
海洋动物体内氧化三甲胺和甘氨酸甜菜碱的浓度特征及影响因素
2021-12-03 15:52陆长坤宋若晗曲克明崔正国赵婉玉胡清静毕相东
安徽农业科学 2021年22期
陆长坤 宋若晗 曲克明 崔正国 赵婉玉 胡清静 毕相东

摘要 氧化三甲胺(TMAO)和甜菜碱(GBT)广泛存在于海洋生物体内,它们被降解后产生的有机胺可通过海气交换进入大气中,进而可以促进新粒子生成及增长,具有潜在重要的气候效应。多数研究认为浮游植物体内含有大量的TMAO和GBT,它们是海洋大气中有机胺的主要贡献者。也有研究发现海洋动物体内也含有TMAO和GBT,但它们对大气中有机胺的贡献报道较少。概述了不同类型海洋动物体内TMAO和GBT合成方式及它们降解为有机胺的途径,归纳了不同海洋动物体内TMAO和GBT的浓度分布特征,探讨了影响动物体内TMAO和GBT浓度的因素,剖析了该领域待解决的科学问题,并对今后的研究工作进行了展望,以期为认识海洋环境中有机胺来源及其气候效应提供科学参考。
关键词 海洋动物;氧化三甲胺;甘氨酸甜菜碱;浓度;影响因素
中图分类号 S917.4 文献标识码 A 文章编号 0517-6611(2021)22-0018-11
doi:10.3969/j.issn.0517-6611.2021.22.005
开放科学(资源服务)标识码(OSID):
Concentration Characteristics and Influencing Factors of Trimethylamine Oxide and Glycine Betaine in Marine Animals
LU Chang-kun SONG Ruo-han QU Ke-ming 3 et al
(1. College of Fishery, Tianjin Agricultural University, Tianjin 300392;2.Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Key Laboratory of Sustainable Development of Marine Fisheries, Ministry of Agriculture and Rural Affairs, Qingdao,Shandong 266071;3. Laboratory for Marine Fisheries Science and Food Production Processes, Pilot National Laboratory for Marine Science and Technology (Qingdao), Qingdao,Shandong 266071)
Abstract Trimethylamine oxide (TMAO) and Glycine betaine (GBT) are widely present in marine organisms. The organic amines produced after they are degraded can enter the atmosphere through air-sea exchange, which can promote the generation and growth of new particles, which has a potentially important climate effect. Most studies believe that phytoplankton contains a large amount of TMAO and GBT, which are the main contributors to organic amines in the ocean atmosphere. Some studies have found that marine animals also contain TMAO and GBT, but their contribution to organic amines in the atmosphere is rarely reported. This article summarized the synthesis methods of TMAO and GBT in different types of marine animals and their degradation to organic amines, concluded the distribution characteristics of TMAO and