
库尔勒香梨短果枝花序维管束与萼片脱落关系研究
2021-11-22 03:27:42金敏张倩张天正包建平吴翠云张锐陶书田
中国农业科技导报 2021年11期
金敏, 张倩, 张天正, 包建平,2,3*, 吴翠云,2,3, 张锐,2,3, 陶书田
(1.塔里木大学植物科学学院,新疆 阿拉尔 843300;2.塔里木大学,新疆生产建设兵团塔里木盆地生物资源保护利用重点实验室,新疆 阿拉尔 843300;3.塔里木大学,南疆特色果树高效优质栽培与深加工技术国家地方联合工程实验室,新疆 阿拉尔 843300;4.南京农业大学园艺学院,南京 210095)
库尔勒香梨(PyrusbrestschneideriRehd.,以下简称“香梨”)属于蔷薇科、梨属,是新疆梨和西洋梨的自然杂交后代,是新疆主栽梨品种,以皮薄、肉脆、汁多、味甜、酥香、爽口、耐储藏、营养丰富等特点驰名中外[1]。根据萼片脱落与否,香梨果实分为脱萼果和宿萼果,而宿萼和脱萼果实严重影响库尔勒香梨的外观品质,降低经济效益,挫伤梨农种植积极性[2-3]。因此,深入研究萼片脱落或宿存的影响机制,对提高香梨果实的脱萼率,改善果实的综合品质,以及香梨优质栽培具有重要的现实意义。
维管束是一个相互连通的重要输导系统,连接于植物的器官之间,为信号分子的传递、水及营养物质的输送提供了通路。梨果实中呈网状分布的维管束是一个相互连通的重要输导系统,对果实的生长发育和品质形成有重要作用,是果实水分和营养物质运输的主要通道。目前研究发现,植物特定组织或器官内的维管组织分化与其生长素浓度有关,一般生长素浓度较高的植物体组织或器官,其维管系统也相应较为发达[4-5]。生长素在植物维管组织分化及维管束形成过程中起重要作用。有研究指出,外源生长素的施用部位可以决定维管组织的分化区域,在该区域产生向下延伸的新维管束,且这些新维管束是以外源生长素的施用部位为起点的[6]。