
基于细胞线粒体能量代谢研究甘草次酸拮抗乌头碱的心肌毒性作用
2021-11-29 17:55何苗,李耀伟,王志琪,周志华,蒲科婷,丛梦静,王嘉星
湖南中医药大学学报 2021年11期
何苗,李耀伟,王志琪,周志华,蒲科婷,丛梦静,王嘉星

〔摘要〕 目的 从线粒体能量代谢角度研究甘草次酸对乌头碱的心肌细胞毒性拮抗作用。方法 将大鼠心肌细胞H9c2细胞随机分为10组,即空白组、甘草次酸组、4个乌头碱组(乌头碱120、240、480、960 μmol/L)、4个乌头碱配伍甘草次酸组(不同浓度乌头碱分别与60 μmol/L甘草次酸配伍)。采用噻唑蓝(methyl thiazolyl tetrazolium, MTT)法测定不同浓度给药组对H9c2细胞的影响;采用ELISA法测定各组细胞三磷酸腺苷(adenosine 5-triphosphate, ATP)含量、ATP酶活性水平;采用活性氧探针(2,7-dichlorodi-hydrofluorescein diacetate, DCFH-DA)检测细胞内活性氧(reactive oxygen species, ROS)、线粒体膜电位探针(5,5′6,6′-tetrachloro-1,1′,3,3′-tetraethyl-imidacarbocyanine iodide, JC-1)检测细胞内线粒体膜电位、钙离子荧光探针(rhod-2 acetoxymethyl ester,Rhod-2AM)检测细胞内钙离子浓度。结果 与单用乌头碱比较,乌头碱配伍甘草次酸后心肌细胞的ATP含量、Ca2+-ATP酶、Ca2+-Mg2+-ATP酶、Na+-K+-ATP酶活性升高,ROS含量减少,线粒体膜电位、线粒体内钙离子浓度降低(P<0.01或P<0.05)。结论 甘草次酸可拮抗乌头碱的心肌细胞毒性,其机制可能是与保护心肌细胞线粒体、增强线粒体能量代谢、维持钙稳态有关。
〔关键词〕 线粒体能量代谢;乌头碱;甘草次酸;心肌毒性;钙稳态
〔中图分类号〕R285.5 〔文獻标志码〕A 〔文章编号〕doi:10.3969/j.issn.1674-070X.2021.11.002
Glycyrrhetinic Acid Against the Cardiotoxicity of Aconitine Based on Mitochondrial
Energy Metabolism
HE Miao1,2, LI Yaowei1, WANG Zhiqi2,3*, ZHOU Zhihua1, PU Keting3, CONG Mengjing3, WANG Jiaxing3
(1. Graduate School, Hunan University of Chinese Medicine, Changsha, Hunan 410208, China; 2. Research Center of Standardization and Functional Engineering of Traditional Chinese Medicine in Hunan Province, Changsha, Hunan 410208, China; 3. School of Pharmacy, Hunan University of Chinese Medicine, Changsha, Hunan 410208, China)
〔Abstract〕 Objective To explore the antagonistic effect of glycyrrhetinic acid on cardiomyocytes cytotoxicity of aconitine from the perspective of mitochondrial energy metabolism. Methods The H9c2 cells of rat cardiomyocytes were randomly divided into 10 groups: blank group, glycyrrhetinic acid group, 4 aconitine groups (aconitine 120, 240, 480, 960 μmol/L), and 4 aconitine combined with glycyrrhetinic acid groups (aconitine in different concentrations was compatible with 60 μmol/L glycyrrhetinic acid). The effect of different concentrations on the H9c2 cell was determine by methyl thiazolyl tetrazolium (MTT). The content of adenosine5-triphosphate (ATP) and the level of ATP enzyme activity in each group were measured by ELISA. The intracellular reactive oxygen species (ROS) was detected by 2,7-dichlorodi-hydrofluorescein diacetate (DCFH-DA), 5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethyl-imidacarbocyanine iodide (JC-1) was used to detect intracellular mitochondrial membrane potential and rhod-2 acetoxymethyl ester (Rhod-2AM) probe was used to detect intracellular calcium ion concentration. Results Compared with aconitine alone, the ATP content, Ca2+-ATPase, Ca2+-Mg2+-ATPase and Na+-K+-ATPase activities of cardiomyocytes were significantly increased after aconitine combined with glycyrrhetinic acid. The ROS content, mitochondrial membrane potential and intracellular calcium concentration were significantly decreased (P<0.01 or P<0.05). Conclusion Glycyrrhetinic acid can reduce the cardiotoxicity of aconitine, which may be related to the protection of mitochondria of cardiomyocytes, the enhancement of mitochondrial energy metabolism and the maintenance of calcium homeostasis.