GBT in different marine animals, and discussed the factors that affect the concentrations of TMAO and GBT in animals, analyzed scientific problems to be solved in this field, and forecasted future research work, in order to provide a scientific reference for understanding the sources of organic amines in the marine environment and their climate effects.
Key words Marine animals;Trimethylamine oxide;Glycine betaine;Concentration;Influencing factors
近年研究表明,大氣中的甲胺(MMA)、二甲胺(DMA)和三甲胺(TMA)可以促进新粒子的生成及颗粒物增长,进而增加云凝结核数浓度,通过改变辐射强迫对气候变化产生重要影响[1-4]。因此,有机胺是当今国际的研究热点之一。海洋是有机胺的重要来源,海洋生物体内的氧化三甲胺(TMAO)、甘氨酸甜菜碱(GBT)、胆碱(CHO)等有机胺前体物被细菌降解后会产生有机胺[5-10],然后通过海气交换进入大气中,其对全球大气中有机胺的贡献约占28%[11]。多数研究发现,海洋水体或大气中有机胺的浓度随浮游植物生物量的增加而增加,且在浮游植物体内检测出大量的TMAO和GBT[12-16],因此,推测浮游植物是海洋环境中有机胺的主要贡献者。但是,除了浮游植物外,海洋动物体内也含有大量的TMAO和GBT等有机胺前体物[17-19]。1938年Beatty[20]在腐烂的鱼体中第一次发现了TMAO的代谢产物TMA。Namies′nik等[21]在Gdańsk城市的鱼摊附近测得大气中MMA、DMA和TMA的浓度分别高达112.4、140.1和24.3 mg/m3。Hu等[15]研究发现我国近海大气颗粒物中DMA+、TMA+的浓度比世界其他海域高1~3个数量级,推测其可能与我国近海大规模的海水养殖有关。这说明海洋鱼类等动物体内的TMAO可能对大气中有机胺具有重要的贡献。尽管现阶段对于不同种类海洋浮游植物体内有机胺前体物的浓度特征及影响因素的研究开展了大量的工作[7,9,22-25],但关于海洋动物体内TMAO和GBT的浓度特征及影响因素的研究较少。此外,TMAO和GBT还具有为海洋动物提供浮力[26]、调节渗透压[27-29]、维持其蛋白质结构和功能稳定等重要作用[30-37]。笔者对不同种类海洋动物体内TMAO和GBT的代谢途径、浓度特征和影响因素等进行了详细的综述,从而为进一步认识海洋动物对大气中有机胺的贡献、评估其潜在气候效应提供理论依据。
1 海洋动物体内TMAO和GBT的合成与降解
1.1 海洋动物体内TMAO和GBT的合成
海洋动物体内的TMAO和GBT主要有2个来源(图1):①动物自身合成,一些浮游植物体内的CHO、磷脂酰胆碱和肉碱等在单加氧酶的催化下氧化为甜菜碱醛,然后在甜菜碱醛脱氢酶的作用下氧化为GBT,GBT经甜菜碱脱甲基作用后产生TMA,最后由三甲胺氧化酶将TMA氧化为TMAO[17,38-42];②从摄取的食物中直接积累[43],部分海洋动物不能合成TMAO或GBT[44],必须通过摄食藻类或小型动物等获得[ 45-49]。
1.2 海洋动物体内TMAO和GBT的降解
海洋动物体内TMAO的降解主要有2种途径(图1):①外源途径,主要通过海洋细菌产生的TMAO还原酶等降解为TMA[1 18,2 50-52];②海洋动物的内源性酶解,即在动物体内TMAO去甲基化酶的作用下直接降解为DMA和甲醛[53-55]。GBT是通过甜菜碱脱甲基作用降解为TMA[56],之后TMA经过三甲胺脱氢酶的作用生成DMA,或是先由三甲胺氧化酶催化生成TMAO,再经氧化三甲胺脱甲基酶生成DMA;最后DMA在二甲胺脱氢酶的催化作用下生成MMA,MMA再进一步被降解为氨气(图1)。
2 海洋动物体内TMAO和GBT的含量特征
2.1 海洋动物体内TMAO和GBT的含量
