
半干旱黄土区不同管理措施下草地群落结构对短期氮、水添加的响应
2021-11-18 02:41:56井光花陈智坤路强强贺丽燕毛祝新
生态学报 2021年20期
井光花,陈智坤,路强强,张 昭,赵 宁,贺丽燕,毛祝新,李 伟
1 陕西省科学院土壤资源与生物技术应用重点实验室,陕西省西安植物园(陕西省植物研究所),西安 710061
2 西北农林科技大学水土保持研究所,杨凌 712100
我国草原面积为3.92×108hm2,约占国土面积的41.7%,是我国最大的陆地生态系统[1],在生态系统功能和服务发挥重要的作用。然而,在长期的过度放牧、乱垦乱挖和人为破坏下,草地退化十分严重。因此,草地资源的管理与保护在草原生态系统中就显得尤为重要。草地管理措施通过调节本地种和非本地种之间植物种类、植株数量的配比的过程直接影响群落结构组成、物种多样性和生产力水平[2- 3]。目前,封育、刈割和火烧是人类在草地生态系统管理实践中的主要措施,对草地群落结构和功能均不同程度地产生了重要影响[4- 6],如封育减少牲畜践踏和啃食,成为恢复退化草地最有效的管理措施之一[5,7];刈割和火烧能去除草地中积累的凋落物,改变草地植物生长环境和空间,从而影响植物群落组成和物种多样性[4]。随着草地管理措施的不断实施,草地群落结构发生显著变化。
氮元素和水分是限制我国半干旱区草地植物生长和发育的关键因子[8- 9],氮添加和水分添加在一定程度上影响草地生态系统稳定性[10- 11]。在全球气候变化背景下,氮沉降和降水变异的增强影响草地生态系统群落功能群组成、物种多样性和生产力,导致草地群落物种多样性和生产力的关系发生变化。目前,关于氮添加和水添加对草地群落物种多样性和生产力的研究进行了大量报道[12- 15]。然而,氮添加和水添加对不同管理措施下草地群落结构影响的研究不足,尤其是在半干旱区黄土高原典型草原研究甚少。
鉴于此,本研究以云雾山国家自然保护区典型草原为研究对象,系统分析封育草地、刈割草地和火烧草地群落地上生物量、功能群组成和群落多样性对氮添加和水添加的响应特征,探讨不同管理措施下草地群落结构对氮添加和水添加的响应机制,为揭示恢复草地生态系统应对全球变化响应机制奠定良好的基础,旨在为黄土高原地区恢复草地生态系统的后续健康和可持续管理提供参考价值。
1 材料与方法
1.1 研究区概况
研究区位于宁夏自治区固原市云雾山草原国家自然保护区(106°21′—106°27′ E,36°10′—36°17′ N),始建于1982年,是典型的黄土高原干旱半干旱草原群落。该研究区属于典型的半干旱气候特征,年均降雨量约400—450 mm,主要集中在7—8月。最热月为7月气温在22—25℃之间,最冷在1月,最低气温能达到-30℃左右,年积温为2370—2882℃。土壤类型主要包括黄绵土、黑垆土和灰褐土。建群种和优势种以长芒草(Stipabungeana)、百里香(Thymusmongolicus)、白莲蒿(Artemisiasacrorum)、大针茅(Stipagrandis)、甘青针茅(Stipaprzewalskyi)和冷蒿(Artemisiafrigida)为主,伴生种类型以猪毛蒿(Artemisiascoparis)、百里香(Thymusmongolicus)和星毛委陵菜(Comarumacaulis)为主,其中丛生禾本科植物长芒草在该区分布范围最广。
1.2 试验设计
本试验选择的样地植被恢复年限为26a,坡度为14°—16°,坡位为东偏北5°属于阴坡,样地为保护区核心区域,地形平缓。在封育样地、刈割样地和火烧样地上分别进行氮添加、水添加和氮添加+水添加,共计12个处理(封育样地:FCK、FN、FW、FNW;刈割样地:CCK、CN、CW、CNW;火烧样地:BCK、BN、BW和BNW),每个处理5次重复,小区面积为6×8 m2。由于试验占地较大,草地立地条件存在差异,在野外试验设计时,尽量选择条件相似的地段作为研究区域。刈割样地是在封育样地上用割草机齐地刈割形成的样地,刈割时间分别为2016年9月和2017年9月。火烧样地是每年3月份对封育样地进行一次性自然火烧处理的小区。从2017年开始,每年5月份选择雨天对施肥小区一次性施加氮肥(尿素),施肥量10 g/m2;水添加量为近50年年均降雨量的20%,约83 mm,于每年4月份分两次进行增水。
1.3 植被调查
每年在植被生物量最大的8月中下旬在小区里选取50 cm×50 cm的样方进行群落生态特征调查,调查的主要指标为群落高度、盖度、多度,同时采集地上生物量和枯落物,根据植物多度和盖度对群落物种进行分类,包括菊科、禾本科、莎草科和杂类草四个功能群。
(1)高度:每种植物随机选5株,直尺测量其伸长高度,求其算术平均值。