〔Keywords〕 mitochondrial energy metabolism; aconitine; glycyrrhetinic acid; cardiotoxicity; calcium homeostasis
附子为典型的毒效共存中药,配伍是降低其毒性常用的方法之一[1-2]。在“中药方剂数据库”网站检索发现:约有96%含乌头类药材(附子或川乌、草乌)方剂中同时含甘草[3]。課题组前期研究[4-6]表明,附子单煎液对离体心衰蛙心及大鼠心脏都有强心作用;与甘草配伍后,其强心作用较附子单煎液弱;乌头碱和甘草次酸Tmax均推迟,Cmax均降低,吸收均受到抑制,t1/2均缩短[7]。
一般认为,附子的主要毒性成分是乌头类生物碱,其中毒性较强是乌头碱,主要影响心血管、神经等系统[8],心脏是其毒性作用的主要靶器官[9],大剂量时会诱发心律失常。研究还发现,乌头碱的心肌毒性可表现为影响心肌细胞能量代谢,配伍甘草次酸后可有效拮抗乌头碱引发的心律失常[10]。线粒体是心肌细胞中主要负责能量代谢的细胞器,膜电位、三磷酸腺苷(adenosine 5-triphosphate, ATP)、Ca2+浓度等是表征相关功能的经典指标[11],且其正常的能量代谢功能与细胞内钙稳态密切相关[12]。当心脏组织受到损伤时,心肌细胞内活性氧(reactive oxygen species, ROS)释放增多、Ca2+浓度失衡、能量代谢紊乱[13]。
故本研究以心肌细胞线粒体能量代谢为切入点,以ATP含量、线粒体膜电位、ROS浓度、线粒体Ca2+浓度、Ca2+-ATP酶、Ca2+-Mg2+-ATP酶、Na+-K+-ATP酶活性为指标,在细胞水平对甘草次酸拮抗乌头碱心肌毒性作用进行研究。
1 材料
1.1 实验细胞系
大鼠心肌细胞(H9c2)购自普诺赛生物公司,货号:CL-0089。将H9c2细胞以含10%胎牛血清的DMEM培养基、5% CO2、37 ℃条件下进行培养,待细胞密度达90%左右后0.25%胰酶消化传代,平均2~3 d一代,取对数生长期细胞进行实验。
1.2 实验药物
乌头碱(aconitine, AC)储备液:将20 mg乌头碱溶于150 μL DMSO中,而后将含乌头碱的DMSO溶解于31 mL空白DMEM培养基中,涡旋混匀,使之成为1 mmol/L的乌头碱储备液,以0.22 μm针头式一次性过滤器除菌后分装于15 mL的离心管中,-20 ℃条件下保存备用。
甘草次酸(alycyrrhizic acid, GA)储备液:将20 mg甘草次酸溶于200 μL DMSO中,而后将含甘草次酸的DMSO溶解于21 mL空白DMEM培养基中,涡旋混匀,使之成为2 mmol/L的甘草次酸储备液,以0.22 μm针头式一次性过滤器除菌后分装于15 mL的离心管中,-20 ℃条件下保存备用。
噻唑蓝(methyl thiazolyl tetrazolium, MTT)储备液:将250 mg MTT溶解于50 mL PBS中,涡旋混匀,使之成为5 mg/mL的MTT储备液,以0.22 μm针头式一次性过滤器除菌后分装于15 mL的离心管中,-20 ℃条件下保存备用。