王丰等[7]的研究表明,用生长素抑制剂处理叶片可发现周围部分组织的生长素浓度升高,维管束发达;其他组织生长素减少,维管束生长受抑制。用生长素转运抑制剂NPA抑制生长素流能够抑制导管元的分化[8]。果实中的内源激素主要来自叶片、枝条的供给,而果实中内源激素运输的通道为维管束,其数目和面积直接影响激素的运输和积累,进而影响萼片的脱落与宿存。目前关于脱萼宿萼机理,特别是库尔勒香梨萼片脱落与宿存在显微结构方面的差异变化的研究还很少。本研究通过对大蕾期库尔勒香梨树体喷施吲哚乙酸(indoleacetic acid,IAA)和三碘苯甲酸(triiodobenoic acid,TIBA)处理,分期取样观察其显微结构变化,旨在通过库尔勒香梨宿萼果和脱萼果萼片组织结构的变化研究,分析产生脱萼果和宿萼果的原因,为进一步探明脱萼机理、改善库尔勒香梨果实品质奠定理论基础。
1 材料与方法
1.1 试验地点及材料
试验地点位于新疆阿拉市塔里木大学校园。供试品种为以杜梨作砧木的香梨,南北行向,灌溉方式为大水漫灌。土壤类型为砂壤土。果园常规管理,树龄23 a,株行距2 m×4 m。
1.2 材料采集
选择树体生长状况相对一致的30株香梨作为采样株。2019年4月2日(大蕾期)进行处理,喷施100 mg·L-1IAA(上海蓝季生物有限公司),100 mg·L-1TIBA(源叶生物有限公司),以喷施清水为对照(CK),每个处理各10株,树冠西侧外围短果枝上的花芽编号挂牌。4月2日喷施前采第1次样,以后每隔2 d采样一次,采至幼果坐稳期(4月17日),共计6次,采样覆盖整个香梨萼片脱落时期。依次取下脱萼率较高的第4序位花,迅速将样品带回实验室,分别将花萼、胴部和果柄3个部位放入 FAA固定液固定,备用。
1.3 试验方法
