
日照潮下带海域大型底栖动物春、秋季群落结构特征及其与环境因子的关系
2021-11-19 09:01王尽文王建勇
应用海洋学学报 2021年4期
王尽文,王 燕,黄 娟,王 波*,王建勇,林 森,孙 滨
(1.山东省海洋生态环境与防灾减灾重点实验室,山东 青岛 266061;2.自然资源部第一海洋研究所,山东 青岛 266071;3.国家海洋局北海预报中心,山东 青岛 266061)
海洋大型底栖动物是指在分选大型底栖动物样品时,留在网孔为0.5 mm网筛上的动物,由于对环境污染敏感,不少种类,如小头虫(Capitellacapitata)、菲律宾蛤仔(Ruditapesphilippinarum)等可作为海洋污染的指示种。对于大型底栖动物的研究可以很好地评价海洋环境的质量状况。日照市位于山东半岛东南部,全市海域面积约为6 000 km2,沿岸有潮白河、傅疃河、绣针河等多条河流入海。沿岸河流带入的丰富营养物质加上日照近海适宜的生态环境给众多海洋生物提供了良好的栖息场所。据记载19世纪80年代日照市近岸海域有无脊椎动物140余种[1]。上世纪90年代以来,日照市对沿海开始较大规模的开发,大型的集装箱码头、发电厂、沿海观光公路等工程迅速增加。港口和临港产业的飞速发展在极大地促进当地经济发展的同时也给所在海域的生态环境造成了不可逆转的危害[2]。有研究表明日照市近岸海域的渔业资源群落结构已发生了巨大的变化[3-7]。目前针对该海域大型底栖动物群落结构及其影响因子的研究报道相对较少,其中钟海霞等(2018)和赵凡淇等(2018)在对山东南部海域的大型底栖动物群落结构研究时有涉及,但调查站位较少且仅有一个季节[8-9];齐磊磊等(2013)利用2006年的春、秋两季调查资料分析了该海域的大型底栖动物群落结构[10]。本研究利用2016年5月和11月的调查资料对日照潮下带海域大型底栖动物群落结构及其影响因子作了分析,以期为该海域海洋生态研究提供基础数据。
1 材料与方法
1.1 站位布设
自然资源部第一海洋研究所分别于2016年5月(春季)和2016年11月(秋季)在日照潮下带海域进行了大型底栖动物调查,依据水深自近岸向外布设A、B、C、D、E、F共6个断面19个站位(图1)。

图1 日照近岸海域大型底栖动物调查站位图
1.2 样品采集和分析

1.3 评价方法
1.3.1 优势度指数 大型底栖动物群落优势度指数采用以下公式[13]:
Y=ni/N×fi
(1)
式(1)中:Y为优势度指数,N为所有种的个体总数,ni为第i种的个体数目,fi为第i种的出现频率,即该种出现的站位数目占总站位数目的比例。将Y≥0.01的物种定为优势种。
1.3.2 多样性指数 利用种类多样性指数(H′)[12]、丰度指数(d)[12]和均匀度指数(J)[12]来分析群落结构:
(2)
(3)
J=H′/Hmax
(4)
式(2)至(4)中:S为样品中的种类总数,N为大型底栖动物总个体数目,Pi为第i种大型底栖动物个体数目占总大型底栖动物个体数目的比例。
1.3.3 丰度/生物量曲线 丰度/生物量曲线(ABC 曲线)为PRIMER 5.0软件自带的Dominance Plot模块绘制。若丰度曲线在生物量曲线下方则说明大型底栖动物群落未受扰动;反之,则为严重扰动;若两曲线重叠或交叉则为中度干扰[14]。
1.3.4 群落结构及环境因子分析 本研究中为减少机会种对结果的影响,在用PRIMER 5.0 软件自带的CLUSTER模块对群落进行聚类分析时,去除样品中小于1%的物种,保留其中相对丰度大于 3%的物种[15]。利用SIMPER模块对群落进行相似性分析;利用BIOENV模块对群落结构和环境因子进行分析并做RELATE关联检验。利用SPSS 25.0对丰度和生物量与环境因子做Pearson相关分析。
2 结果与讨论
2.1 环境因子
本研究所测20项水质环境因子中水温、pH、DO和盐度4项因子各站位之间差别较小,其他16项环境因子各站位之间差别较大(表1)。