1.3 主要试剂与仪器
特级胎牛血清(批号:124210-500)、DMEM高糖培养基(批号:PM150210)、PBS缓冲液(批号:PB180327)、胰蛋白酶-EDTA消化液(批号:PB180226)、无血清非程序冻存液(批号:PB180438 )、D-Hanks液(批号:PB180321)均购于普诺赛生命科技有限公司;噻唑蓝(MTT)(批号:M8180)、二甲基亚砜(DMSO)(培养级,批号:D8371)、线粒体膜电位试剂盒(批号:M8650)、青链霉素混合液(批号:P1400)均购于索莱宝生物科技有限公司;Hanks平衡盐溶液(HBSS)(批号:M120702)购于上海源培生物科技股份有限公司;二甲基亚砜(DMSO)(分析级,批号:D806645)、乌头碱对照品(批号:DST200729-006,纯度≥98%)、甘草次酸对照品(批号:DST200619-007,纯度≥98%)均购于德斯特生物技术有限公司;ATP含量测定试剂盒(批号:A095-2-1)、Ca2+-ATP酶试剂盒(批号:A070-4-1)、Na+-K+-ATP酶试剂盒(批号:A070-2-2)、Ca2+-Mg2+-ATP酶试剂盒(批号:A070-3-2)、ROS测定试剂盒(批号:E004-1-1)均购于南京建成生物科技研究所;Rhod-2 AM钙荧光探针(批号:21060)购于美国AAT BIOQUES公司。
HERACELL 150i三气培养箱(赛默飞世尔科技有限公司);SW-CJ洁净工作台(苏州安泰空气技术有限公司);BX51倒置显微镜(日本OLYMPUS公司);IX51荧光倒置显微镜(日本OLYMPUS公司);TS2荧光倒置显微镜(尼康光学仪器中国有限公司);HH.S21-8恒温水浴锅(上海博迅实业有限公司医疗设备厂);5180R高速离心机(德国EPPENDORF公司);TG16W医用离心机(长沙平凡仪器仪表有限公司);XW-18D旋涡混合器(绍兴市苏珀仪器有限公司);F570-86 -80 ℃超低温冰箱(德国EPPENDORF公司);BCD-258WDPM医用冷藏冷冻箱(青岛海尔生物医疗股份有限公司);1510多功能酶标仪(赛默飞世尔科技有限公司);SQ510C立式压力蒸汽灭菌锅(重庆雅马拓科技有限公司);BGZ-240电热鼓风干燥箱(上海博迅实业有限公司医疗设备厂);TY-80B水平摇床(北京佳航博创科技有限公司)。
2 方法
2.1 实验分组
2.1.1 甘草次酸给药浓度的确定 本部分按照文献[14]将实验用细胞分为共7组,分别为空白组(接种细胞,给与培养基、MTT、DMSO),甘草次酸20 μmol/L组、30 μmol/L组、60 μmol/L组、90 μmol/L组、120 μmol/L组、调零孔组(不接种细胞,给与培养基、MTT、DMSO)。