1.3.1石蜡切片的制作 将样本参照文献[9-10]的方法制作石蜡切片。先将样本从FAA固定液中取出使用系列浓度的乙醇溶液与透明剂完成样本的脱水和透明;然后使用石蜡进行包埋,切片机切片,烘箱烘片;接着使用系列浓度的乙醇溶液与透明剂复水,使用1%番红染液和0.5%固绿染液双重染色;最后得到韧皮部为绿色,木质部为红色的石蜡切片;中性树胶封片,使用奥林帕斯显徽镜(BX50F-3)进行观察拍照同时测量其萼片主维管束(横切形状可近似为圆形)面积、数目。
1.3.2脱萼率调查 花后20 d,调查喷施清水(CK)、喷施IAA处理和TIBA处理下香梨短果枝第4序位的萼筒脱萼率,计算公式如下。
萼筒脱萼率=萼筒脱萼果/总果数×100%
(1)
1.4 数据处理
采用Microsoft Excel 2010进行数据处理及作图,采用DPS 7.05软件进行单因素对数据进行方差分析。
2 结果与分析
2.1 不同处理下香梨的脱萼率
从图1可以看出,清水处理(对照)、IAA处理和TIBA处理下香梨的脱萼率分别为43.4%、38.4%和78.1%。 TIBA处理下香梨脱萼率显著高于其他2个处理,对照显著高于IAA处理的香梨萼片脱萼率。说明喷施IAA会促进萼片的宿存,影响萼片的脱落;而生长素抑制剂TIBA则会促进萼片的脱落,提高脱萼率。
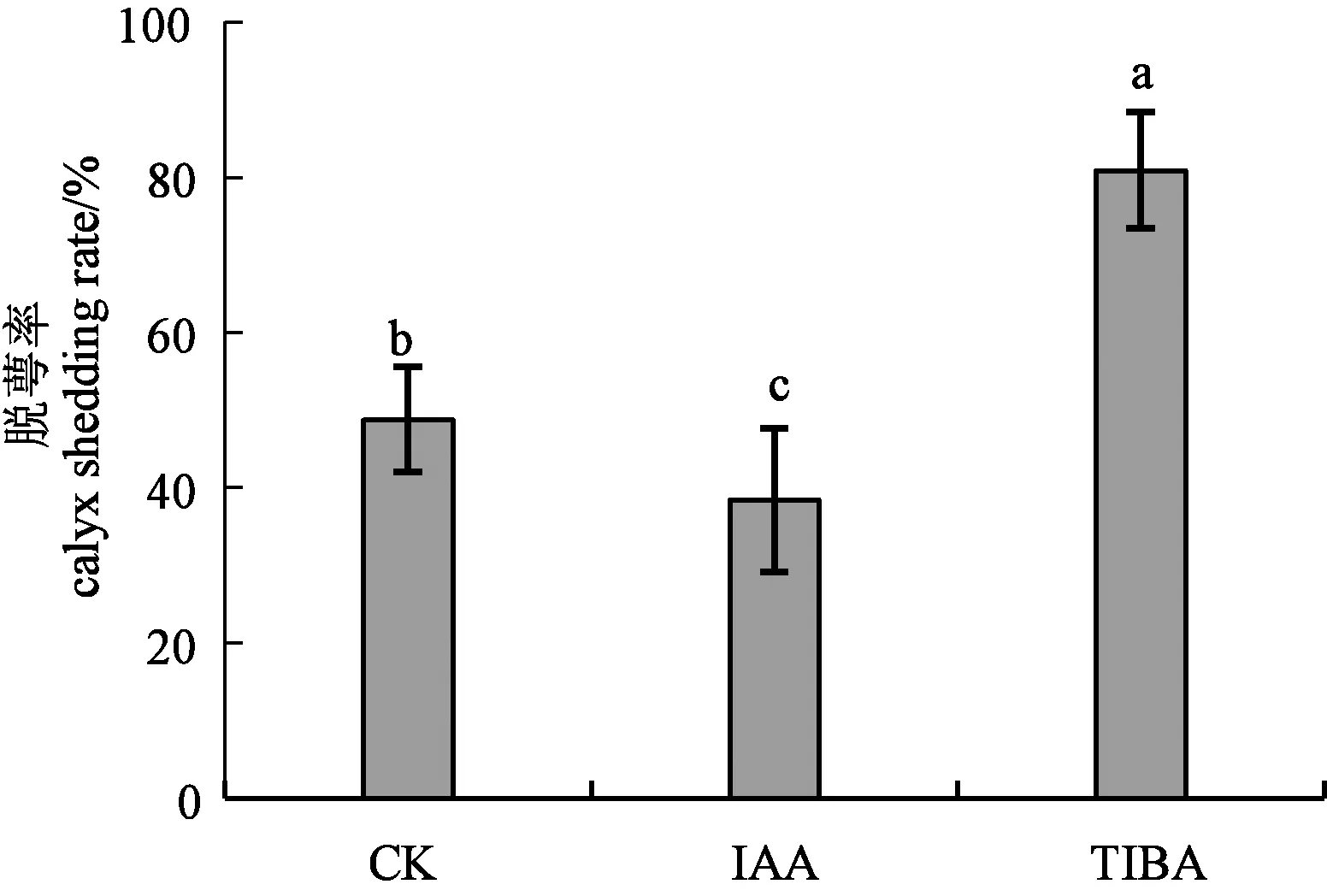
注:不同小写字母表示差异显著(P<0.05)。
2.2 不同处理下香梨果实维管束数目
2.2.1不同处理下香梨果实萼筒维管束数目 从图2可以看出,对照与IAA、TIBA处理的香梨萼筒维管束数目变化趋势相似,均呈上升-下降-上升-下降的趋势。对照和IAA处理在4月8日出现最大值,均为11.67个,TIBA处理在4月14日达到最大,为9.83个。IAA和TIBA处理在4月11日出现最小值,分别为10.33和8.33个,对照组在4月2日是最低值,为9.0个。3个处理均在4月2日至8日期间呈逐渐上升的趋势,于4月11日发生下降。由此可见,4月8日是香梨萼筒维管束数目发育的临界时期。4月2日,萼筒维管束数目IAA处理显著高于对照,与TIBA处理间差异不显著。4月5日,对照与TIBA处理的萼筒维管束数目不存在显著性差异,两者均显著低于IAA处理的萼筒维管束数目,4月8日、11日和17日,萼筒维管束数目对照显著高于TIBA处理,与IAA处理间差异不显著。4月14日时TIBA处理萼筒维管束数目显著低于对照和IAA处理。