表1 日照潮下带海域环境参数的季节变化
2.2 类群组成
经鉴定,两个季节共采集到大型底栖动物6门60种(表2),其中环节动物门30种,占总种类数目的35.71%;节肢动物门16种,占19.05%;软体动物门8种,占9.52%;棘皮动物门4种,占4.76%;脊索动物门(硬骨鱼类)1种,占1.19%;腔肠动物门1种,占1.19%。两个季节大型底栖动物均以环节动物门出现种类数目最多,其次为节肢动物门和软体动物门,其他3个门类均较少。两个季节出现总种类数目没有明显差别,门类分布稍有不同:春季环节动物门出现种类数目小于秋季,软体动物门出现种类数目大于秋季。

表2 日照潮下带海域大型底栖动物种名录
2.3 优势种
表3为两个季节优势度指数排名前5的物种,共10种。其中环节动物门6种,节肢动物门3种,软体动物门1种。环节动物门在两个季节均为优势门类,共6种,占60.00%;节肢动物门仅在秋季为优势门类,共3种,占30.00%;软体动物门仅在春季为优势门类,共1种,占10.00%。春季排名前5的物种优势度指数和出现频率相差较小,最大值分别为最小值的1.78倍和1.25倍;秋季排名前5的物种优势度指数和出现频率相差较大,最大值分别为最小值的36.75倍和6.33倍。春季优势种有4个,依次为巴氏钩毛虫(Sigambrabassi)、扇栉虫(Amphicteisgunneri)、小头虫(Capitellacapitata)和醒目云母蛤(Yoldianotabilis);秋季优势种有4个,依次为绒螯近方蟹(Hemigrapsuspenscillatus)、索沙蚕科(Lumbrineriiae)1种、哈氏美人虾(Callanssaharmandi)和长鳃树蛰虫(Pistabranchia);两个季节之间优势种物种更替率为100%。

表3 日照潮下带海域大型底栖动物优势度指数排名前5的物种
2.4 密度及其平面分布
在春季调查中大型底栖动物的密度变化范围在20.00~220.00 ind./m2之间,平均为96.83 ind./m2。密度组成以环节动物门种类占绝对优势,其密度为55.79 ind./m2,占总密度的57.62%;软体动物门种类密度为21.05 ind./m2,占21.74%;节肢动物门种类密度为17.89 ind./m2,占18.48%;棘皮动物门和腔肠动物门种类密度相对较低,均为1.05 ind./m2,分别占总密度的1.08%。秋季调查密度变化范围在20.00~380.00 ind./m2之间,平均为112.63 ind./m2,为春季的1.95倍。密度组成以环节动物门种类占绝对优势,其密度为67.37 ind./m2,占总密度的59.81%;软体动物门种类密度为11.58 ind./m2,占10.28%;节肢动物门种类密度为28.42 ind./m2,占25.23%;棘皮动物门和脊索动物门种类密度相对较低,为4.21 ind./m2和1.05 ind./m2,分别占3.74%和0.93%。图2为春、秋两季大型底栖动物的密度分布,可以看出春季密度大致呈北高南低的趋势,秋季则相反,大致呈北低南高的趋势。总体来说研究区域大型底栖动物密度较低且分布不均匀。

图2 日照潮下带海域大型底栖动物密度平面分布
2.5 生物量及其平面分布
研究区域春季调查中大型底栖动物生物量变化范围较大,在0.10~23.44 g/m2之间,平均为6.21 g/m2。生物量组成以环节动物门种类最高,平均生物量为2.53 g/m2,占总生物量的40.74%;其次为节肢动物门种类,为2.27 g/m2,占36.48%;软体动物门种类平均生物量为1.07 g/m2,占17.22%;棘皮动物门种类平均生物量为0.26 g/m2,占4.15%;脊索动物门种类平均生物量最低,仅0.09 g/m2,占1.45%。秋季生物量在0.32~74.30 g/m2之间,平均为14.75 g/m2,为春季的2.38倍。生物量组成以节肢动物门种类占明显优势,为6.93 g/m2,占总生物量的47.01%;其次为环节动物门种类,为3.38 g/m2,占22.91%;软体动物门、棘皮动物门和脊索动物门种类平均生物量较低,分别为1.14、1.52 g/m2和1.78 g/m2,占比分别为7.73%、10.30%、12.07%。图3为春、秋两季大型底栖动物的生物量分布,两个季节生物量分布都不均匀。这主要是因为部分站位出现了个体较大的物种,如锥唇吻沙蚕(Glyceraonomichiensis)、绒毛细足蟹(Remipestestudinarius)等。