(2)盖度:采用目测法测定样方总盖度,针刺法测定每种植物的分盖度。
(3)多度:对样方中出现的植物按种进行分类,同时记录同一种植物出现的次数。
(4)地上生物量:将样方中的所以植物进行按种分类,对地上部分进行地面刈割,除去黏附的土壤、砾石、杂物后按物种装入塑料自封袋,带回实验室备用。在(65±2)℃条件下将物种地上部分烘干至恒重,称其干重。所有物种干重之和为群落地上生物量。
1.4 指标计算
(1)Patrick丰富度指数:
R=S[16]
(3)Pielou均匀度指数:
J=H′/ln(S)[16]
式中,H′为物种多样性,S为样方中的总物种数,pi样方中物种i的个体数占群落中总个体数的比例。
1.5 数据分析
本研究数据在Excel 2013软件进行预处理,采用SPSS软件进行方差分析、相关性分析和回归分析,应用Origin 2019进行图表处理。
2 结果与分析
2.1 氮添加和水添加对不同管理措施群落地上生物量的影响
从图1和表1可以看出,2018年群落地上生物量总体高于2017年,氮添加和水添加对不同管理措施群落地上生物量的影响因年份不同有所差异。图1显示,在封育草地上,2017年氮添加对地上生物量无显著影响,水添加和氮添加+水添加使地上生物量分别增加了10.3%和11.5%。2018年氮添加和水添加处理对群落地上生物无显著影响,水添加+氮添加处理使群落地上生物量提高了6.5%。图1得知,在刈割样地上,2017年氮添加、水添加和氮添加+水添加使地上生物量分别增加了21.7%、13.1%和24.9%;2018年氮添加对地上生物量无显著影响,水添加和氮添加+水添加使地上生物量分别增加了14.1%和23.3%。由图1可以看出,在火烧样地上,氮添加使2017年和2018年群落地上生物量分别增加了18.6%和13.9%,水添加使2017年和2018年群落地上生物量分别增加了12.7%和9.0%,而氮添加+水添加使2017年和2018年群落地上生物量分别增加了30.1%和26.3%。

表1 不同管理措施下草地地上生物量、功能群组成和群落多样性指数年份和处理双因素分析Table 1 Two-factor analysis of year and treatment on aboveground biomass, functional group composition, and community diversity index of grasslands with different management practices

图1 氮添加和水添加对不同管理措施草地群落地上生物量的影响Fig.1 Effects of nitrogen addition and water addition on community aboveground biomass of grasslands with different management practices 不同字母代表处理间在P=0.05水平下差异显著,相同字母代表处理间在P=0.05水平下差异不显著;FCK:封育对照Fencing control;FN:封育和氮添加Fencing with nitrogen addition;封育和水添加Fencing with water additon;FNW封育和氮添加+水添加Fencing with nitrogen and water additon;CCK:刈割对照Cutting control;CN:刈割和氮添加Cutting with nitrogen addition;CW:刈割和水添加Cutting with water addition;CNW:刈割和氮添加和水添加Cutting with nitrogen and water addition;BCK:火烧Burning control;BN:火烧和氮添加Burning with nitrogen addition;BW:火烧和水添加Burning with water addition;BNW:火烧和氮添加+水添加Burning with nitrogen and water addition;N: 氮; W: 水
2.2 氮添加和水添加对不同管理措施草地群落功能群组成的影响
由图2和表1可知,年份和处理共同作用于群落功能群组成比例,且差异显著。2017年禾本科和菊科所占比例高于莎草科和杂草类植物所占比例;2018年杂草类植物所占比例最高,禾本科次之,菊科和莎草科所占比例最少。
图2表明,在封育样地上,2017年不同功能群对氮添加和水添加响应不敏感,差异不显著。2018年氮添加显著增加禾本科、菊科和莎草科所占比例,降低杂草类植物所占比例;水添加显著降低禾本科所占比例,显著增加杂草类植物所占比例,对菊科和莎草科植物所占比例无显著影响;氮添加+水添加使禾本科和莎草科所占比例分别增加了11.9%和15.2%,使杂草类植物降低10.9%,对菊科所占比例影响较小。