2.1.2 乌头碱配伍甘草次酸前后心肌细胞存活率实验 本部分实验用细胞共分为11组,分别为空白组(接种细胞,给与培养基、MTT、DMSO);乌头碱单用组参考文献[15]及预实验将按等比共设为4组,即120、240、480、960 μmol/L乌头组,分别简称乌头碱单用一组、乌头碱单用二组、乌头碱单用三组、乌头碱单用四组;60 μmol/L甘草次酸组;乌头碱配伍60 μmol/L甘草次酸组共4组,即120 μmol/L乌头碱配伍60 μmol/L甘草次酸组、240μmol/L乌头碱配伍60 μmol/L甘草次酸组、480 μmol/L乌头碱配伍60 μmol/L甘草次酸组、960 μmol/L乌头碱配伍60 μmol/L甘草次酸组,分别简称配伍甘草次酸一组、配伍甘草次酸二组、配伍甘草次酸三组、配伍甘草次酸四组;调零孔组(不接种细胞,给与培养基、MTT、DMSO)。
2.1.3 心肌细胞能量代谢相关指标检测实验 本实验用细胞共分为10组,不设调零孔组,其余各组参见“2.1.2”项下。
2.2 细胞存活率计算
细胞存活率参照文献[16]中公式计算:细胞活力=[A给药组-A调零孔组]/[A空白组-A调零孔组]×100%,其中A给药组:经给药处理并以MTT、DMSO处理后的孔的吸光度,A调零孔组:无细胞,其余步骤相同处理的孔的吸光度,A空白组:未经给药并以MTT、DMSO处理后的孔的吸光度。
2.3 MTT法测定甘草次酸的给药浓度
取对数生长期的H9c2细胞接种于96孔板中,每孔1×104个细胞,培养24 h后更换无血清DMEM继续培养12 h,然后弃去DMEM,按照“2.1.1”分组,分组后给予对应药物,空白组、调零孔组给予空白DMEM,在37 ℃、5% CO2培养箱中培养24 h,吸出药液,各孔加入100 μL 0.5 μmol/L的MTT,培养箱内孵育4 h,弃去MTT,加入100 μL DMSO,置于摇床低速震荡10~15 min后使用酶标仪在490 nm处测定吸光度。
2.4 MTT法测定乌头碱配伍甘草次酸前后心肌细胞存活率
取對数生长期的H9c2细胞接种于96孔板中,每孔1×104个细胞,培养24 h后更换无血清DMEM继续培养12 h,弃去DMEM,按照“2.1.2”分组,分组后给予对应药物,空白组、调零孔组给予空白DMEM,在5% CO2、37 ℃培养箱中培养24 h,其余步骤参照“2.3”项。
2.5 心肌细胞能量代谢指标检测
2.5.1 ELISA法检测心肌细胞内ATP含量 取对数生长期的H9c2细胞以1×105个/mL的浓度接种于6孔板,培养24 h后更换空白DMEM继续培养12 h,吸去培养基,按照“2.1.3”分组,分组后给予对应药物,其中空白组给予空白DMEM,培养24 h,吸去含药培养基,用PBS清洗细胞1次,然后每孔加入300 μL裂解液,裂解细胞。按照ATP含量试剂盒操作步骤,用多功能酶标仪测定RLU值(相对光单位),根据标准曲线计算样本中ATP浓度。
2.5.2 DCFH-DA染色法测定心肌细胞内ROS浓度 按照“2.5.1”中的相关步骤培养H9c2细胞,药物处理后,按照ROS测定试剂盒的使用说明书进行后续操作,最后按照异硫氰酸荧光素荧光检测条件,使用荧光倒置显微镜下观察、拍照。检测完成后使用Image J软件对其进行分析。
