
柳树AP2/ERF基因家族全基因组鉴定和表达分析
2021-12-07 13:35:32戴晓港李淑娴
江苏林业科技 2021年5期
戴晓港,李淑娴
(南京林业大学林学院,南方杨树工程技术研究中心,江苏省杨树种质创新与品种改良重点实验室,江苏 南京 210037)
发展可再生能源是应对温室效应引起全球气候变化的重要手段[1]。林木为人类提供了最重要的可再生资源,杨柳科植物由于易于繁殖、生长速度快、产量高,是营建生物质能源林重要的树种之一[2]。全球柳属有520个种,主产北半球温带地区[3],我国柳树资源丰富,包含种257个,变种122个和变型33种,且基本处于野生和半野生状态,其遗传变异丰富,改良潜力大[4]。全球范围内,培育高产和抗逆性强的柳树无性系以适应边际土地开发和利用的需求也在不断增长。
AP2/ERF转录因子是植物中最大的转录因子家族之一。在模式植物中,AP2/ERF基因家族的生物学功能已有深入研究。研究结果表明,该基因家族在很多植物生长过程中起着至关重要的作用,这些过程包括调控植物生长和发育[5-9],对生物胁迫和非生物胁迫的应答[11-16]等。如AP2亚家族中的基因被发现参与调控植物开花时间[5]、调节花器官生长和发育[6]、胚珠发育[7]、决定小穗分生组织形成[8]及叶表皮细胞形成[9],而该基因家族的RAV基因受乙烯和油菜素内酯等激素调控[10],过表达此类基因可以提高植物对盐胁迫和干旱胁迫的抗性[11-12],ERF亚家族基因则主要参与对生物胁迫和非生物胁迫的应答,包括病原菌和疾病刺激[13-14]、水分胁迫[15]、盐胁迫[16]及高温和低温胁迫等[17-18]。
AP2/ERF基因家族重要的生物学功能,使得该家族的基因成为植物育种和基因工程重要的基因资源。簸箕柳基因组染色体水平的组装序列[19],为开展簸箕柳全基因组AP2/ERF基因家族分析提供了基础。对簸箕柳AP2/ERF基因家族开展研究,有助于全面了解柳树中该家族基因的组成和进化历史。柳树不仅耐水湿,而且对干旱生境的适应性很强[20]。本文还对柳树干旱条件下差异表达的AP2/ERF基因进行了分析,旨在获得柳树与干旱胁迫应答有关的AP2/ERF基因家族候选基因。
1 材料与方法
1.1 AP2/ERF转录因子查找