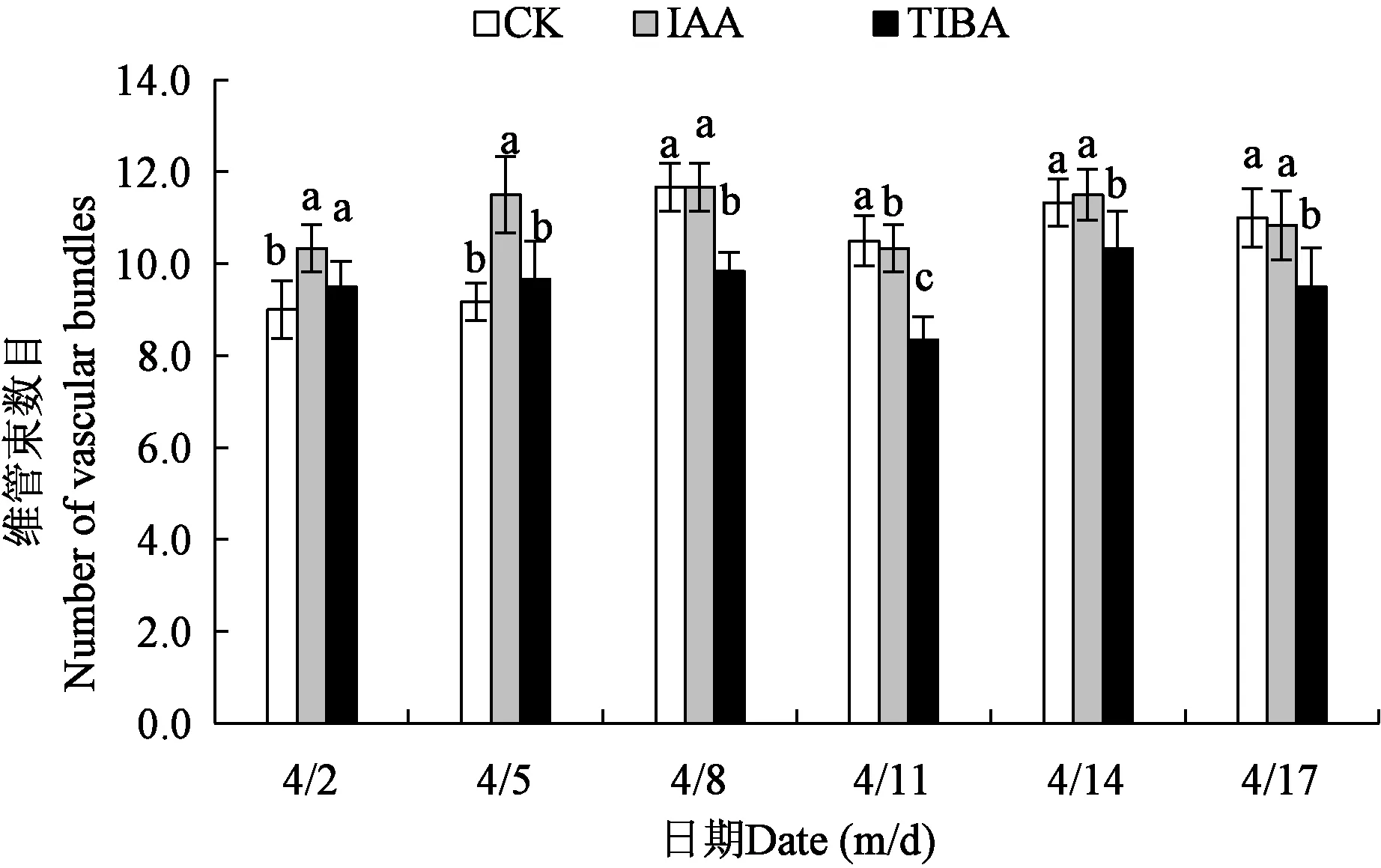
注:同一日期下不同小写字母表示差异显著(P<0.05)。
2.2.2不同处理下香梨果实胴部维管束数目 从图3可以看出,对照和IAA处理在4月14日出现最大值,均为11.67个;TIBA处理在4月8日达到最大,为10.0个。IAA和TIBA处理在4月11日出现最小值,分别为10.33和9.50个;对照在4月2日出现最低值,为9.0个。对照和TIBA处理下均在4月2—8日期间呈逐渐上升的趋势,4月11日发生下降,因此,认为4月8日是香梨胴部维管束数目发育的临界时期。在4月2—5日,对照和TIBA处理的胴部维管束数目无显著差异,显著低于IAA处理。4月8日,IAA处理和TIBA处理的胴部维管束数目无显著差异,显著低于对照。4月11—17日,对照与IAA处理的胴部维管束数目无显著差异,均显著高于TIBA处理。对照组与IAA、TIBA处理的香梨胴部维管束数目变化趋势大致相似,呈上升-下降-上升-下降的趋势。

注:同一日期下不同小写字母表示差异显著(P<0.05)。
2.2.3不同处理下香梨果实果柄维管束数目 从图4可以看出,3个处理在调查期间的变化趋势大致相似,呈升高-降低-升高的变化,但出现最大值或最小值的时期不同,对照组在4月5日出现最大值,为10.33个;而IAA处理和TIBA处理在4月14日达到最大,为10.50个。3个处理均在4月11日出现最小值,分别为10.17、9.67和9.00个。对照在4月2—8日期间呈逐渐上升的趋势,于4月11日发生下降并达到最低值,因此,认为4月8日是香梨果柄维管束数目发育的临界时期。4月2日、5日、14日3个处理间的香梨果柄维管束数目差异不显著。4月8日、11日,果柄维管束数目TIBA处理显著低于对照组。与IAA处理间差异不显著。 4月17日时,TIBA处理果柄维管束数目显著低于IAA处理,与对照组间不存在显著性差异。