由图2可知,在刈割样地上,2017年氮添加使禾本科和菊科所占比例分别显著降低了22.0%和12.2%,使杂草类植物所占比例显著提高了69.8%,对莎草科所占比例无显著影响;水添加降低禾本科和菊科所占比例,提高莎草科和杂类植物所占比例;氮添加+水添加对禾本科所占比例无显著影响,显著降低菊科所占比例,增加莎草科和杂草类植物所占比例。2018年氮添加显著降低禾本科、菊科和莎草科所占比例,显著增加杂草类植物所占比例;水添加使禾本科所占比例显著降低了15.9%,使莎草科和杂草类所占比例分别提高了10.8%和11.9%,对菊科所占比例无显著影响;氮添加+水添加使禾本科和莎草科所占比例分别显著降低了39%和21%,使菊科和杂草类所占比例分别显著提高了25.0%和25.6%。
图2显示,在火烧草地上,2017年氮添加使禾本科和莎草科所占比例分别显著提高了12.5%和49.0%,使菊科所占比例显著降低了36.1%;水添加使禾本科显著降低了25.3%,使莎草科所占比例分别提高了58.8%,对菊科和杂类植物所占比例无显著影响;氮添加+水添加使菊科所占比例显著降低了29.0%,使莎草科和杂草类所占比例分别显著增加51.0%和23.5%。2018年,氮添加对禾本科和莎草科所占比例无显著影响,增加菊科所占比例,降低杂草类植物所占比例;水添加使禾本科所占比例显著降低了15.9%,使菊科和莎草科所占比例分别提高了27.5%和14.3%;氮添加+水添加使菊科所占比例提高了25.0%,使莎草科所占比例显著降低了25.9%。
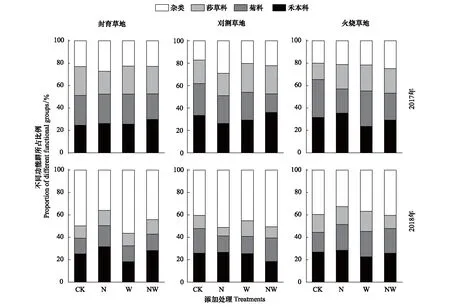
图2 氮添加和水添加对不同管理措施草地群落功能群组成的影响Fig.2 Effects of nitrogen addition and water addition on different functional groups of grasslands with different management practices
2.3 氮添加和水添加对不同管理措施草地群落多样性的影响
图3和表1表明,氮添加和水添加对不同管理措施草地群落Patrick指数、Shannon-Weiner指数和Pielou指数的影响因年份不同而有所差异,且差异显著。
图3表明,在封育草地上,2017年氮添加、水添加和氮添加+水添加使Patrick指数分别降低了18.6%、10.8%和18.8%;2018年氮添加和氮添加+水添加显著降低Patrick指数,水添加使Patrick指数显著降低了13.9%。在刈割草地上,2017年水添加显著Patrick指数;2018年水添加、氮添加+水添加使Patrick指数分别显著提高了21.7%和17.0%。在火烧样地上,氮添加和水添加对2017年和2018年Patrick指数无显著影响,而氮添加+水添加使2017年和2018年分别显著提高了18.1%和23.6%。
由图3可知,在封育草地上,2017年氮添加、水添加和氮添加+水添加均降低Shannon-Weiner指数;2018年氮添加使Shannon-Weiner指数降低了10.2%,水添加使Shannon-Weiner指数显著增加了17.8%,氮添加+水添加对Shannon-Weiner指数无显著影响。在刈割样地上,氮添加提高Shannon-Weiner指数;水添加和氮添加+水添加Shannon-Weiner多样性指数均无显著影响。在火烧样地上,氮添加和水添加对Shannon-Weiner指数均无显著影响,氮添加+水添加显著增加火烧样地Shannon-Weiner指数。
图3还显示,在封育样地上,2017年氮添加和水添加对Pielou指数无显著影响,氮添加+水添加降低Pielou指数;2018年氮添加使Pielou指数降低7.2%,水添加使Pielou指数增加12.0%,氮添加+水添加对Pielou指数无显著影响。在刈割样地上,氮添加使2017年和2018年Pielou指数分别增加了9.3%和12.9%;水添加、氮添加+水添加对2017年和2018年Pielou指数影响较小。从图3和表1可以看出,2017年和2018氮添加、水添加和氮添加+水添加对Pielou指数均无显著影响。

图3 氮添加和水添加对不同管理措施草地群落多样性指数的影响Fig.3 Effects of nitrogen addition and water addition on community diversity index of grasslands with different management practices