簸箕柳基因组参照Wei等[19]发表的序列。基因家族中基因查找方法如下:首先,以拟南芥AP2/ERF基因家族中具有代表性的AT1G25560.1和AT2G28550.1为参考序列[21],采用BLASTp(2.2.26+)[22]将所有柳树蛋白质序列比对到上述2个基因序列上,比对参数为E-value=1e-5,Identity≥50%;然后,将所有柳树蛋白质序列采用hmmer-3.0的默认参数[23]和含有AP2 domain的Pfam文件PF00847比对查找含有AP2 domain的簸箕柳基因;最后,将以上查找的基因去冗余后,再采用SMART(http:∥smart. embl-heidelberg.de/)在线搜索[24],进一步确认上述查找的序列中是否含有AP2 domain序列。
1.2 AP2/ERF转录因子系统进化
将拟南芥AP2/ERF基因家族[25]和簸箕柳AP2/ERF基因家族蛋白质的domain序列采用ClustalW2.1[26]进行多重比对,所有参数均为默认设置。采用MEGA-X[27]中的邻接法构建进化树,参数设置为泊松校正、pair-wise deletion、bootstrap,重复为1 000次,采用Figtree对构建的进化树进行可视化(http://tree.bio.ed.ac.uk/software/figtree/)。
1.3 AP2/ERF转录因子保守基序和基因结构
利用在线预测工具MEME(Multiple Em for Motif Elicitation, https://meme-suite.org/meme/tools/meme)对簸箕柳AP2/ERF蛋白的保守基序(Motif)进行分析,设置Motif个数为10个,Minimum width=10, Maximum width=70,其他参数均为默认值。根据基因注释的gff文件,利用TBtools[28]进行基因结构可视化。
1.4 AP2/ERF转录因子启动子顺式作用元件
根据基因组注释文件,提取基因家族成员起始密码子上游2 000 bp序列作为基因的启动子序列,用Plant CARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)在线进行预测分析,按照顺式元件中与非生物胁迫和激素相关的顺式作用元件对预测结果分类整理,最后利用TBtools[28]工具整合进化树和对启动子区域的顺式作用元件进行可视化。
1.5 AP2/ERF转录因子染色体分布及扩增方式