注:同一日期下不同小写字母表示差异显著(P<0.05)。
2.3 不同处理下香梨果实维管束面积
2.3.1不同处理下香梨果实萼筒维管束面积 从图5可以看出,3个处理在调查期间的变化趋势大致相似,呈上升-下降-上升的趋势,但下降变化时间不一致,对照在4月2—11日上升,随后发生下降,4月14—17日又升高。IAA处理和TIBA处理的萼筒维管束面积,4月2—8日上升,随后发生下降,4月11—17日又升高。4月2—17日,对照的萼筒维管束面积均与IAA处理不存在显著差异,与TIBA在4月8日、14日和17日,不存在显著差异外,其他时期均显著高于TIBA处理。除4月14日,TIBA处理的萼筒维管束面积与IAA处理不存在显著差异外,其余时期均显著低于IAA处理。

注:同一日期下不同小写字母表示差异显著(P<0.05)。
2.3.2不同处理下香梨果实胴部维管束面积 从图6可以看出,不同处理的胴部维管束面积变化趋势大致相似,呈上升-下降-上升的趋势,4月2—8日逐渐上升,于4月11日骤然下降,随后又升高,由此可见,4月8日是香梨胴部维管束面积发育的界时期。4月2日,对照的胴部维管束面积显著高于IAA处理和TIBA处理,TIBA处理有显著高于IAA处理的胴部维管束面积。4月5日,不同处理的维管束面积无显著性差异。4月8日,对照与IAA处理的胴部维管束面积无显著差异,但两者显著低于TIBA处理。4月14日,对照的胴部维管束面积显著高于IAA处理和TIBA处理,IAA处理有显著高于TIBA处理的胴部维管束面积。4月17日,对照的胴部维管束面积显著高于TIBA处理,与IAA处理无显著差异,IAA处理的胴部维管束与TIBA处理的也无显著差异。