TMAO广泛存在于海洋动物中[57-58],其含量高达鱼类、甲壳类等动物组织干重的7%[4 59]。通过对230种海洋动物统计分析发现,海洋动物体内TMAO含量是0.01~25.01 g/kg,但不同种类动物体内TMAO的含量存在差异(表1),整体呈现软骨鱼类>头足类>甲壳类>硬骨鱼类>贝类的趋势(图2)。软骨鱼类体内TMAO平均含量为9.58 g/kg,70%软骨鱼类TMAO含量在5.00 g/kg以上,其中,鲨鱼、鳐鱼含量分别为10.09、12.41 g/kg,深海鳐更是高达21.71 g/kg[5]。软骨鱼TMAO含量约为甲壳类动物的2倍,是贝类含量的10倍之多。头足类动物TMAO含量次之,平均在8.02 g/kg,是硬骨鱼类的2倍多。甲壳类动物TMAO平均含量为5.36 g/kg,其中50%甲壳类动物TMAO含量分布在1.00~5.00 g/kg,含量最高的是真虾(Caridean shrimp),达到22.46 g/kg[60],最低的刀额新对虾(Metapenaeus ensis)仅含0.05 g/kg[61]。70%的硬骨鱼含量低于3.50 g/kg,但有个别极地硬骨鱼肌肉中TMAO含量高达21.78 g/kg[62],其中,含量最低的是长尾蛇鱼(Macrurous berglax)和鲶鱼(Silurus asotus),均为0.02 g/kg[63-64]。在所有种类海洋动物中,贝类中TMAO平均含量最低,仅为0.98 g/kg,这可能是因为其常栖息于河口附近或有少量淡水注入的浅海内湾中,环境盐度较低,从而导致TMAO含量较低。因此,各种动物生活习性以及所处环境的不同会导致体内TMAO含量存在差异。
GBT也是甲壳类、贝类等海洋动物体内主要的渗透调节物质之一,但目前国际上对于海洋动物体内GBT的报道较少。通过对42种不同种类海洋动物统计分析发现,GBT在海洋动物体内的含量为0.01~16.82 g/kg(表2),其在不同种类动物体内的含量与TMAO相反,整体呈现贝类>硬骨鱼类≈甲壳类≈头足类>软骨鱼类(图2)。含量最高的是贝类,平均在4.95 g/kg,约为软骨鱼类(2.60 g/kg)的2倍,其中贻贝含量可高达10.00 g/kg[66]。贝类体内GBT较高,可能与它们摄食含GBT较高的浮游植物有关。头足类、硬骨鱼类和甲壳类含量相差不明显,分别为4.10、4.13和 4.26 g/kg。
2.2 海洋动物组织器官中TMAO和GBT的分布特征
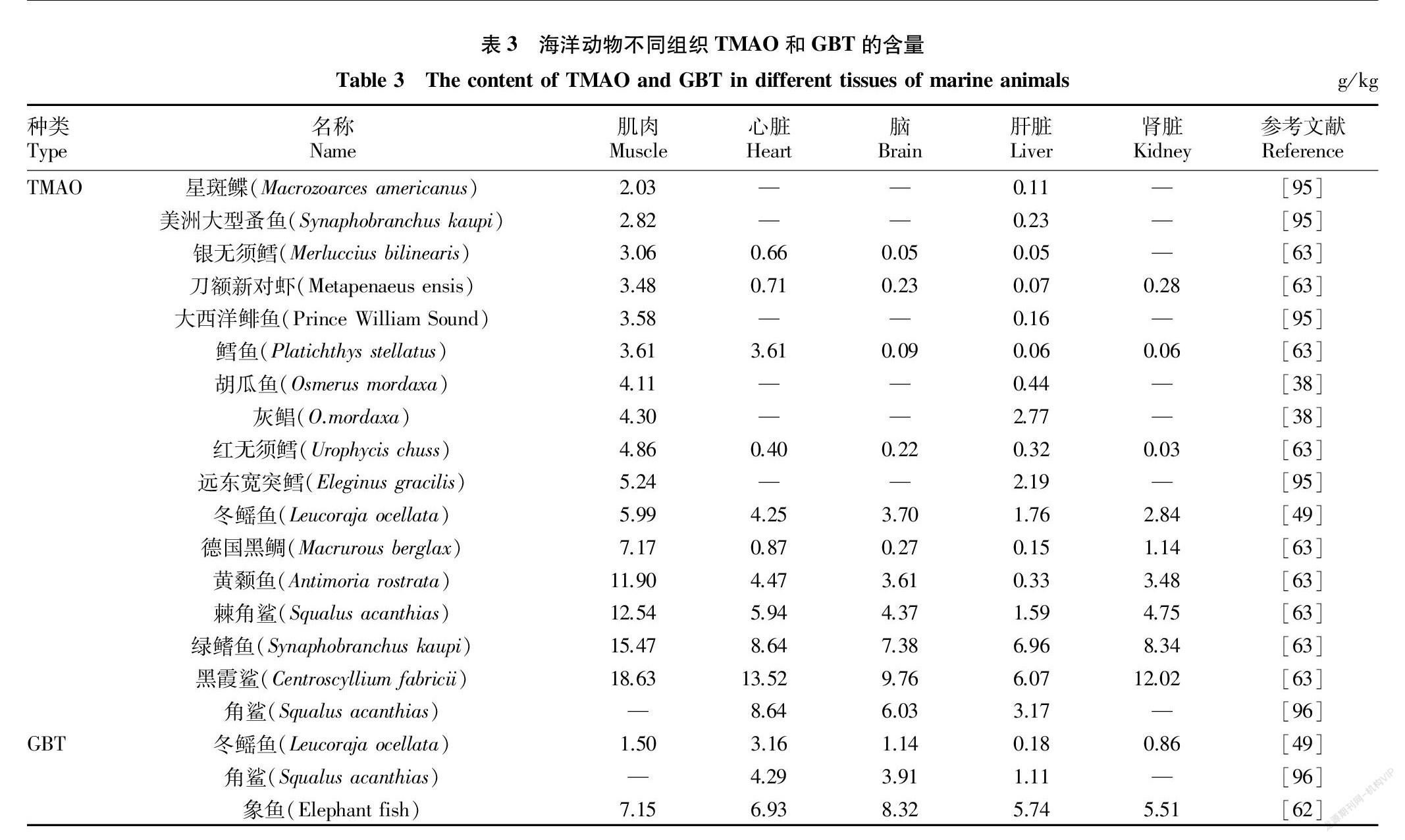
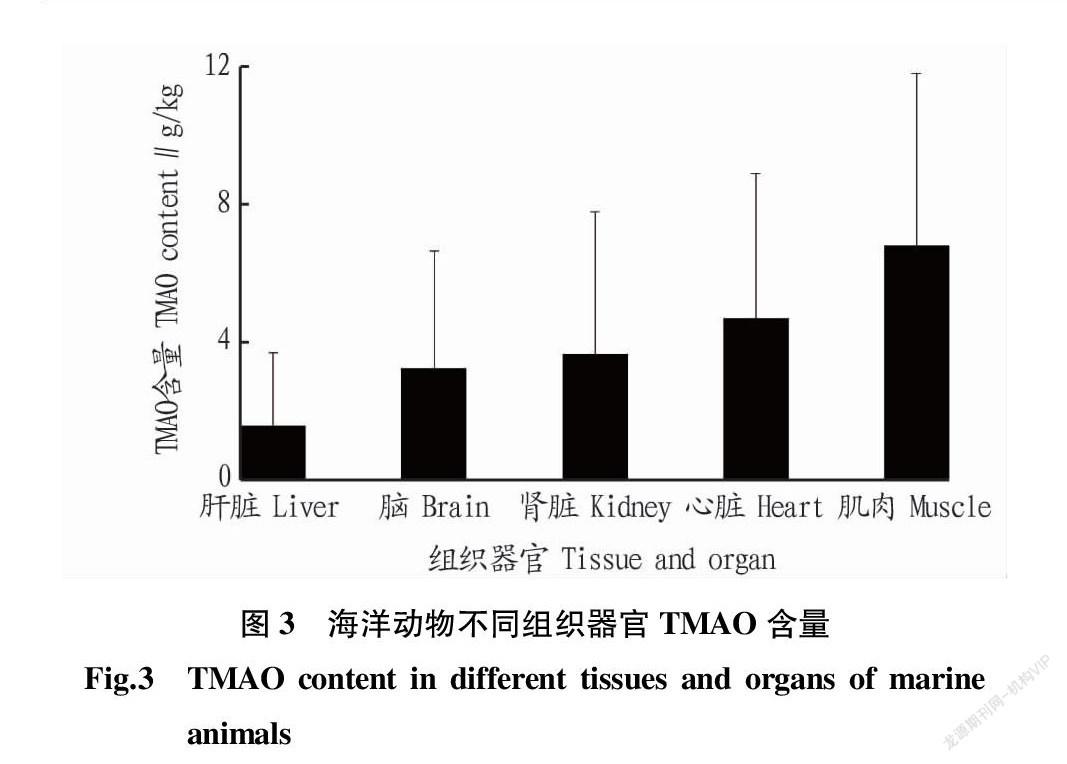
TMAO和GBT在海洋动物体内的分布并不均勻(表3),整体呈现肌肉>心脏>肾脏≈脑>肝脏的趋势(图3)。海洋动物肌肉中TMAO平均含量为6.80 g/kg,其中黑霞鲨(Centroscyllium fabricii)肌肉中TMAO含量达18.63 g/kg[63]。海洋动物心脏中含量次之(4.70 g/kg),黑霞鲨心脏中TMAO含量可达13.52 g/kg[63]。而在肾脏和脑组织中TMAO含量差别不大,其平均含量分别为3.66和3.25 g/kg。肝脏含量最低,TMAO含量为1.55 g/kg。
关于海洋动物不同组织内GBT含量的报道较少,Treberg等[49]报道冬鳐鱼(Leucoraja ocellata)心脏中GBT含量高达3.16 g/kg,显著高于其他组织器官,角鲨组织器官中GBT也主要聚积在心脏。
3 海洋动物体内TMAO和GBT含量的主要影响因素
3.1 深度
海洋动物体内TMAO含量受环境深度的影响较大,主要因为TMAO具有增加海洋动物浮力的作用。海洋动物所处环境越深,外部的压力越大,从而促使动物体内积累更高浓度的TMAO以应对环境压力[ 3 67]。Yancey等[68]在对太平洋几种硬骨鱼类的研究发现,随着水深从0到1 400 m逐渐增加时,其肌肉中TMAO含量从3.00 g/kg增加至11.25 g/kg。Keller等[69]通过对不同深度下同一物种中TMAO含量进行检测发现,深海(4 850 m)硬骨鱼肌肉中TMAO的含量是近岸浅水区个体的5倍。在大西洋4 800 m深处的长尾鳕鱼肌肉中TMAO含量高达19.58 g/kg[70]。在水深7 000 m的克马德里海沟,黑线鳕体内TMAO含量高达28.95 g/kg,几乎是透光区鱼类平均含量的8倍[71]。