根据AP2/ERF基因注释gff文件中基因的位置,采用MapChart[29]将AP2/ERF基因绘制在染色体上。
1.6 AP2/ERF转录因子表达分析
从EBI网站中下载了蒿柳杂交子代(S.viminalis×[S.viminalis×S.schwerinii)]基因型520号干旱胁迫处理和对照的根系转录组数据(http://www.ebi.ac.uk/ena/data/view/PRJEB10883),使用BWA[30](0.7.15-r1140)软件将上述序列分别比对到所有AP2/ERF基因上,比对参数为小于等于2个错配,其余为默认参数,根据比对结果,统计对照和干旱胁迫处理分别比对到每个基因的序列数量。采用RPKM (reads per kilobase of transcripts per million mapped reads) 的方法对统计每个基因表达量。用DEGseq[31]软件包来计算差异表达的基因,根据log2RPKM的值≥1,即其表达量差异超过1倍的、且P-value小于0.01的基因定义为差异表达基因。
2 结果与分析
2.1 AP2/ERF转录因子数量及分类
根据隐马尔可夫模型对Pfam文件PF00847的搜索和拟南芥序列的比对,同时采用SMART对上述结果进行验证,结果表明簸箕柳AP2/ERF转录因子基因家族中含有208个基因(见表1)。根据基因含有AP2 domain的数量和序列的相似度对这208个基因进行分类,24个基因含有2个完整的AP2 domain序列被分到AP2家族,177个基因属于ERF家族,其中每个基因只含有1个AP2 domain;4个基因属于RAV家族,每个基因除了含有1个AP2 domain,还有1个B3 domain。
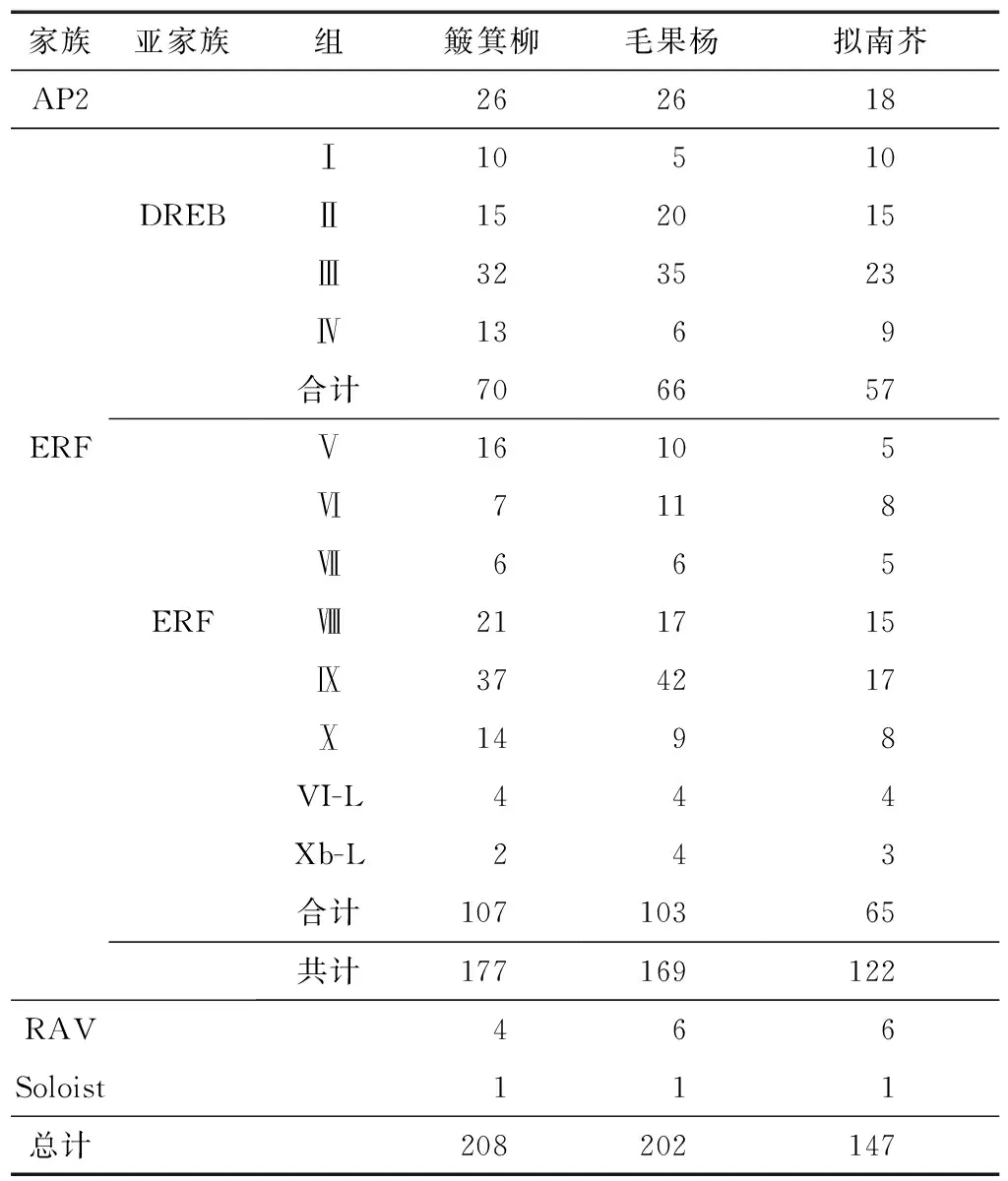
表1 簸箕柳AP2/ERF家族基因与毛果杨和拟南芥的比较
2.2 AP2/ERF转录因子进化分析
以簸箕柳和拟南芥基因蛋白质保守域序列,用MEGAX构建簸箕柳AP2/ERF基因家族的进化树。进化分析结果表明,虽然EVM0033490.1和EVM0002376.1只含有1个AP2 domain序列,但是这2个基因和AP2家族基因序列相似度较高,将这2个基因分类到AP2家族;还有1个基因EVM0011433.1虽然只含有1个AP2 domain序列,但与ERF和AP2家族分化较远而被定义为Soloist基因。