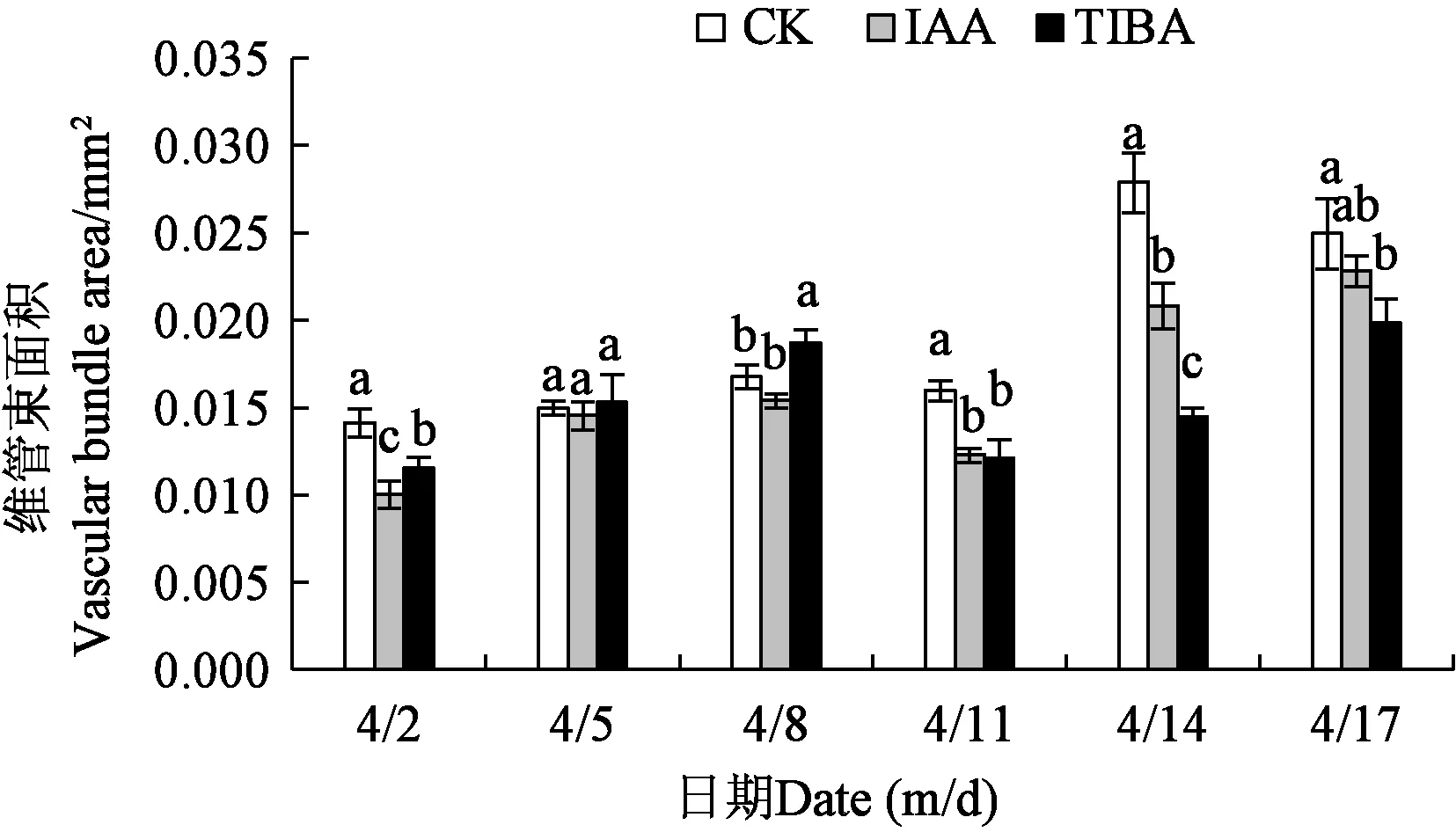
注:同一日期下不同小写字母表示差异显著(P<0.05)。
2.3.3不同处理下香梨果实果柄维管束面积 从图7可以看出,不同处理的果柄维管束面积的变化趋势大致相似,呈上升-下降-上升的趋势,但IAA处理的后期变化时间与其他两个处理有所区别,对照与TIBA处理在4月2—8日逐渐上升,于4月11日下降,4月14日又升高;而IAA处理在前期与两者一致,但从4月11日下降后在4月14日持续降低,4月17日再次升高。4月2日,对照与IAA处理的果柄维管束面积无显著差异,均显著高于TIBA处理。4月5日,不同处理间的维管束面积无显著性差异。4月8日、11日和17日,对照显著高于IAA处理和TIBA处理的果柄维管束面积,IAA处理与TIBA处理无显著差异。4月14日,对照的果柄维管束面积显著高于IAA处理,与TIBA处理无显著差异,IAA处理的果柄维管束与TIBA处理的也无显著差异。
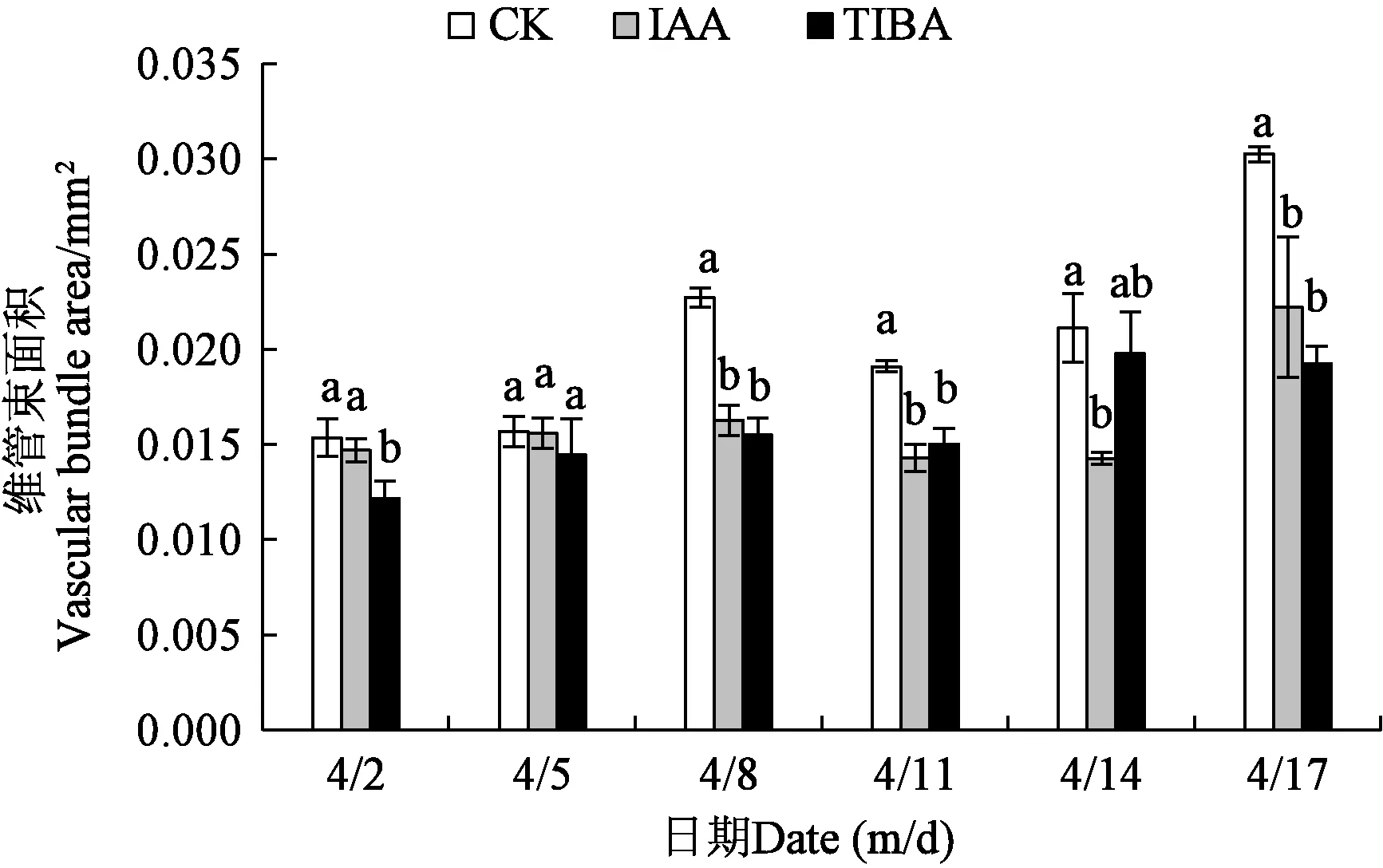
注:同一日期下不同小写字母表示差异显著(P<0.05)。
2.4 脱萼率、维管束数目和维管束面积的相关性
由表1可知,脱萼率与胴部维管束数目、萼筒维管束数目均呈极显著性负相关,相关系数分别为-0.67、-0.61;与果柄维管束数目和萼筒维管束面积均呈显著性负相关,相关系数分别为-0.52、-0.46;与胴部维管束面积、果柄维管束面积相关性不显著。果柄维管束数目与胴部维管束数目、萼筒维管束数目、果柄维管束面积和萼筒维管束面积呈显著性正相关,相关系数分别为 0.50、0.52、0.51、0.54。胴部维管束数目与萼筒维管束数目呈极显著性正相关,相关系数为0.79;与果柄维管束面积、胴部维管束面积、萼筒维管束面积呈显著性正相关,相关系数为0.47、0.54、0.50。萼筒维管束数目与萼筒维管束面积呈显著性正相关,相关系数为0.56;果柄维管束面积与胴部维管束面积、萼筒维管束面积呈极显著性正相关,相关系数分别为0.69、0.74。胴部维管束面积与萼筒维管束面积呈极显著正相关,相关系数为0.59。