图3 日照潮下带海域大型底栖动物生物量平面分布
2.6 群落多样性
表4列出了两个季节各站位大型底栖动物的H′、J和d值。春季H′值在0.00~3.28之间,平均为1.45;秋季比春季稍高,在0.00~2.45之间,平均为1.66。春季19个站位中有5个站位H′值为0.00,秋季则有3个。从空间分布来看,春季距岸最近的A 断面H′平均值最高,为2.19;距岸最远的F断面次之,为2.12;C断面H′平均值最低,为0.62。秋季距岸较远的E断面H′平均值最高,为2.39,距岸最近的A 断面最低,为1.33。整体来看研究区域H′值处在较低水平,春季H′值呈由近岸海域向远岸海域先降低再升高的趋势,秋季H′值呈由近岸海域向远岸海域逐渐升高的趋势。春季J值在0.72~1.00之间,平均为0.96;秋季比春季略低,在0.79~1.00之间,平均为0.93。两个季节J值均处在较高的水平,这和两次调查各站出现种类数目较少且密度较低有关。春季d值在0.00~0.60之间,平均为0.40;秋季比春季稍高,在0.00~0.77之间,平均为0.44。总体来看,研究区域的d值较低,春、秋季分别有5个和3个站位d值为0.00。其空间分布呈和种类多样性指数相同的趋势。