按照Nakano等[25]对AP2/ERF基因家族的分类标准,ERF家族中的177个基因细分为ERF亚家族和DREB亚家族共12个group,其中DREB亚家族4个group (I—IV)分别含有10,15,32和13个基因成员,ERF亚家族8个group (V—X, VI-L, Xb-L) 分别含有16,7,6,21,37,14,4和2个基因成员(如图1)。
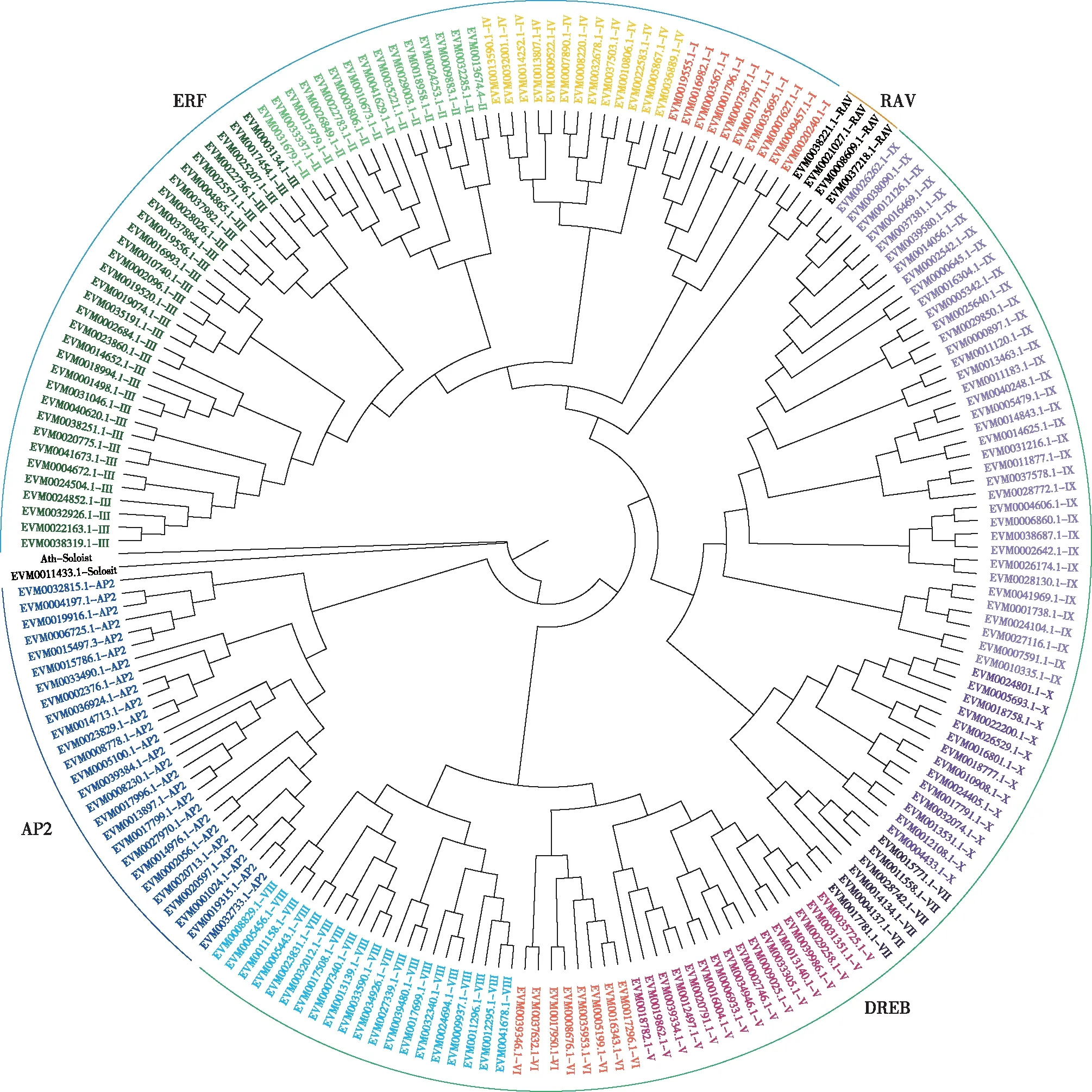
图1 簸箕柳208个AP2/ERF转录因子进化树
2.3 AP2/ERF转录因子蛋白基序分析
利用MEME分析簸箕柳AP2/ERF基因家族的Motif,结果显示只有Solisit基因含有1个Motif,其余所有AP2/ERF家族基因含有2—6个不等的Motif(如图2)。簸箕柳208个AP2/ERF基因,98.6%的基因都含有Motif 2,其次93.8%和91.8%的基因含有Motif 1和Motif 4。大多数的AP2亚家族的基因都有6个Motif,其中Motif 3,Motif 5和Motif 6是AP2亚家族特有的Motif,AP2亚家族的26个基因中,分别有24个基因含有Motif 3、25个含有Motif 5、23个含有Motif 6。Motif 8是ERF亚家族中特有的,其中group Ⅲ的32个基因中均含有Motif 8。Motif 9和Motif 10是DREB亚家族中的group IX特有的。

图2 簸箕柳AP2/ERF转录因子蛋白基序
2.4 AP2/ERF家族基因结构
对簸箕柳AP2/ERF家族基因结构分析发现,不同基因亚家族之间基因结构存在较大的差异。AP2家族中的26个基因全部含有内含子,其内含子数目在5—11个不等;而ERF家族70个基因中,只有14个基因含有内含子,除了EVM0020240.1基因含有3个内含子外,其余基因内含子数量为1—2个;DREB家族中107个基因,也只有48个基因有内含子,其内含子数目在1—2个,而含有内含子的基因主要集中在V,VII和X 3个亚家族,共30个基因含有内含子;RAV家族中的4个基因都没有内含子,这与AP2家族中的基因结构截然不同(如图3),而Soloist家族中1个基因含有5个内含子。

图3 簸箕柳AP2/ERF家族基因结构
2.5 AP2/ERF转录因子启动子区域顺式作用元件