通过对28种软骨鱼及58种硬骨鱼体内TMAO含量及其所栖息的深度分析发现(图4),这2类鱼类体内TMAO含量随深度的增加而显著增高[3 49,67,69-70]。通过对<500 m、500~2 000 m和>2 000 m 3种水深中的5大類(软骨鱼类、头足类、甲壳类、硬骨鱼类和贝类)动物统计分析发现(图5),当水深<500 m时,软骨鱼类体内TMAO含量最高(为5.63 g/kg),是其他4类(头足类、甲壳类、硬骨鱼类和贝类)的3.1~5.8倍;当水深在500~2 000 m时,依然是软骨鱼类体内TMAO含量最高,而甲壳类和头足类含量相近,约占软骨鱼类的75%,硬骨鱼类含量最低,约为软骨鱼类的47%;当水深>2 000 m时,甲壳类TMAO平均含量为16.49 g/kg,与软骨鱼类和头足类含量相差不大,是硬骨鱼类的1.9倍。对于同种类型的动物来说,水深>2 000 m的软骨鱼类体内TMAO含量分别是500~2 000 m和<500 m的1.4和2.7倍,硬骨鱼类分别为1.6和5.6倍,甲壳类动物分别为2.0和9.0倍,头足类动物为2.3和12.4倍。说明头足类动物体内TMAO随深度变化最显著,硬骨鱼类、甲壳类次之,软骨鱼类变化最小。
并非所有报道都表明TMAO含量随深度增加而增加,Laxson等[ 17]研究表明TMAO含量不会随着深度的变化而发生变化。这也说明不同海洋动物对深度的敏感性存在差异。
3.2 盐度
TMAO是海洋动物重要的渗透调节物质,盐度也会影响海洋动物体内TMAO含量。广盐性白真鲨从淡水中放入海水中,体内TMAO含量增加到之前的1.5倍[72]。当盐度减小为80%时,广盐性鳐鱼体内TMAO含量降低了5%;当盐度减小为50%时,TMAO的含量降低了36%[73]。Summers等[18]研究发现,当海洋鱼类进入到低盐度河口水域时,其体内TMAO含量往往很低。将鳉鱼(Killifish)从100%的海水转移到淡水时,也观察到其肌肉中TMAO含量下降了46%[74]。在对白斑角鲨(Squalus acanthias)的研究中发现,随着环境盐度降低至70%时,白斑角鲨的腮中TMAO含量会下降至原来的85%[65]。Chung等[75]通过对香港89种(共266条)淡水鱼、海水和淡水两栖的鱼、海水鱼体内的TMAO分析发现,75%的海水和淡水两栖的鱼中可检测出TMAO,而仅有33%的淡水鱼中可检测出TMAO。因此,海洋动物为应对较高盐度压力,其体内会积累更多的TMAO。
盐度对海洋动物体内GBT含量也具有显著的影响。海洋动物在高盐度环境中积累大量GBT,并在盐度降低时释放。一般情况下,海洋动物体内GBT含量是淡水动物的10~100倍[76]。Brictkux-Grégoire等[77]研究发现,盐度下降50%时,贻贝的肌肉中GBT含量会降低35%。Delgado-Gaytán等[78]研究也表明,与对照组盐度(35‰)相比,在40‰、50‰和60‰的盐度下,虾鳃中GBT含量高出7.3~8.3倍,而在肌肉中GBT含量则高出9倍。因此,为适应盐度的压力,海洋动物体内TMAO或GBT含量远高于淡水动物[57,65]。
3.3 温度
TMAO和GBT作为海洋动物重要的冷冻保护剂,其含量也会受温度的影响。Treberg等[33]的室内试验显示,低温可以诱导胡瓜鱼(Osmerus mordaxa)体内TMAO的积聚。在格陵兰鳕鱼(Gadus ogac)等一些冷水动物的血清和肝脏中,TMAO水平异常高(0.60~2.10 g/kg)[79-81]。在夏季温度较高时,长鳍线指鰕翁(Nemadactylus macropterus)和绿鳍鱼(Chelidonichthys kumu)TMAO含量会降低[18]。在冬季温度较低时,三文鱼(Oncorhynchus)的血清、彩虹鱼(Poecilia)的血浆和鲱鱼(Herring)的肌肉中TMAO含量都有所增加[6 79,82]。林海生等[66]研究发现我国南北方海域牡蛎中GBT含量呈现“南低北高”的特征,这可能是由于北方温度较南方低造成的。因此,温度较低时,海洋动物体内会产生较多的TMAO或GBT以适应温度较低的环境[3 67]。
4 展望
综上所述,虽然目前国内外对海洋动物体内TMAO和GBT的合成与降解途径、浓度特征及影响因素进行了较多研究,但还有许多问题需要解决,具体体现在以下3点:
①现阶段国内外对海洋动物体内TMAO开展了一定研究,但对GBT报道较少,未来需加强在海洋动物体内GBT的代谢机制、不同组织的浓度特征及影响因素等方面的研究;
②揭示海洋动物体内TMAO和GBT的转化机制,阐明不同种类海洋动物体内两者浓度存在差异的原因;
③我国是世界海水养殖第一大国,进一步探索我国大规模海水养殖生物体内TMAO和GBT对水体及大气中有机胺的贡献及其潜在的气候效应。
参考文献
[1] SELLEGRI K,PEY J,ROSE C,et al.Evidence of atmospheric nanoparticle formation from emissions of marine microorganisms[J].Geophysical research letters,201 43(12):6596-6603.