2.5.3 线粒体膜电位探针(5,5′6,6′-tetrachloro-1,1′,3,3′-tetraethyl-imidacarbocyanine iodide, JC-1)染色法测定心肌细胞线粒体膜电位。按照“2.5.1”项中的相关步骤培养H9c2细胞,药物处理后,采用JC-1染色法检测线粒体膜电位,具体步骤按试剂盒说明书进行操作,使用荧光显微镜观察、拍照。检测完成后使用Image J软件对其进行分析。
2.5.4 Rhod-2 AM探针法检测心肌细胞线粒体Ca2+浓度 按照“2.5.1”项下步骤培养细胞,药物处理后,按照以下步骤操作:按1∶1 000用无酚红无血清培养基稀释Rhod-2AM荧光探针,使其终浓度为5 μmol/L,去除含药培养基,以HBSS清洗细胞1次,每孔加入1 mL 5 μmol/L的 Rhod-2 AM荧光探针,放入37 ℃细胞培养箱内孵育30 min后,用HBSS平衡盐溶液洗涤细胞2次,加入少量空白无酚红培养基,荧光倒置显微镜下按照四甲基罗丹明-5(6)-异硫氰酸荧光条件观察、拍照。检测完成后使用Image J软件对其进行分析。
2.5.5 ELISA法检测Ca2+-ATP酶、Ca2+-Mg2+-ATP酶、Na+-K+-ATP酶活性 按照“2.5.1”项步骤培养细胞,药物处理后,首先去除含药培养基,以PBS清洗细胞2次后,每孔加入0.5 mL 0.25%胰酶消化50 s,每孔加入1 mL完全培养基终止消化。收集细胞至2 mL离心管,350 g离心5 min,弃去上清后加入300 μL 生理盐水,涡旋混匀,制备成106~107/cm3的细胞悬液,使用反复冻融法破碎细胞。反复冻融法步骤如下:将细胞悬液在-20 ℃冰箱内冷冻30 min以上,然后在37 ℃水浴中解冻,重复3次裂解细胞。再严格按照BCA试剂盒说明书测定各样本总蛋白浓度。最后严格按照Ca2+-ATP酶、Ca2+-Mg2+-ATP酶、Na+-K+-ATP酶试剂盒说明书测定各样本ATP酶活性。
2.6 统计学方法
采用SPSS 21.0统计软件对实验数据进行处理,先对数据进行正态性检验及方差齐性检验,若服从正态分布且满足方差齐性,则进行单因素方差分析,各组数据之间采用LSD多重比较,若服从正态分布而不满足方差齐性,则进行单因素方差分析,各组数据之间采用Games-Howell多重比较,若不服从正态分布,则进行非参数检验,计量数据以“x±s”表示。均以P<0.05为差异有统计学意义。
3 结果
3.1 甘草次酸给药浓度确定
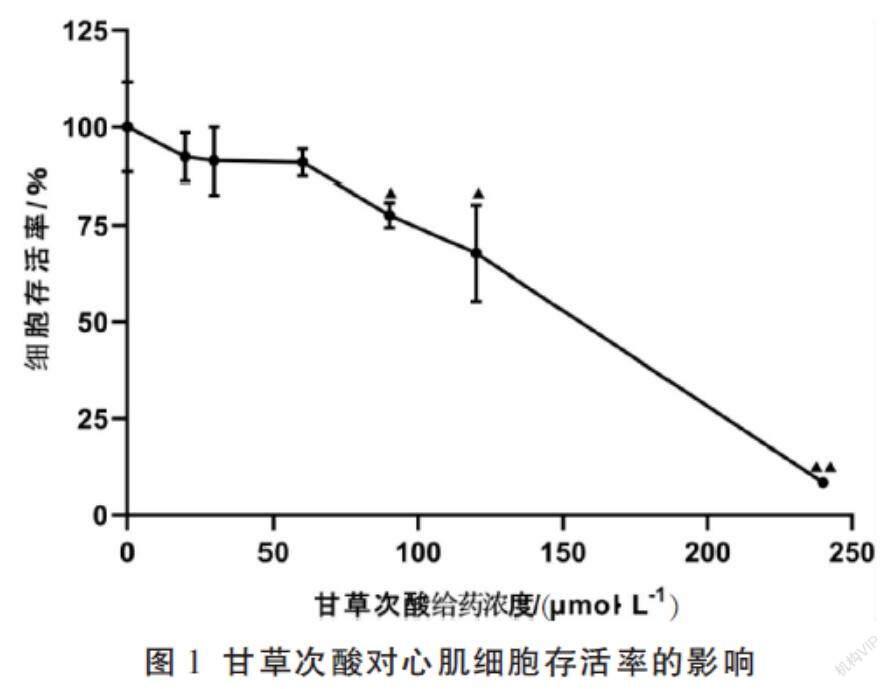
甘草次酸选择心肌细胞存活率与空白组无显著差异的最高安全浓度,以期各组分在安全浓度范围下最大可能地发挥其减毒作用,因此,确定甘草次酸的给药浓度为60 μmol/L。见图1。
3.2 乌头碱与甘草次酸配伍后心肌细胞存活率提高
与空白组比较,乌头碱会降低心肌细胞存活率(P<0.01),且该作用与剂量呈正比关系;与单用乌头碱组相比,乌头碱与甘草次酸配伍后,心肌细胞存活率均提高(P<0.01)。见表1。