为了解簸箕柳AP2/ERF家族基因的潜在功能,利用PlantCARE软件对基因上游2 000 bp的序列开展启动子顺式作用元件分析。结果显示(如图4),顺式作用元件中含有参与激素、光、低温、防御胁迫、昼夜节律控制和干旱诱导等响应元件。在激素类响应元件中,响应水杨酸和茉莉酸甲酯应答的基因最多,分别有148个和138个基因,而应答生长素和赤霉素的基因成员较少,分别有21个和34个。簸箕柳AP2/ERF家族中也有大量参与非生物胁迫等相关的作用元件,如干旱诱导、低温响应、防御胁迫等。其中分别有89个基因成员含有干旱诱导和防御胁迫作用元件,78个基因含有低温响应作用元件。推测这些基因在对非生物胁迫的应答中起着重要的作用。
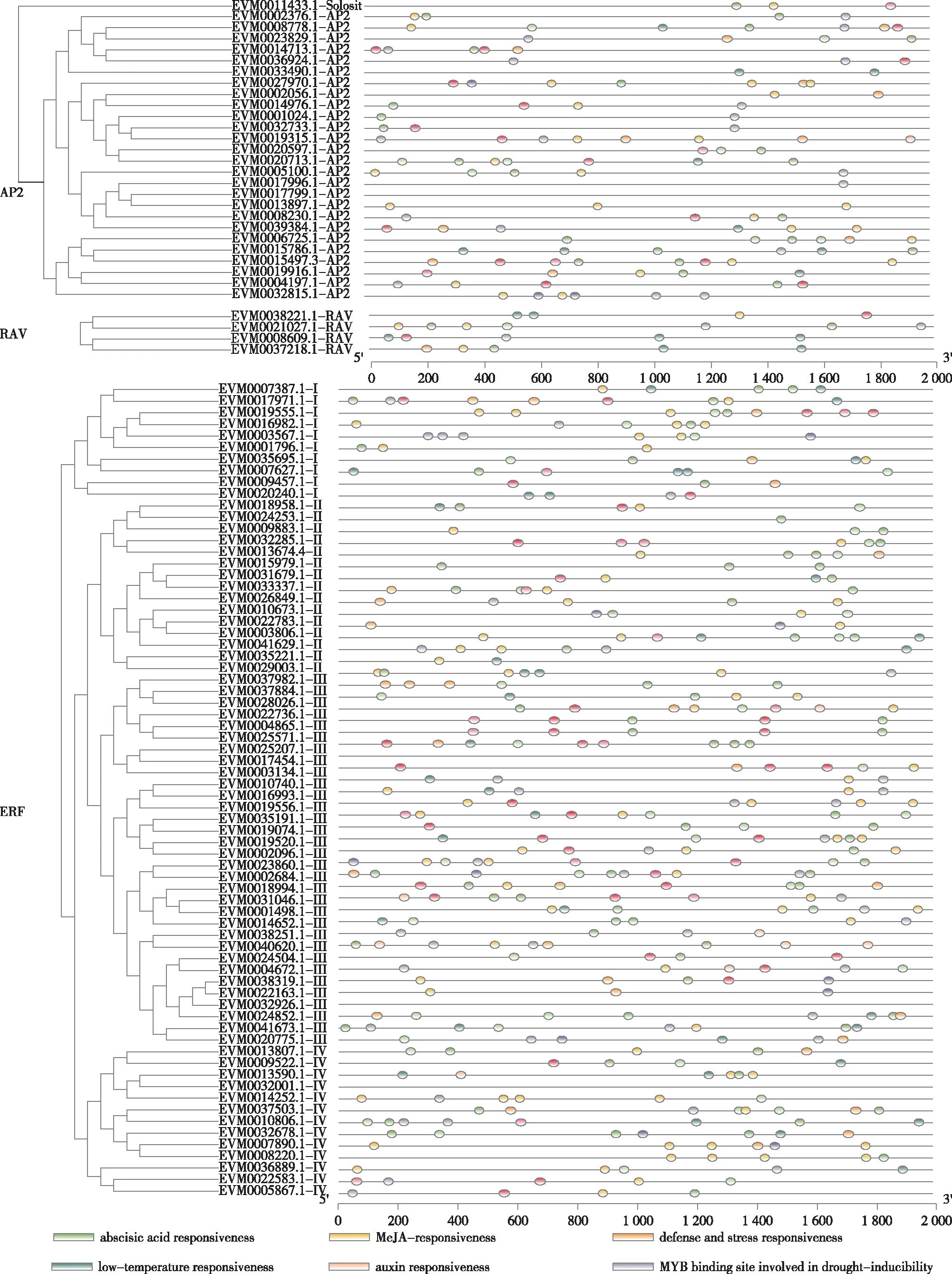
图4 簸箕柳AP2/ERF转录因子启动子区域顺式作用元件
2.6 AP2/ERF基因在染色体上的分布
通过物理作图绘制出簸箕柳AP2/ERF基因在染色体上的分布(如图5)。从图5中可以看出,簸箕柳AP2/ERF基因在19条染色体上的分布是不均匀的,Ⅲ号染色体和XVI号染色体含有AP2/ERF基因最多,分别含有23和22个基因,Ⅱ号染色体含有18个AP2/ERF基因,V号和Ⅷ号染色体分别含有15个AP2/ERF基因,VI号和X号染色体分别含有15个AP2/ERF基因,然而IX号染色体却只含有2个AP2/ERF基因。这个结果和对杨树AP2/ERF基因家族的分析相似,多数AP2/ERF基因成簇的聚集在一起。基因的复制方式分析发现,这个基因家族中有19个串联复制事件共产生42个AP2/ERF基因,每个串联复制基因簇基因数量为2—3个。在这19条染色体中,Ⅱ,Ⅲ和XVI号染色体均发生3次串联复制包含20个基因,VI和XIV号染色体发生2次串联复制包含8个基因,有7条染色体上均检测到1个串联复制。串联重复导致基因家族成员扩张是引起基因家族成员在染色体上分布不均匀的重要原因之一。
2.7 柳树AP2/ERF抗旱相关基因分析
Pucholt等[32]完成了蒿柳杂交子代干旱胁迫及对照根系的转录组测序,得到了一批抗旱有关的基因,主要包括MYBs转录因子家族、bZIPs转录因子家族和叶绿素a/b结合蛋白等,有1个AP2基因和ERF基因在对照和干旱胁迫中存在差异表达。将蒿柳杂交子代干旱胁迫及对照处理根系的转录组序列分别比对到簸箕柳AP2/ERF家族基因序列上,查找差异表达的基因,结果如表2所示。