[2] ALMEIDA J,SCHOBESBERGER S,KRTEN A,et al.Molecular understanding of sulphuric acid-amine particle nucleation in the atmosphere[J].Nature,201 502(7471):359-363.
[3] CHEN H H,VARNER M E,GERBER R B,et al.Reactions of methanesulfonic acid with amines and ammonia as a source of new particles in air[J].The journal of physical chemistry B,201 120(8):1526-1536.
[4] YAO L,GARMASH O,BIANCHI F,et al.Atmospheric new particle formation from sulfuric acid and amines in a Chinese megacity[J].Science,2018,361(6399):278-281.
[5] LAXSON C J,CONDON N E,DRAZEN J C,et al.Decreasing urea:Trimethylamine N-oxide ratios with depth in chondrichthyes:A physiological depth limit?[J].Physiological & biochemical zoology,201 84(5):494-505.
[6] HATTON A D,GIBB S W.A technique for the determination of trimethylamine-N-oxide in natural waters and biological media[J].Analytical chemistry,1999,71(21):4886-4891.
[7] CARPENTER L J,ARCHER S D,BEALE R.Ocean-atmosphere trace gas exchange[J].Chemical society reviews,201 41(19):6473-6506.
[8] OREN A.Cyanobacteria in hypersaline environments:Biodiversity and physiological properties[J].Biodiversity and conservation,201 24(4):781-798.
[9] CHARLOTTE C.Distributions of glycine betaine and the methylamines in coastal waters:Analytical developments and a seasonal study[D].Devon,UK:University of Plymouth,2015.
[10] SUN Y,DING S S,HE M W,et al.Construction and analysis of the immune effect of Vibrio harveyi subunit vaccine and DNA vaccine encoding TssJ antigen[J].Fish and shellfish immunology,2020,98:45-51.
[11] GE X L,WEXLER A S,CLEGG S L.Atmospheric amines-Part I.A review[J].Atmospheric environment,201 45(3):524-546.
[12] GIBB S W,MANTOURA R F C,LISS P S.Ocean-atmosphere exchange and atmospheric speciation of ammonia and methylamines in the region of the NW Arabian Sea[J].Global biogeochemical cycles,1999,13(1):161-178.
[13] KING G M.Methanogenesis from methylated amines in a hypersaline algal mat[J].Applied and environmental microbiology,1988,54(1):130-136.
[14] VAN PINXTEREN M,FOMBA K W,VAN PINXTEREN D,et al.Aliphatic amines at the Cape Verde Atmospheric Observatory:Abundance,origins and sea-air fluxes[J].Atmospheric environment,2019,203(APR):183-195.
[15] HU Q J,YU P R,ZHU Y J,et al.Concentration,size distribution and formation of trimethylaminium and dimethylaminium ions in atmospheric particles over marginal seas of China[J].Journal of the atmospheric sciences,201 72(9):3487-3498.
[16] YU P R,HU Q J,LI K,et al.Characteristics of dimethylaminium and trimethylaminium in atmospheric particles ranging from supermicron to nanometer sizes over eutrophic marginal seas of China and oligotrophic open oceans[J].Science of the total environment,201 572:813-824.
[17] SEIBEL B A,WALSH P J.Trimethylamine oxide accumulation in marine animals:Relationship to acylglycerol storage[J].Journal of experimental biology,200 205(Pt 3):297-306.
[18] SUMMERS G,WIBISONO R D,HEDDERLEY D I,et al.Trimethylamine oxide content and spoilage potential of New Zealand commercial fish species[J].New Zealand journal of marine and freshwater research,2017,51(3):393-405.
[19] PIERCE S K,DRAGOLOVICH J,CROMBIE B N.Variations in intracellular choline levels may account for differences in glycine betaine synthesis between conspecific oyster populations responding to hyperosmotic stress[J].Journal of experimental zoology,1997,278(5):283-289.
[20] BEATTY S A.Studies of fish spoilage:II.The origin of trimethylamine produced during the spoilage of cod muscle press juice[J].Journal of the fisheries research board of Canada,1938,4(2):63-68.
[21] NAMIES'NIK J,JASTRZEBSKA A,ZYGMUNT B.Determination of volatile aliphatic amines in air by solid-phase microextraction coupled with gas chromatography with flame ionization detection[J].Journal of chromatography A,200 1016(1):1-9.
[22] IAN L.Microbial methylated amine metabolism in marine surface waters[D].Coventry,UK:University of Warwick,2015.
[23] GREEN T K,HATTON A D.The claw hypothesis:A new perspective on the role of biogenic sulphur in the regulation of global climate[M]//HUGHES R N,HUGHES D J,SMITH I P.Oceanography and marine biology:Volume 52.Boca Raton,FL:CRC Press,2014:315-336.