3.3 乌头碱与甘草次酸配伍后心肌细胞内ATP含量增加
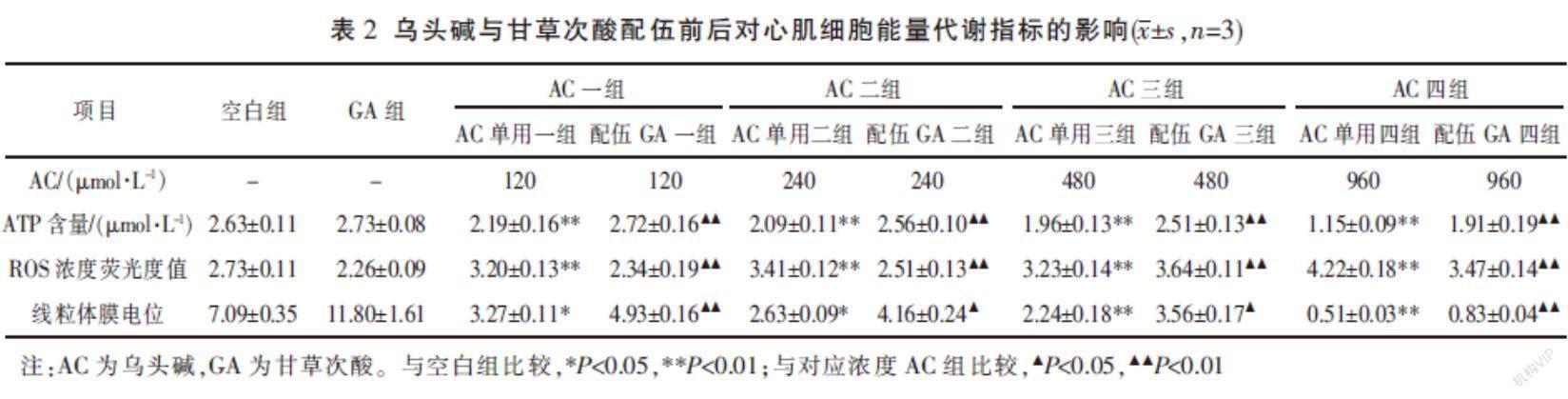
与空白组比较,乌头碱降低心肌细胞ATP含量(P<0.01),且该作用与剂量呈正比关系;与单用乌头碱组相比,乌头碱与甘草次酸配伍后,心肌细胞内ATP含量均升高(P<0.01)。见表2。
3.4 乌头碱与甘草次酸配伍后心肌细胞内ROS含量降低
与空白组比较,乌头碱提高心肌细胞内ROS含量(P<0.01),且该作用与剂量呈现正比关系;与单用乌头碱组相比,乌头碱与甘草次酸配伍后,心肌细胞内ROS含量均降低(P<0.01)。见表2、图2。
3.5 乌头碱与甘草次酸配伍后心肌细胞线粒体膜电位提高
与空白组比较,乌头碱降低心肌细胞线粒体膜电位(P<0.01或P<0.05),且该作用与剂量呈现正比关系;与单用乌头碱组相比,乌头碱与甘草次酸配伍后,心肌细胞线粒体膜电位提高(P<0.01或P<0.05)。见表2、图3。
3.6 乌头碱与甘草次酸配伍后心肌细胞线粒体内 Ca2+浓度降低
与空白组比较,乌头碱使心肌细胞线粒体内钙离子浓度增加(P<0.01),该作用与剂量呈现正比关系;与单用乌头碱组相比,乌头碱与甘草次酸配伍后,可降低心肌细胞内线粒体内钙离子浓度(P<0.01)。见图4、表3。
3.7 乌头碱与甘草次酸配伍后心肌细胞内Ca2+-ATP酶、Ca2+-Mg2+-ATP酶、Na+-K+-ATP酶活性提高
与空白组比较,乌头碱降低心肌细胞内Ca2+-ATP酶活性(P<0.01)、Ca2+-Mg2+-ATP酶活性(P<0.01或P<0.05)、Na+-K+-ATP酶活性(P<0.01),该作用与剂量呈现正比关系。与单用乌头碱组相比,乌头碱与甘草次酸配伍后,可提高心肌细胞内Ca2+-ATP酶活性(P<0.01或P<0.05)、Ca2+-Mg2+-ATP酶活性(P<0.01或P<0.05)、Na+-K+-ATP 酶活性(P<0.01或P<0.05)。见表3。
4 讨论
“附子-甘草”是相畏相杀的经典药对,也是中药配伍减毒研究的热点。组分配伍研究是一种阐明中药配伍关系的新模式,遵循“复杂-简单-复杂”研究原则,具有作用靶点明确、作用机制清楚等优点[17]。故本文将乌头碱与甘草次酸组分配伍作为研究对象,以探讨“附子-甘草”药对配伍减毒机制。H9c2细胞在心肌细胞信号传导通路、细胞周期凋亡调控相关基因表达和细胞增殖及心肌细胞损伤保护机制方面的研究中应用广泛[18-21]。本研究结果显示,单用乌头碱降低心肌细胞存活率,乌头碱与甘草次酸配伍后心肌细胞存活率显著提高,证实甘草次酸可拮抗乌头碱所致的心肌细胞毒性作用。
