的化感作用,发现不同树种释放的植物化学物质会累积在空气或土壤中,改变了附近的微生境条件,抑制或促进其他植物的生长发育。松科树种能分泌刺激性化学物质,被认为对其他物种具有很强的化感作用,造成群落结构简单、物种组成单一、林地生产力下降和地力退化[24]。化感作用能明显形成密度制约效应[25],形成开阔的空间和环境,给其他树种的定植和生长给予空间和资源。
浙江省亚热带针阔混交林样地是介于阔叶林(常绿和落叶)和针叶林之间的过渡性类型,在浙江省主要分布于高海拔山地,群落内物种组成丰富,群落结构复杂。该区域原生植被受早期农耕或木材采伐等人为干扰造成了很大的破坏,后经建立自然保护区实施封育天保工程,森林植被得以修复,形成了目前的针阔混交林群落。黄山松(Pinustaiwanensis)种群与马褂木(Liriodendronchinense)种群是针阔混交林样地中的优势种和建群种,黄山松和马褂木是落叶乔木,作为森林植被类型中的演替先锋树种,在森林恢复中发挥着积极的作用。对次生林优势种群的研究,对了解森林演替的一般规律具一定的价值[26-28]。本文以浙江景宁草鱼塘为例,运用Ripley’s L-函数分析我国亚热带针阔混交林固定样地内优势种群黄山松与马褂木不同径级结构的空间分布格局特征,采用双变量成对相关函数(pair correlation function,PCF)分析不同物种间的空间关联性,以期研究我国亚热带针阔混交林优势种的空间分布特征,以及这些空间分布特征在亚热带针阔混交林群落构建、物种共存或生物多样性维持机制中的作用。
1 材料与方法
1.1 研究区域概况
研究区域位于浙江省景宁畲族自治县东南部草鱼塘林场内 (东经119°38′28″—119°41′12″、北纬27°54′4″—27°55′48″),属亚热带海洋性季风气候区,沿海山地湿润型气候,温暖湿润,雨量充沛,四季分明,自然气热条件十分优越。年平均气温12.8 ℃, 1月平均气温2.8 ℃, 7月平均气温22.6 ℃。无霜期196 d左右。年日照时数1 617.6 h,大于0 ℃的年积温6 401.4 ℃,大于10 ℃年积温5 493.5 ℃。年降水量1 918 mm。森林覆盖率96.85%,主要树种为马褂木、云锦杜鹃 (Rhododendronfortune)、微毛柃 (Euryahebeclados)、黄山松、红豆树(Ormosiahosiei)等。
1.2 数据收集
2018年5—7月,在浙江景宁草鱼塘建立1个1 hm2(100 m ×100 m)的亚热带针阔混交林动态监测样地,进行群落调查。采用径级结构来替代年龄结构进行种群的空间格局动态分析。结合浙江省针阔混交林中实际的径级情况,按胸径(DBH)的大小来划分年龄段,分别为径级I (幼树, 1 cm ≤DBH≤5 cm)、径级II (中树, 5 cm ≤DBH≤10 cm)、径级III (大树,DBH≥10 cm) 3个生长阶段[9,29]。
1.3 点格局分析与零模型
运用Ripley’s K函数描述样地中树木的空间分布特征[30]。Ripley’s K函数为[11]:
式中,M为样地内研究物种的总个体数;d为以研究目标个体为圆点的取样圆半径;I为指示函数,dij为树木个体i和j之间的距离。当dij≤d时,I=1.0;当dij>d时,I= 0[30-31];Wij为边界效应的校正系数,是以树木i为圆心,以dij为半径的圆在样地中的周长与圆的周长之比的倒数;A为研究样地的面积。