[24] WELSH D T.Ecological significance of compatible solute accumulation by micro-organisms:From single cells to global climate[J].FEMS Microbiology Reviews,2000,24(3):263-290.
[25] TORSTENSSON A,YOUNG J N,CARLSON L T,et al.Use of exogenous glycine betaine and its precursor choline as osmoprotectants in Antarctic sea-ice diatoms[J].Journal of phycology,2019,55(3):663-675.
[26] WITHERS P C,HEFTER G T,PANG T S.Role of urea and methylamines in buoyancy of elasmobranchs[J].Journal of experimental biology,199 188(1):175-189.
[27] YANCEY P H,CLARK M E,HAND S C,et al.Living with water stress:Evolution of osmolyte systems[J].Science,198 217(4566):1214-1222.
[28] CHOLETTE C,GAGNON A.Isosmotic adaptation in Myxine glutinosa L.-II.Variations of the free amino acids,trimethylamine oxide and potassium of the blood and muscle cells[J].Comparative biochemistry & physiology part A:Physiology,197 45(4):1009-1021.
[29] FORSTER R P,GOLDSTEIN L.Intracellular osmoregulatory role of amino acids and urea in marine elasmobranchs[J].The American journal of physiology,197 230(4):925-931.
[30] YANCEY P H,SIEBENALLER J F.Trimethylamine oxide stabilizes teleost and mammalian lactate dehydrogenases against inactivation by hydrostatic pressure and trypsinolysis[J].The journal of experimental biology,1999,202(Pt24):3597-3603.
[31] QU Y X,BOLEN D W.Hydrogen exchange kinetics of RNase A and the urea:TMAO paradigm[J].Biochemistry,200 42(19):5837-5849.
[32] BASKAKOV I,WANG A J,BOLEN D W.Trimethylamine-N-oxide counteracts urea effects on rabbit muscle lactate dehydrogenase function:A test of the counteraction hypothesis[J].Biophysical journal,1998,74(5):2666-2673.
[33] TREBERG J R,BYSTRIANSKY J S,DRIEDZIC W R.Temperature effects on trimethylamine oxide accumulation and the relationship between plasma concentration and tissue levels in smelt(Osmerus mordax)[J].Journal of experimental zoology part A:Comparative experimental biology,200 303(4):283-293.
[34] VILLALOBOS A R A,RENFRO J L.Trimethylamine oxide suppresses stress-induced alteration of organic anion transport in choroid plexus[J].Journal of experimental biology,2007,210(Pt3):541-552.
[35] GILLETT M B,SUKO J R,SANTOSO F O,et al.Elevated levels of trimethylamine oxide in muscles of deep-sea gadiform teleosts:A high-pressure adaptation?[J].Journal of experimental zoology,1997,279(4):386-391.
[36] YANCEY P H,SOMERO G N.Methylamine osmoregulatory solutes of elasmobranch fishes counteract urea inhibition of enzymes[J].The journal of experimental zoology,1980,212(2):205-213.
[37] TRZECIAKOWSKI J P.Analysis of stimulus-response chains using nonlinear dynamics[J].Journal of pharmacological & toxicological methods,199 36(2):103-121.
[38] RAYMOND J A.Trimethylamine oxide and urea synthesis in rainbow smelt and some other northern fishes[J].Physiological zoology,1998,71(5):515-523.
[39] SCHLENK D.Occurrence of flavin-containing monooxygenases in non-mammalian eukaryotic organisms[J].Comparative biochemistry and physiology.Part C:Pharmacology,toxicology & endocrinology,1998,121(1/2/3):185-195.
[40] USSHER J R,LOPASCHUK G D,ARDUINI A.Gut microbiota metabolism of l-carnitine and cardiovascular risk[J].Atherosclerosis,201 231(2):456-461.
[41] UFNAL M,ZADLO A,OSTASZEWSKI R.TMAO:A small molecule of great expectations[J].Nutrition,201 31(11/12):1317-1323.
[42] BAKER J R,CHAYKIN S.The biosynthesis of trimethylamine-N-oxide[J].The journal of biological chemistry,196 237(3):1309-1313.
[43] DE VOOYS C G N,GEENEVASEN J A J.Biosynthesis and role in osmoregulation of glycine-betaine in the Mediterranean mussel Mytilus galloprovincialis LMK[J].Comparative biochemistry and physiology part B,200 132(2):409-414.
[44] VAN WAARDE A.Biochemistry of non-protein nitrogenous compounds in fish including the use of amino acids for anaerobic energy production[J].Comparative biochemistry and physiology part B:Comparative biochemistry,1988,91(2):207-228.
[45] TOKUNAGA T.Trimethylamine oxide and its decomposition in the bloody muscle of fish.I.TMAO,TMA,and DMA contents in ordinary and bloody muscles[J].Nippon suisan gakkaishi,1970,36(5):502-509.
[46] NIIZEKI N,DAIKOKU T,HIRATA T,et al.Mechanism of biosynthesis of trimethylamine oxide in tilapia reared under seawater conditions[J].Fisheries science,200 69(1):74-87.
[47] DELGADO-GAYTN M F,ROSAS-RODRGUEZ J A,YEPIZ-PLASCENCIA G,et al.Cloning and molecular characterization of the betaine aldehyde dehydrogenase involved in the biosynthesis of glycine betaine in white shrimp(Litopenaeus vannamei)[J].Chemico-biological interactions,2017,276:65-74.
[48] PIERCE S K,ROWLAND-FAUX L M,O′BRIEN S M.Different salinity tolerance mechanisms in Atlantic and Chesapeake Bay conspecific oysters:Glycine betaine and amino acid pool variations[J].Marine biology,199 113(1):107-115.