本研究发现,单用乌头碱时,心肌细胞ATP含量、膜电位水平降低,ROS含量则明显上升;乌头碱与甘草次酸配伍后,心肌细胞ATP含量、膜电位水平明显上升,ROS含量则明显降低。乌头碱的心肌毒性主要是损伤心肌细胞线粒体[22],线粒体的损伤途径主要包括:(1)破坏线粒体内膜完整性,使线粒体ATP减少;(2)产生大量ROS,对线粒体造成氧化损伤;(3)诱导线粒体通透性转换孔过度开放,引起线粒体膜电位下降[23]。ATP含量可以直观反映心肌细胞内能量代谢的水平,而线粒体是心肌细胞中负责能量代谢的重要细胞器,线粒体正常膜电位是维持线粒体能量代谢的先决条件。一方面,线粒体在呼吸氧化过程中,将所产生的能量以电化学势能储存于线粒体内膜,在内膜两侧造成质子以及其他离子浓度的不对称分布而形成膜电位。另一方面,线粒体在产能的同时会产生大量的ROS,而当ROS过高时,线粒体通透性转换孔的开放又会导致线粒体通透性的增加。故,此部分研究结果提示,甘草次酸缓解乌头碱对心肌细胞的损伤与其改善心肌细胞线粒体功能有关。
线粒体还是维持心肌細胞内钙稳态的重要缓冲器,而心肌细胞内Ca2+的转运异常则会引起心脏结构变化和功能的紊乱[24],最终容易诱发心律失常、心力衰竭等症。正常情况下,心肌细胞内相关ATP酶对维持心肌细胞钙稳态以及线粒体钙稳态都具有重要作用,此类酶的活性也能反映细胞内能量代谢的水平。其中,Ca2+-ATP酶主要是将细胞内游离Ca2+转出细胞或摄入肌浆网,以预防细胞内钙超载[25];Ca2+-Mg2+-ATP酶促进肌浆网内Ca2+的释放,并转运至胞外,维持胞内钙稳态[26];Na+-K+-ATP酶通过泵出细胞内的Na+和泵入胞外的K+以维持细胞内外离子浓度稳定和动态平衡,从而维持心肌细胞的正常节律; Na+/Ca2+交换间接可调控细胞内的Ca2+活动,对维持线粒体膜内外离子平衡、渗透压平衡发挥重要作用[27]。本实验研究发现,单用乌头碱时,心肌细胞线粒体内Ca2+浓度提高,心肌细胞内Ca2+-ATP酶、Ca2+-Mg2+-ATP酶、Na+-K+-ATP酶的活性出现显著降低,表明乌头碱有心肌毒性;与甘草次酸配伍后,线粒体内Ca2+浓度降低,3种ATP酶活性提高。由此可见,甘草次酸拮抗乌头碱的心肌毒性与增加ATP酶活性,从而促进细胞内Ca2+转入肌浆网或细胞外,进而缓解线粒体钙超载,维持钙稳态有关。
综上所述,本实验研究结果证实,甘草次酸可拮抗乌头碱的心肌毒性,改善心肌细胞线粒体能量代谢,其解毒作用环节涉及增加ATP生成、抑制ROS释放、平衡膜电位等,具体机制可能与缓解线粒体钙超载有关。
参考文献
[1] 彭 伟,王 琳,傅超美,等.基于网络药理学的附子抗心力衰竭作用和心脏毒性的毒效二重性研究[J].中医杂志,2021,62(6):523-529.
[2] 宋亚刚,崔琳琳,李 艳,等.中药“减毒药对”研究方法探讨及思考[J].中华中医药学刊,2020,38(10):76-80.
[3] 刘曾晶,张梦林,康前前,等.基于数据挖掘的抗肿瘤中药方剂用药规律分析[J].亚太传统医药,2020,16(8):143-146.
[4] 彭 兰.附子配甘草对离体心脏的作用及其指纹图谱研究[D].长沙:湖南中医药大学,2019.
[5] 张序晴,彭 兰,李 赛,等.附子-甘草对离体蛙心的影响及其指纹图谱研究[J].湖南中医药大学学报,2019,39(2):184-189.
[6] 彭 兰,张序晴,王志琪,等.附子-甘草对离体大鼠衰竭心脏的影响及其指纹图谱研究[J].中南药学,2019,17(3):379-383.