从表2中可以看出,蒿柳杂交子代在干旱胁迫下有5个ERF基因(见表2)和对照相比存在显著的上调表达,Log2RPKM都大于1。在这5个上调表达的基因中,EVM0039334.1和EVM0014843.1属于DREB亚家族,且这2个基因的启动子区域都分别含有1个干旱诱导的响应元件,其余3个基因属于ERF亚家族,其中EVM0028026.1在对照处理中不表达,只有在干旱胁迫状态下才高表达。
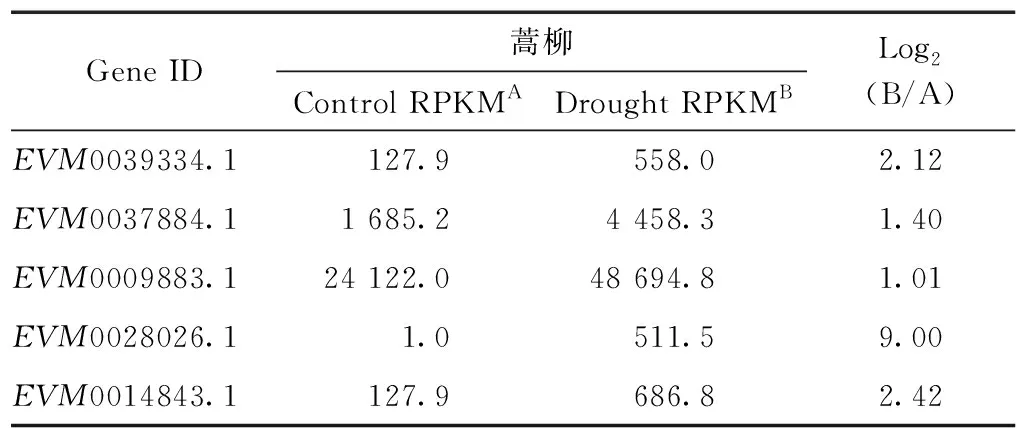
表2 蒿柳杂交子代干旱处理后根系差异表达的AP2/ERF基因
3 讨论
本研究从簸箕柳的全基因组中查找出208个AP2/ERF转录因子,基因家族成员总数量和毛果杨(202个)[33]中的数量基本相等,高于钻天柳(173个)[34]和拟南芥(148个)[25]中的数量。根据Nakano等[25]的分类标准,簸箕柳的208个AP2/ERF转录因子,其中26个基因属于AP2家族,107个基因属于DREB亚家族,70个基因属于ERF亚家族,4个基因被分类到RAV家族中,还有1个基因被分类到Soloist中。在簸箕柳和毛果杨基因组中,AP2/ERF转录因子每个亚家族基因的数量几乎是相等的,都含有26个AP2基因和1个Soloist基因,ERF亚家族分别含有70个和66个基因,DREB亚家族中分别含有107个和103个基因,簸箕柳RAV家族只含有4个基因,而毛果杨中含有6个基因。每个亚家族中基因的数量分布在杨树和柳树中基本相似,上述结果也说明了AP2/ERF转录因子家族在杨柳科植物的进化过程中是高度保守的。
AP2/ERF转录因子在植物基因组中的分布是不均匀的,如毛果杨中40%的基因主要分布在其中的4条染色体上(2n=38),葡萄[35]中36%的基因主要分布在其中的3条染色体上(2n=38),AP2/ERF基因在簸箕柳染色体上的分布与杨树相似,58.2%的基因主要分布在其中的7条染色体上(2n=38)。这种基因在染色体上的不均匀分布可能与基因复制扩张的方式有关[33-34]。在簸箕柳208个AP2/ERF转录因子中,有42个基因是通过串联复制产生的,如第Ⅱ,Ⅲ和XVI是含有AP2/ERF转录因子最多的染色体,在这3条染色体上共有9个串联复制,包含20个AP2/ERF基因。簸箕柳第I号染色体的物理长度约是其他18条染色体长度的2倍,但簸箕柳第I染色体只含有12个AP2/ERF转录因子,其数量约是第Ⅲ和第XVI等染色体的1/2。