[49] TREBERG J R,DRIEDZIC W R.The accumulation and synthesis of betaine in winter skate(Leucoraja ocellata)[J].Comparative biochemistry and physiology part A,2007,147(2):475-483.
[50] CHISTOSERDOVA L.Modularity of methylotrophy,revisited[J].Environmental microbiology, 201 13(10):2603-2622.
[51] 付雪媛,劉丰海,姜城子,等.高效液相色谱-质谱联用检测水产品中氧化三甲胺[J].食品安全质量检测学报,201 7(1):269-275.
[52] 蔡英华,舒绪刚,廖列文,等.氧化三甲胺在动物养殖中的应用研究进展[J].广东饲料,2009,18(3):32-35.
[53] DEHAUT A,DUTHEN S,GRARD T,et al.Development of an SPME-GC-MS method for the specific quantification of dimethylamine and trimethylamine:Use of a new ratio for the freshness monitoring of cod fillets[J].Journal of the science of food and agriculture,201 96(11):3787-3794.
[54] NITISEWOJO P,HULTIN H O.Characteristics of TMAO degrading systems in Atlantic short finned squid(Illex illecebrosus)[J].Journal of food biochemistry,198 10(2):93-106.
[55] LUNDSTROM R C,CORREIA F F,WILHELM K A.Enzymatic dimethylamine and formaldehyde production in minced American plaice and blackback flounder mixed with a red hake TMAO-ase active fraction[J].Journal of food science,198 47(4):1305-1310.
[56] CHAREST R P,CHENOWETH M,DUNN A.Metabolism of trimethylamines in kelp bass(Paralabrax clathratus)and marine and freshwater pink salmon(Oncorhynchus gorbuscha)[J].Journal of comparative physiology B,1988,158(5):609-619.
[57] ANTHONI U,B RRESEN T,CHRISTOPHERSEN C,et al.Is trimethylamine oxide a reliable indicator for the marine origin of fish?[J].Comparative biochemistry and physiology B:Comparative biochemistry,1990,97(3):569-571.
[58] LI F,LIU H Y,XUE C H,et al.Simultaneous determination of dimethylamine,trimethylamine and trimethylamine-n-oxide in aquatic products extracts by ion chromatography with non-suppressed conductivity detection[J].Journal of chromatography A,2009,1216(31):5924-5926.
[59] BOCKUS A B,SEIBEL B A.Synthetic capacity does not predict elasmobranchs' ability to maintain trimethylamine oxide without a dietary contribution[J].Comparative biochemistry and physiology part A,2018,217:35-42.
[60] KELLY R H,YANCEY P H.High contents of trimethylamine oxide correlating with depth in deep-sea teleost fishes,skates,and decapod crustaceans[J].The biological bulletin,1999,196(1):18-25.
[61] 姜城子,崔潔,周苗苗,等.青岛地区部分水产品中氧化三甲胺含量的测定[J].食品安全质量检测学报,201 5(1):41-46.
[62] BEDFORD J J,HARPER J L,LEADER J P,et al.Betaine is the principal counteracting osmolyte in tissues of the elephant fish,Callorhincus millii(Elasmobranchii,Holocephali)[J].Comparative biochemistry and physiology part B,1998,119(3):521-526.
[63] TREBERG J R,DRIEDZIC W R.Elevated levels of trimethylamine oxide in deep-sea fish:Evidence for synthesis and intertissue physiological importance[J].The journal of experimental zoology,200 293(1):39-45.
[64] 陈帅,朱军莉,励建荣.非抑制离子色谱检测海产品中氧化三甲胺、三甲胺和二甲胺[J].中国食品学报,201 13(4):163-171.
[65] MACLELLAN R J,TUNNAH L,BARNETT D,et al.Chaperone roles for TMAO and HSP70 during hyposmotic stress in the spiny dogfish shark(Squalus acanthias)[J].Journal of comparative physiology B,201 185(7):729-740.
[66] 林海生,秦小明,章超桦,等.中国沿海主要牡蛎养殖品种的营养品质和风味特征比较分析[J].南方水产科学,2019,15(2):110-120.
[67] DOWNING A B,WALLACE G T,YANCEY P H.Organic osmolytes of amphipods from littoral to hadal zones:Increases with depth in trimethylamine N-oxide,scyllo-inositol and other potential pressure counteractants[J].Deep sea research part I:Oceanographic research papers,2018,138:1-10.
[68] YANCEY P H,RHEA M D,KEMP K M,et al.Trimethylamine oxide,betaine and other osmolytes in deep-sea animals:Depth trends and effects on enzymes under hydrostatic pressure[J].Cellular and molecular biology,200 50(4):371-376.
[69] KELLER M D,KIENE R P,MATRAI P A,et al.Production of glycine betaine and dimethylsulfoniopropionate in marine phytoplankton.II.N-limited chemostat cultures[J].Marine biology,1999,135(2):249-257.
[70] SAMEROTTE A L,DRAZEN J C,BRAND G L,et al.Correlation of trimethylamine oxide and habitat depth within and among species of teleost fish:An analysis of causation[J].Physiological and biochemical zoology,2007,80(2):197-208.
[71] YANCEY P H,GERRINGER M E,DRAZEN J C,et al.Marine fish may be biochemically constrained from inhabiting the deepest ocean depths[J].Proceedings of the national academy of sciences of the United States of America,201 111(12):4461-4465.
[72] PILLANS R D,GOOD J P,ANDERSON W G,et al.Freshwater to seawater acclimation of juvenile bull sharks(Carcharhinus leucas):Plasma osmolytes and Na+/K+-ATPase activity in gill,rectal gland,kidney and intestine[J].Journal of comparative physiology B,200 175(1):37-44.