[7] 王志琪,曾 嵘,谭志荣,等.附子与甘草配伍前后乌头碱和甘草次酸在大鼠体内的药动学比较[J].中成药,2012,34(12):2305-2309.
[8] 彭 成.中药附子毒效多维评价与整合分析的思路与实践[J].世界中医药,2017,12(11):2543-2550.
[9] 谢晓芳,彭 成.附子心脏毒效的多维评价和整合分析研究进展[J].世界中医药,2017,12(11):2555-2562.
[10] 杨继媛,吴红金.甘草次酸抗实验性心律失常及机制的研究[J].中西医结合心脑血管病杂志,2010,8(10):1229-1231.
[11] 高 艳,谢灵燕,卢志强.附子毒理及减毒增效配伍的研究进展[J].天津药学,2020,32(1):65-69.
[12] 赵佳伟,何家乐,马增春,等.附子对H9c2心肌细胞系线粒体的毒性作用机制[J].中国药理学与毒理学杂志,2015,29(5):816-824.
[13] 肖 晨,陈思君,危当恒,等.线粒体功能的检测方法[J].生命的化学,2020,40(2):222-229.
[14] 王利勤,张宇燕,何 昱,等.附子、甘草有效成分不同配伍比例对H9c2心肌细胞缺氧缺糖损伤的影响[J].中医杂志,2016,57(15):1327-1331.
[15] 周天梅.附子甘草活性物质配伍对乌头碱致传代心肌细胞损伤保护作用的实验研究[D].杭州:浙江中医药大学,2013.
[16] 刘长河,李开言,张雪侠,等.木豆叶、鲜地黄提取物对ox-LDL损伤的HUVEC的保护作用[J].中药新药与临床药理,2021,32(8):1073-1078.
[17] 李 文,傅超美,傅 舒.附子配伍甘草对大鼠心肌细胞毒性机制的研究进展[J].中药与临床,2018,9(6):60-64.
[18] 覃斐章,董 敏,秦秋华,等.玉郎伞查尔酮激活Nrf2/ARE信号通路减轻缺氧/复氧所致的H9c2细胞凋亡及氧化应激损伤[J].天然产物研究与开发,2020,32(6):1038-1044.
[19] 李 津,李 羽,刘明洁.蓝萼甲素对H2O2诱导H9c2心肌细胞凋亡的保护作用及机制研究[J].国际中医中药杂志,2020,42(12):1137-1144.
[20] 杨 萍,代 天,张苏川.当归多糖促进大鼠心肌细胞系H9c2增殖[J].基础医学与临床,2021,41(4):551-557.
[21] 杨 洁,郑瑞芳,都研文,等.田蓟苷抗H9c2心肌细胞缺血再灌注损伤的作用机制研究[J].药学学报,2021,56(4):1070-1078.
[22] 王宁宁.乌头碱激活Sirt3改善心肌细胞线粒体功能[D].广州:广东药科大学,2019.
[23] 雷 蕾,彭双清.药物损伤心肌细胞线粒体致心脏毒性的分析[J].河南大学学报(医学版),2012,31(2):138-141.
[24] TAKAHASHI M, YOKOSHIKI H, MITSUYAMA H, et al. SK channel blockade prevents hypoxia-induced ventricular arrhythmias through inhibition of Ca2+/voltage uncoupling in hypertrophiedhearts[J]. American Journal of Physiology Heart and Circulatory Physiology, 2021, 320(4): H1456-H1469.
[25] SKOGESTAD J, ARONSEN J M, TOVSRUD N, et al. Coupling of the Na+/K+-ATPase to Ankyrin B controls Na+/Ca2+ exchanger activity in cardiomyocytes[J]. Cardiovascular Research, 2020, 116(1): 78-90.
[26] DESHPANDE L S, DELORENZO R J, CHURN S B, et al. Neuronal-specific inhibition of endoplasmic Reticulum Mg2+/Ca2+ ATPase Ca2+ uptake in a mixed primary hippocampal culture model of status epilepticus[J]. Brain Sciences, 2020, 10(7): 438.
[27] SILVA C I D, GON?覶ALVES-DE-ALBUQUERQUE C F, DE MORAES B P T, et al. Na/K-ATPase: Their role in cell adhesion and migration in cancer[J]. Biochimie, 2021, 185: 1-8.
(本文編辑 苏 维)