表1 脱萼率、维管束数目和维管束面积的相关性分析
3 讨论
香梨花萼包括萼筒和萼片两部分。萼筒上着生花瓣和雄蕊,开花期萼筒呈盘状。萼筒前端裂片为萼片。脱萼果是从萼筒下部连同萼片一起脱落,萼凹圆而光;宿萼果是萼筒随幼果发育成果顶的一部分,呈凸出状[11]。宿萼果石细胞含量高,肉质硬,果实品质差,目前生产上采用一些有效的方法促使花萼脱落,如选择不同砧木类型、人工授粉、喷施生长调节剂或剪萼等[12],以减少宿萼果比例,改善果实品质。前人研究表明,幼果中低浓度的IAA可促进萼片脱落,高浓度IAA则利于宿萼果的产生[13-15]。高疆生等[16]和陈晓艺等[17]研究认为,PP333和烯唑醇均能提高库尔勒香梨的脱萼果率。Rosales等[18]用生长素极性运输抑制剂TIBA(三碘苯甲酸)处理西葫芦花朵离区,增加了西葫芦花的脱落率。这些研究都与本研究结果一致。本研究表明,不同处理的脱萼率大小顺序为TIBA处理>对照>IAA处理,经TIBA处理的库尔勒香梨幼果萼筒在发育后期出现了离层,萼筒在离层处断裂、脱落,形成了脱萼果。而IAA处理的香梨幼果萼筒在发育后期较少出现离层,大多数香梨萼筒没有脱落而是发育成幼果果顶的一部分,形成了宿萼果,降低了脱萼果率。
生长素在维管组织发育过程起着非常重要的作用。经生长素调节剂和生长素抑制剂处理的库尔勒香梨幼果在发育过程中显微结构具有很大差异。本研究发现,4月8日为香梨萼筒、胴部和果柄的维管束发育的临界期,数目与面积均在4月11日降低,而据调查,在4月11日香梨果萼脱萼处于高峰时期。3个处理的香梨维管束数目变化趋势大致相似,呈现升高-降低-升高-降低的变化趋势,均在8~12个不等,对照和IAA处理的果柄、胴部和萼筒部位的维管束数目均大于TIBA处理,而对照与IAA处理的萼筒维管束数目差异不显著。