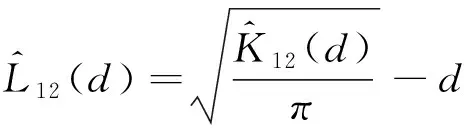
2 结果与分析
2.1 黄山松与马褂木的径级结构
整个样地中黄山松2 112 株,马褂木567株。从图1可以看出,黄山松与马褂木的径级结构均呈反“J”型。黄山松种群中存在大量的胸径在1—2.5 cm的小树,49.5%的黄山松个体胸径在1—5 cm之间,胸径在30 cm以上的黄山松有18株;马褂木种群幼小植株更多,70.9%的马褂木个体胸径在1—5 cm之间,胸径在30 cm以上的马褂木仅有11株。黄山松的各年龄段个体数量上趋于相对接近;马褂木的各年龄段数量从小树到大树逐渐减少。

A黄山松,B马褂木图1 黄山松与马褂木种群的径级分布
2.2 黄山松种群与马褂木种群空间分布格局
在0—30 m的空间尺度上,黄山松种群表现出聚集分布,且随着空间距离的增大,聚集程度降低至随机分布(如图2-A)。具体来看,小尺度上的黄山松小树和中树表现出聚集状态,大尺度上表现出随机分布(如图3-A,-B),而大树在0—100 m的尺度上呈现出随机分布(如图3-C);马褂木种群在0—100 m的尺度上呈随机分布(如图2-B)。小尺度上马褂木小树、中树和大树均呈现聚集分布,且随着距离的增大,聚集程度降低至随机分布(如图3-D,-E, -F)。

注:横坐标表示空间距离,实线值表示实际计算值,虚包迹线为由异质Poisson模型拟合模拟产生的95%置信区间的上限和下限。(A黄山松,B马褂木)图2 黄山松与马褂木种群所有个体的分布格局

注:横坐标表示空间距离,实线值表示实际计算值, 虚包迹线为由异质Poisson模型拟合模拟产生的95%置信区间的上限和下限。(A,B,C分别为黄山松小树、中树、大树;D,E,F分别为马褂木小树、中树、大树) 图3 黄山松种群与马褂木种群不同径级结构的空间分布
2.3 黄山松种群与马褂木种群不同径级间的空间关联
当d≤19 m时,黄山松种群中树对小树表现为正相关,d在20—30 m时呈现出显著负相关,当d>30 m时表现出无显著相关性(如图4-A)。当d≤ 25 m时,黄山松种群大树对小树和中树表现为负相关,当d>38 m时表现出无显著相关性(如图4-B,-C)。当d≤14 m时,马褂木种群大树和中树对小树表现为正相关,在19—35 m时呈现出负相关,当d>40 m时表现出无显著相关性(如图4-D,-E)。当d≤35 m时,大树对中树呈显著负相关,大尺度上(d>35 m)无显著相关性(如图4-F)。

注:横坐标表示空间距离,实线值表示实测观察值, 虚包迹线为由异质Poisson模型拟合模拟产生的95%置信区间的上限和下限。(A黄山松种群的中树对小树;B黄山松种群的大树对小树;C黄山松种群的大树对中树;D马褂木种群的中树对小树;E马褂木种群大树对小树;F马褂木种群的大树对中树)图4 黄山松和马褂木种群不同径级间空间相关性
2.4 黄山松与马褂木种群不同径级间的空间关联性
黄山松种群对马褂木种群的各个径级段在小尺度上(d≤20 m)都表现为显著负相关性,随着尺度的增大,相关性减弱直至无显著相关性(如图5)。马褂木种群小树对黄山松种群小树和马褂木种群大树对黄山松种群大树在任何尺度上均表现出无显著相关性(如图6-A,-F)。马褂木种群中树对黄山松种群小树和中树在小尺度上(d≤35 m)呈显著负相关,在大尺度上(d>40 m)无显著相关性(如图6-B,-C)。而马褂木种群大树对黄山松种群小树和中树在小尺度上(d≤10 m)表现出显著正相关,随着尺度的增大(d>15 m),表现出无显著相关性(如图6-D,-E)。