AP2/ERF转录因子保守的广泛分布于植物中,参与控制植物的多种代谢[36],主要包括控制植物生长和发育、对生物和非生物胁迫的应答反应等。由于该基因家族的可塑性和这个基因家族每个成员的特异性,通过转基因和过表达AP2/ERF某些转录因子可以提高植物对生物胁迫和非生物胁迫的抗逆性,如DREB亚家族转录因子主要和顺式作用脱水应答元件相互作用提高植物对胁迫的响应,在烟草中过表达AhDREB1基因可以提高抗盐和抗干旱能力[37];在拟南芥中过表达DREB19可以显著提高植株对干旱和高盐胁迫的能力,且对植株的表型不会带来有害的影响[38]。Pucholt等[32]对蒿柳杂交子代在干旱胁迫下及对照处理转录组的分析,发现了1个AP2和1个ERF基因在干旱胁迫处理后显著上调表达,而全基因组测序的完成为转录组差异表达分析提供了重要的参考,将上述对照和干旱处理获得的柳树转录组数据,比对到簸箕柳基因组上,发现5个ERF家族基因在干旱胁迫条件下显著上调表达,其中EVM0039334.1和EVM0014843.1属于DREB亚家族。之前的研究也表明,过表达DREB基因可以提高植株抗旱性[37-38],且这2个基因的启动子区域都分别含有1个干旱诱导的响应元件,推测这2个基因在柳树的抗旱中起到重要的作用。簸箕柳AP2/ERF转录因子的鉴定和分析,将为通过转基因培育对生物和非生物胁迫抗逆性高的柳树提供基因资源。
猜你喜欢
内蒙古师范大学学报(自然科学汉文版)(2021年3期)2021-06-01 08:10:30
少儿科技(2021年2期)2021-05-17 09:59:14
思维与智慧(2020年1期)2020-01-08 06:53:48
生物工程学报(2019年6期)2019-07-10 08:38:38
生物学通报(2019年1期)2019-02-15 16:33:43
生物学通报(2018年12期)2018-10-10 06:52:36
故事作文·高年级(2018年8期)2018-08-14 19:49:42
小天使·一年级语数英综合(2018年4期)2018-06-22 10:33:26
少儿科学周刊·儿童版(2017年5期)2017-06-29 22:14:38
学苑创造·A版(2016年3期)2016-04-16 17:59:03