这与马宏超等[19]的研究结果不一致,其研究认为,在整个花期中,经赤霉素和多效唑处理的萼筒维管束数目一直保持不变,均为 8个。库尔勒香梨经多效唑处理的萼筒在发育过程中维管束中只有导管,未见有筛管和异细胞的形成,花萼中水分和养料的供给缺乏,导致萼筒在发育后期出现了离层,萼筒脱落,果实发育形成脱萼果。过量的表达生长素同样会提高维管组织的分化[20]。Fukuda[21]和Alonii[22]研究也发现,对植株局部施用外源生长素后,该部位及其周围区域维管组织都比对照发达。这与本研究结果一致。本研究中,3个处理的果柄和胴部维管束面积大小顺序为:对照>IAA处理>TIBA处理,而对照萼筒的维管束面积对照与IAA处理无显著差异,但均显著高于TIBA处理。与马宏超等[19]的研究中经赤霉素处理的花萼萼筒组织结构中维管束的面积较大的结果一致。综合上述,TIBA处理会抑制萼筒维管束的数目和面积,使得花萼中水分和养料的供给缺乏,导致幼果萼筒在发育后期出现了离层,萼筒在离层处断裂、脱落,形成脱萼果,从而增加香梨的萼筒脱萼率。
猜你喜欢
新疆农业科学(2023年6期)2023-07-13 03:53:58
今日农业(2022年16期)2022-09-22 05:38:22
湖南农业科学(2022年5期)2022-06-22 08:26:06
河北农业大学学报(2021年5期)2021-11-10 13:11:28
农业与技术(2021年5期)2021-03-31 12:26:16
新疆农业科学(2020年9期)2020-10-13 12:37:42
河南农业科学(2019年9期)2019-09-24 09:10:48
山东农业工程学院学报(2019年3期)2019-05-11 07:41:50
烟台果树(2019年1期)2019-01-28 09:34:56
中国三峡(2017年4期)2017-06-06 10:44:22