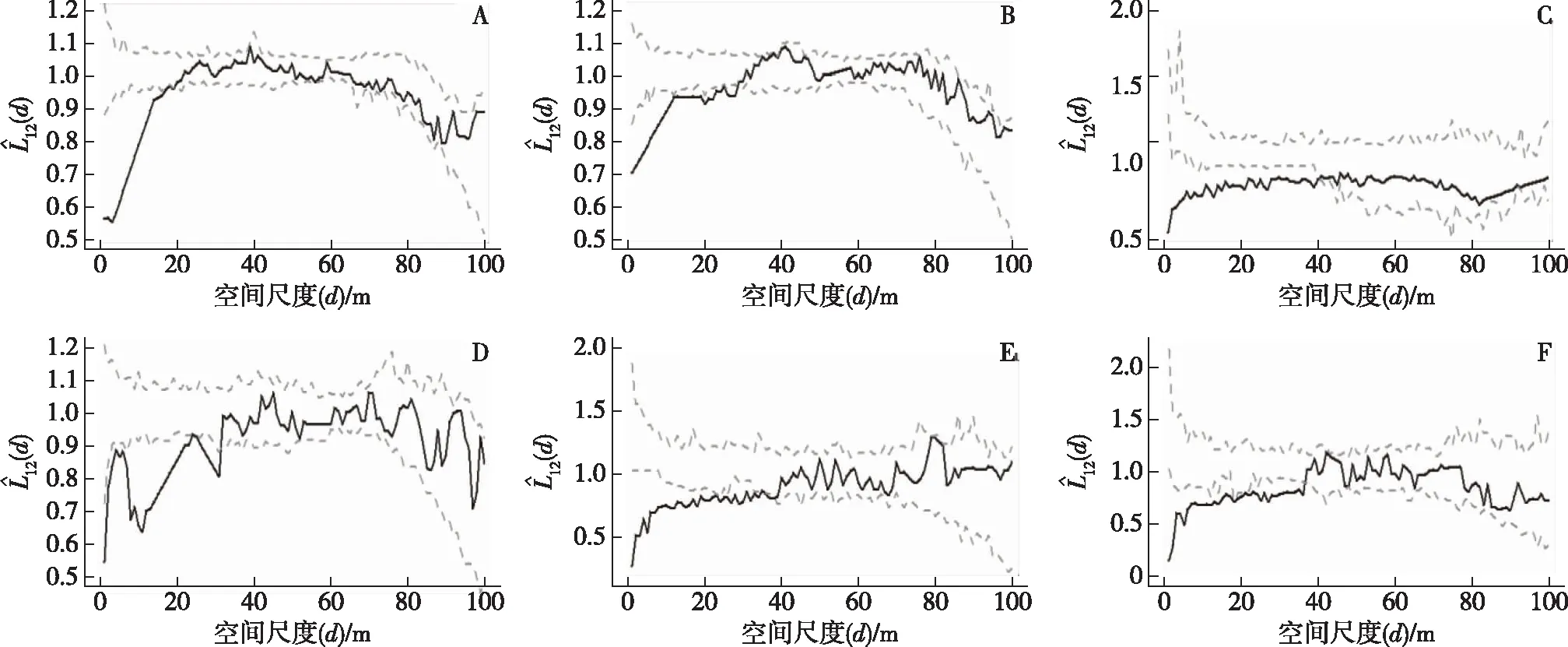
注:横坐标表示空间距离,实线值表示实际计算值, 虚包迹线为由异质Poisson模型拟合模拟产生的95%置信区间的上限和下限。(A黄山松小树对马褂木小树;B黄山松中树对马褂木小树;C黄山松中树对马褂木中树;D黄山松大树对马褂木小树;E黄山松大树对马褂木中树;F黄山松大树对马褂木大树)图5 黄山松种群对马褂木种群不同径级间空间关联性

注:横坐标表示空间距离,实线值表示实际计算值, 虚包迹线为由异质Poisson模型拟合模拟产生的95%置信区间的上限和下限。(A马褂木小树对黄山松小树;B马褂木中树对黄山松小树;C马褂木中树对黄山松中树;D马褂木大树对黄山松小树;E马褂木大树对黄山松中树;F马褂木大树对黄山松大树)图6 马褂木种群对黄山松种群不同径级间空间关联性
3 讨论
浙江针阔混交林样地内优势树种黄山松和马褂木在径级分布上都是呈反”J”型,黄山松和马褂木种群在各径级上的个体数量呈增长型。这2个树种都有着良好的天然更新,符合演替过程中稳定群落物种逐渐在群落中占据生长优势,在演替后期演替先锋种生长优势逐渐展现出弱化的趋势。黄山松种群和马褂木种群都是以种子更新为主要更新方式,萌芽更新较少。浙江景宁亚热带针阔混交林样地属于典型演替早期次生样地,样地内大径级木(胸径≥30 cm)的较少,黄山松与马褂木每年会产生大量的种子,故样地林分恢复良好,以小径级木(胸径≤10 cm)为主要林分构成。物种的空间分布上可以看出其特殊的生境喜好。黄山松作为该地区典型的演替先锋树种,具有快速扩散和繁殖的能力;在山脊位置较好,光照充足的环境条件更有利于黄山松的更新和生长发育。马褂木一般生长在山坡等水分含量较高,地质条件较好的生境条件下;而在样地中海拔较高区域内,由于受人为干扰严重,林地恢复年限较短,恢复程度较慢,马褂木作为阔叶树种,可能受生境的影响,尚未分布于该区域内。
树种的空间分布格局主要是由生物过程和非生物过程决定。生物过程包括了更新、繁殖、扩散和竞争等;非生物过程有生境异质性、干扰或促使物种个体非随机分布的随机事件等。在对这2个优势树种的空间格局动态分析的研究中,发现它们在总体上以及不同年龄阶段上主要呈聚集分布,这可能与种子扩散的限制或生物学特性有关。黄山松种群大树母株产生大量种子,由于黄山松种子受动物的喜食,传播范围更广。并且黄山松种群小树具有较强的喜光性,使得其幼苗和中树大部分都在林地内光照条件较好和林隙较大的区域内聚集分布。本研究发现黄山松和马褂木种群的聚集程度随着径级的增加均表现出下降的趋势。黄山松种群大树和马褂木种群中树在较大取样尺度上均出现随机分布。物种对外界资源 (光照、水分、营养、空间等) 的需求随着径级的增大而增多,加剧种内与种间的相互作用程度[9,37],引起群落中植物自疏或它疏现象,降低物种的聚集程度。因此,在不同径级阶段的树种对生境条件表现出不同的生境需求。小树、中树更多聚集于生境条件较好的地区,更有利于其生长发育繁殖;中树,大树因为对生境空间需求更大,竞争更大,聚集程度相对较小,在样地中较大距离尺度上多表现为随机分布[3]。这2种物种对生境的适应性,都表现在更有利于物种的生长和繁育。黄山松和马褂木种群小树到大树的聚集强度逐渐减弱,表现出不同物种间在空间尺度上不同的聚集表现。径级的增大加强了密度制约效应或病原体的影响将导致种群聚集强度的减小和使相邻个体间的距离增大。这一过程在幼苗阶段到大树阶段一直存在[38-39]。这2个物种的空间格局动态与李立所研究的古田山优势树种甜槠和木荷以及汤孟平在天目山的优势树种所做的研究在一定的空间尺度上的研究结果基本一致。