表4 日照潮下带海域大型底栖动物群落多样性指数
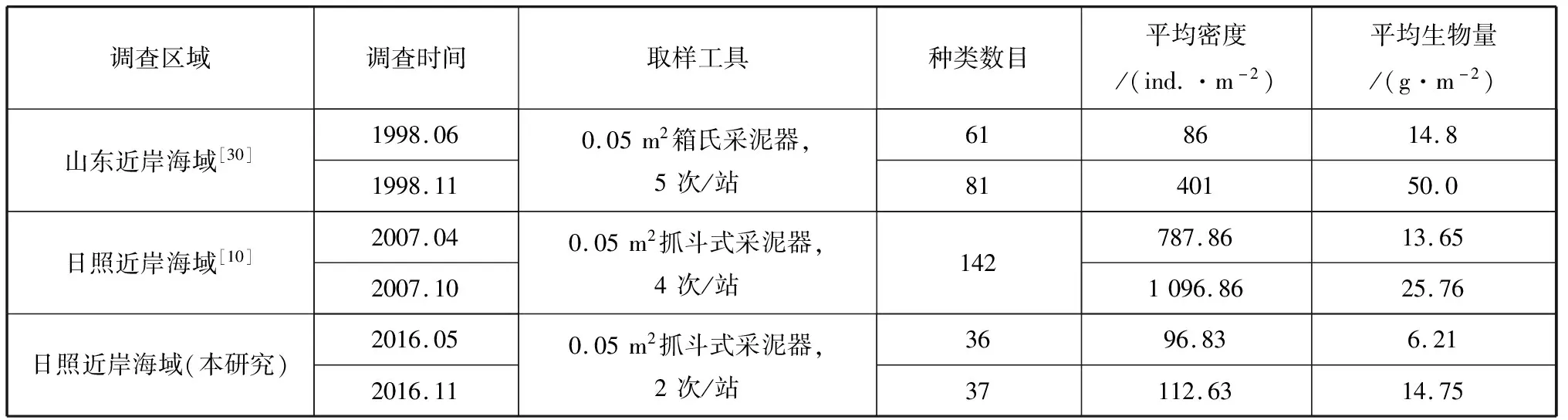
一般来说当环境改变时,生物的群落结构也会发生变化[16]。根据国内外报道大型底栖动物的多样性和所处的沉积物和水质环境有关[17-20]。蔡立哲等(2002)在研究海洋底栖动物种类多样性指数污染程度评价标准时,将其划分为5类:H′>3,清洁;2 春季调查中有5个站位(B3、C2、C3、C4、C5),秋季调查中有3个站位(A2、C4、D2)只出现一个物种,偶然性较大,本研究不再对其作群落相似性分析。图4为研究海域的聚类分析结果,在15%的相似尺度上春季研究海域可划分为4个群落:群落Ⅰ位于调查范围西部靠近海岸线海域,包含A1、A2、A4、B1、C1共5个站位;SIMPER分析表明5个站位平均相似性为20.26%,代表种为醒目云母蛤和管角贝(Siphonodentaliumsp.),二者总贡献率为62.67%;群落Ⅱ位于调查范围东部远离海岸线海域,包含D2、E2、F1、F3共4个站位;站位间平均相似性为20.24%,代表种为锥唇吻沙蚕,其贡献率为46.48%;群落Ⅲ仅包含1个站位D3,不再对其进行SIMPER 分析;群落Ⅳ位于调查范围南部海域,包含A3、B2、E1共3个站位,站位间平均相似性为31.11%,代表种为小头虫,其贡献率为60.71%。在20%的相似尺度上秋季研究海域也可划分为4个群落,其群落Ⅰ、群落Ⅱ和群落Ⅲ都只包含1个站位,分别为B3、A1和C3站位;其余站位为群落Ⅳ。秋季群落Ⅳ站位间平均相似性为28.75%,代表种为索沙蚕科和绒螯近方蟹,其贡献率为88.25%。 图4 日照潮下带海域大型底栖动物聚类分析图 根据聚类分析结果,春季群落Ⅲ和秋季群落Ⅰ、Ⅱ、Ⅲ仅包含一个站位,且出现物种数目小于4,本研究不再对其做ABC曲线分析。图5为春、秋两季各群落大型底栖动物群落ABC曲线,春季群落Ⅰ、Ⅱ、IV和秋季群落IV生物量曲线都位于丰度曲线之下,表明研究区域的整个大型底栖动物群落已受到严重扰动。 图5 日照潮下带海域大型底栖动物群落ABC曲线 2.9.2 大型底栖动物密度和生物量与环境因子的Pearson 相关分析 根据Pearson 相关分析结果,本研究海域春季的大型底栖动物密度和水质中的pH(r=-0.482)和化学需氧量(r=-0.535)呈显著负相关关系(p<0.05),生物量和水质中重金属Cu(r=0.460)呈显著的正相关关系(p<0.05)。秋季的大型底栖动物密度和水质环境因子中的水深(r=0.659)呈极显著的正相关关系(p= 0.001),和水温(r=0.532)呈显著正相关关系(p=0.02);秋季的生物量和所测的20项水质环境因子均未达到显著水平。袁涛萍等(2017)报道了夏季大亚湾的底栖动物种类密度与重金属中Cu、Pb、Cd等环境因素密切相关[28];Mutlu等(2010)研究发现水深对多毛类的种类组成和密度影响显著,这些研究结果与本研究相似[29]。 2.9.3 与历史资料的比较 表5为本次研究结果与本海区历史数据的对比结果。本研究中的大型底栖动物鉴定总物种数目、平均生物量和平均密度在过去的10 a中均呈大幅下降的趋势:其中密度仅为2006年的1/10。分析原因有如下3点:①本研究取样方法为采用0.05 m2抓斗式采泥器,每站采2次,程济生(2004)每站采5次[30],齐磊磊等每站采4次[10],采样强度的差异对以上几个指标都会造成影响;②相比于程济生[30]和齐磊磊等[10],本研究取样范围较小,取样范围的不同会对总物种数目产生影响;③人类活动的干扰:研究区域内分布有海水浴场、渔码头、大型港口和大量的养殖设施,由此产生的生活废水、工业废水、养殖水的循环,水文动力环境的改变都可能会对大型底栖动物群落结构产生干扰。目前已有关于大型底栖动物群落结构由于人为干扰(污染)发生变化的报道:叶属峰等(2004)报道了长江口受经常性疏浚的影响,自1978—2002 年底栖生物种类数目、密度和生物量急剧降低[31]。另有研究表明随着环境污染的加重,棘皮动物会降低甚至绝迹[32]。本研究中两季调查棘皮动物出现频率仅为5.26%和21.05%,这也从侧面印证了研究区域可能已受到了较为严重的人为干扰(污染)。 表5 本研究结果与历史数据对比 (1)本研究在日照潮下带海域,共鉴定大型底栖动物60种,其中环节动物门30种,节肢动物门16种,软体动物门8种,其他类群6种。春季优势种为巴氏钩毛虫、扇栉虫、小头虫和醒目云母蛤;秋季季优势种为绒螯近方蟹、索沙蚕科1种、哈氏美人虾和长鳃树蛰虫。春、秋两季大型底栖动物平均密度分别为96.83 ind./m2和112.63 ind./m2;平均生物量分别为6.21 g/m2和14.75 g/m2;其种类多样性指数和丰度指数均处在一个较低的水平。 (2)综合聚类分析和ABC 曲线分析并结合水质调查结果,认为研究海域大型底栖动物群落受到了严重扰动,底栖生态环境状况处于亚健康状态。 (4)和2006年调查资料相比,研究区域大型底栖动物总物种数目、平均生物量、平均密度均呈下降趋势。2.7 群落相似性

2.8 丰度/生物量比较曲线

2.9 讨论

3 结论

猜你喜欢
今日农业(2021年8期)2021-11-28
金桥(2021年9期)2021-11-02
金桥(2021年9期)2021-11-02
水上消防(2021年3期)2021-08-21
人大建设(2019年5期)2019-10-08
人大建设(2019年3期)2019-07-13
收藏界(2018年1期)2018-10-10
创新作文(小学版)(2018年31期)2018-05-16
摄影之友(影像视觉)(2017年1期)2017-07-18
星星·散文诗(2017年2期)2017-07-05