[73] SULIKOWSKI J A,TREBERG J R,HOWELL W H.Fluid regulation and physiological adjustments in the winter skate,Leucoraja ocellata,following exposure to reduced environmental salinities[J].Environmental biology of fishes,200 66(4):339-348.
[74] OGILVIE J M G,WARREN A A.The occurrence of trimethylamine oxide in fundulus heteroclitus[J].Canadian journal of zoology,1957,35(6):735-745.
[75] CHUNG S W C,CHAN B T P.Trimethylamine oxide,dimethylamine,trimethylamine and formaldehyde levels in main traded fish species in Hong Kong[J].Food additives & contaminants part B,2009,2(1):44-51.
[76] CLOWES L A,FRANCESCONI K A.Uptake and elimination of arsenobetaine by the mussel Mytilus edulis is related to salinity[J].Comparative biochemistry and physiology part C,200 137(1):35-42.
[77] BRICTKUX-GRGOIRE S,DUCHTEAU-BOSSON G,FLORKIN C J M.Constituants osmotiquement actifs des muscles adducteurs de mytilus edulis adaptée a l′eau de mer ou a l′eau saumatre[J].Archives of physiology and biochemistry,196 72(1):116-123.
[78] DELGADO-GAYTN M F,GMEZ-JIMNEZ S,GMEZ-ALEJO L A,et al.Effect of salinity on the synthesis and concentration of glycine betaine in osmoregulatory tissues from juvenile shrimps Litopenaeus vannamei[J].Comparative biochemistry and physiology part A:Molecular & integrative physiology,2019,240:110628.
[79] RAYMOND J A.Seasonal variations of trimethylamine oxide and urea in the blood of a cold-adapted marine teleost,the rainbow smelt[J].Fish physiology and biochemistry,199 13(1):13-22.
[80] GORDON M S,AMDUR B H,SCHOLANDER P F.Freezing resistance in some northern fishes[J].Biological bulletin,196 122(1):52-62.
[81] DEVRIES A L,WOHLSCHLAG D E.Freezing resistance in some Antarctic fishes[J].Science,1969,163(3871):1073-1075.
[82] HUGHES R B.Chemical studies on the herring(Clupea harengus).II.—The free amino-acids of herring flesh and their behaviour during post-mortem spoilage[J].Journal of the science of food and agriculture,1959,10(10):558-564.
[83] 李丰.水产品中氧化三甲胺、三甲胺、二甲胺检测方法及鱿鱼丝中甲醛控制研究[D].保定:河北农业大学,2010:22.
[84] BOCKUS A B,SEIBEL B A.Trimethylamine oxide accumulation as a function of depth in Hawaiian mid-water fishes[J].Deep-sea research part I,201 112:37-44.
[85] DACOSTA K A,VRBANAC J J,ZEISEL S H.The measurement of dimethylamine,trimethylamine,and trimethylamine N-oxide using capillary gas chromatography-mass spectrometry[J].Analytical biochemistry,1990,187(2):234-239.
[86] AGU'STSSON I,STR M A R.Biosynthesis and turnover of trimethylamine oxide in the teleost cod,Gadus morhua[J].The journal of biological chemistry,198 256(15):8045-8049.
[87] 黄国霞,赖春华,李军生,等.6种水产动物中氧化三甲胺的提取與含量测定[J].食品科技,201 37(7):305-307.
[88] STIBOLLER M,RABER G,FRANCESCONI K A.Simultaneous determination of glycine betaine and arsenobetaine in biological samples by HPLC/ICPMS/ESMS and the application to some marine and freshwater fish samples[J].Microchemical journal,201 122:172-175.
[89] DE ZWART F J,SLOW S,PAYNE R J,et al.Glycine betaine and glycine betaine analogues in common foods[J].Food chemistry,200 83(2):197-204.
[90] 陈德慰,苏键,颜栋美,等.广西北部湾常见水产品中甜菜碱含量测定及呈味效果评价[J].现代食品科技,201 27(4):468-472.
[91] 丁源,邢家溧,承海,等.基于高效液相色谱法测定水产品中甘氨酸甜菜碱[J].食品工业科技,2019,40(15):184-187.
[92] FARABEGOLI F,ZIRONI E,GAZZOTTI T,et al.Isotope dilution LC-MS/MS method for glycine betaine in manila clam(Tapes philippinarum)[J].Food analytical methods,2019,12(6):1448-1455.
[93] 王士稳,梁萌青,林洪,等.海水和淡水养殖凡纳滨对虾呈味物质的比较分析[J].海洋水产研究,200 27(5):79-84.
[94] 金蕾,徐善良,邱成功,等.三疣梭子蟹肌肉组织中甜菜碱、糖原及无机盐变化研究[J].生物学杂志,201 31(4):24-28.
[95] TIMM M,J RGENSEN B M.Simultaneous determination of ammonia,dimethylamine,trimethylamine and trimethylamine-n-oxide in fish extracts by capillary electrophoresis with indirect UV-detection[J].Food chemistry,200 76(4):509-518.
[96] BEDFORD J J,HARPER J L,LEADER J P,et al.Identification and measurement of methylamines in elasmobranch tissues using proton nuclear magnetic resonance(1H-NMR)spectroscopy[J].Journal of comparative physiology B,1998,168(2):123-131.
猜你喜欢
趣味(数学)(2021年9期)2022-01-19
环境保护与循环经济(2017年4期)2018-01-22
中成药(2017年4期)2017-05-17
中国市场(2016年36期)2016-10-19
商(2016年27期)2016-10-17
商(2016年27期)2016-10-17
科技视界(2016年20期)2016-09-29
中国洗涤用品工业(2016年2期)2016-02-28
安徽医药(2014年9期)2014-03-20