环境筛选(environmental filtering)和生物竞争(biotic competition)是群落构建中2个最主要的机制[1]。环境筛选作用首先决定了进入群落的物种性状分布范围,生物竞争作用则通过提高物种性状差异来减少竞争强度[40]。黄山松种群大树和中树对小树均在较大距离尺度上出现负相关性,即不对称竞争加剧,出现大树或中树制约小树的生长;而马褂木的大树和中树对小树在小尺度上均表现为正相关性,即对称性竞争,促进马褂木小树的生长。这可能与不同物种的生物学特性有关。黄山松是演替先锋种,生长发育较为迅速,需要从外界获取大量的光照和营养物质等,个体间相互竞争加剧影响了小树的生长;并且大树、中树中携带的病虫菌等有害物质较多,而小树对外界的抵抗能力较差,受到病虫害的威胁更大,抑制了小树与中树和大树之间的共存,造成小树有较高的死亡率。说明了黄山松的大树和中树是抑制小树生长的瓶颈,对后期的森林恢复和抚育有很重要的影响。
植物的竞争是一种邻域的现象,受尺度和自身特性影响着植物的分布格局[41]。从黄山松种群与马褂木种群间的空间相关性来看,黄山松种群对马褂木种群的个体之间有着明显的不对称竞争,对它们的生长发育有着明显的抑制作用,这可能与针叶树种的化感作用有关。植物化感作用对群落的植物组成及分布及其演替过程具有重要作用[21-22]。基于不同树种间的空间关联性结果,发现黄山松对马褂木种群有着极显著的种间竞争作用。种间竞争一直是热带和亚热带森林群落稳定共存的重要驱动机制[42-43]。黄山松一般多采用次生代谢的方式释放化学物质(酚类、萜类、生物碱等)[44],通过化感作用抑制邻体物种生长,满足自身生长需求。化感作用是导致密度制约效应主要原因之一,阔叶树种(马褂木)受针叶树种的化感作用抑制生长,形成密度制约效应[25],提供资源和空间以利于其他树种的生长。在本文所研究的针阔混交林群落中,针叶树种林下植物种类和数量极少,草本层盖度在30%以下,且主要零星簇状分布在相对坑洼的区域。这可能是因为针叶树种的化感物质产生的植物毒素不断积累,降低了群落内的生物多样性,造成了整体林分生产力的下降和林地的地力退化[23-24]。因此,在针阔混交林的天然更新与林业经营中要充分考虑化感作用的影响。
致谢感谢浙江省林业科学研究院与中国林业科学研究院的重大合作项目“基于功能群的亚热带次生阔叶林质量提升技术研究与示范”(No. 2019SY08)的资助。感谢中国林业科学研究院森林生态环境与保护研究所王青博后对文章的指导与建议。
猜你喜欢
广西林业科学(2022年4期)2022-11-07 01:50:34
林业调查规划(2022年3期)2022-06-13 09:31:04
广西林业科学(2021年6期)2022-01-21 07:39:10
黄山学院学报(2017年5期)2017-11-03 02:41:28
山西教育·幼教(2017年11期)2017-04-13 23:40:30
武夷学院学报(2015年3期)2015-07-18 11:03:45
西北植物学报(2015年9期)2015-07-05 07:03:42
红领巾·萌芽(2014年1期)2014-12-01 09:09:01
创新作文(1-2年级)(2014年5期)2014-07-19 02:33:12
高中生·青春励志(2008年8期)2008-08-22 01:13:08