2.4 群落多样性与群落结构的关系
由图4可以看出,群落多样性与群落结构的关系因管理措施的不同存在显著差异。在封育草地上,群落Shannon-Weiner多样性指数、Patrick丰富度指数、Pielou均匀度指数均与地上生物量无显著相关,与禾本科所占比例、菊科所占比例和莎草所占比例呈显著负相关,与杂草类所占比例呈正相关。在刈割草地上,Shannon-Weiner多样性指数与地上生物量和杂草类植物所占比例呈显著正相关,与禾本科、菊科和莎草科所占比例呈显著负相关。Patrick丰富度指数与地上生物量和杂草类所占比例呈显著正相关,与禾本科所占比例呈显著负相关,与菊科和莎草科所占比例无显著相关性。Pielou均匀度指数与地上生物量和杂草类植物所占比例呈显著正相关,与禾本科和莎草科所占比例显著负相关,与菊科所占比例无显著相关性。在火烧样地上,Shannon-Weiner多样性指数和Patrick丰富度指数均与地上生物量呈显著负相关,与莎草科所占比例呈显著正相关,与禾本科、菊科和杂草类植物所占比例无显著相关。Pielou均匀度指数与禾本科和莎草科所占比例显著正相关,与杂类植物所占比例显著负相关,与地上生物量和菊科所占比例无显著相关。
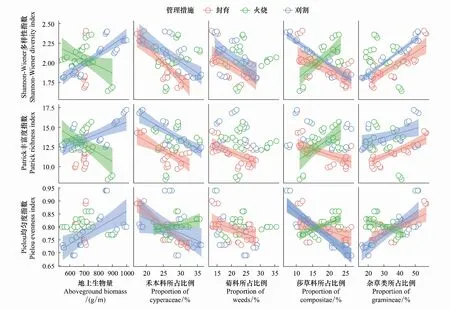
图4 不同管理措施下群落多样性指标与群落结构功能的关系Fig.4 Relationships between community diversity and community structure of grasslands with different management practices
3 讨论
许多研究表明,氮素添加可增加土壤中的有效资源,缓解植物生长的营养元素限制,促进植物地上部分的生长,从而增加地上生物量[17- 19]。不同群落功能群对氮素添加响应的不同,禾本科可高效利用氮素,能快速生长,在群落空间和资源上占据主导地位,从而抑制植株相对矮小的物种,使杂草类植物所占比例降低[17,20- 21];也有研究认为氮素添加在一定程度上抑制了禾本科、菊科植物的生长[22]。大多研究表明,氮素添加促使植物由地下养分资源的竞争转向地上光资源的竞争,一些植株矮小或者不耐阴的物种在竞争中处于劣势,甚至淘汰出该群落,从而降低物种多样性[17,23]。降水量的变化会直接导致初级生产力的年度波动[24],增加供水可减轻或消除水分亏缺对植物生长的限制性,促进了植物生长发育[25]。增水能够提高土壤水分可利用性,促进种子萌发和植株生长,从而提高群落物种多样性和丰富度[17,26-27];也有研究表明增加降水降低物种丰富度[28]或对物种丰富度无显著影响[29]。
3.1 封育草地群落结构对氮添加和水添加的响应
在封育草地上,氮添加处理后,2017年和2018年地上生物量增加幅度较小,差异不显著,表明氮素不是典型草原生产力的限制因子。水添加对草地群落地上生物量的影响因年份不同而有所差异,2017年水添加显著增加草地群落地上生物量,而2018年水添加处理对草地群落地上生物量影响较小,差异不显著,可能是由2017年和2018年降水和气温的差异造成的。然而,氮添加+水添加显著2017年和2018年封育草地群落地上生物量,表明单纯的水分添加或者单纯的氮添加对群落生产力的影响不明显,只有水分和养分交互作用才能显著增加群落生产力。2018年氮添加显著增加禾本科和莎草科所占比例,与禾本科和莎草科氮素吸收利用能力强有关[21];水添加显著增加杂类植物所占比例,表明水分的添加缓解杂草类植物水分的限制,使杂草类植物快速生长;氮添加和水添加的交互作用使强大根系和占据优势地位的禾本科和莎草科所占比例显著增加,显著降低杂草类植物所占比例。此外,水添加处理显著增加群落多样性指数,说明长期封育草地物种多样性受限于水分而非养分。
3.2 刈割草地群落结构对氮添加和水添加的响应
连年刈割带走大量土壤养分和减少草地枯落物的覆盖,增加土壤养分流失和水分蒸发,使得土壤养分和含水量下降,导致草地群落在一定程度上出现贫瘠化和旱化的现象,使得草地群落地上生物量积累量减弱[6,30]。因此,在刈割草地上,氮添加和水添加直接添加补充了水分和养分的流失,改善草地群落土壤微环境,从而提高草地群落地上生物量和增加物种多样性。从2017—2018年数据看,氮添加和水添加均不同程度地增加刈割草地群落地上生物量和物种多样性,氮添加和水添加交互作用增加幅度最为显著,说明氮添加和水添加的交互作用是累加效应的。不同功能群对氮添加和水添加处理的响应有所差异,禾本科和菊科所占比例不同程度地降低,而杂草类所占比例显著增加,可能原因是禾本科和菊科植物对氮添加和水添加响应较慢,而杂草类植物对氮添加和水添加响应敏感,能够快速生长,生态位增加,从而提高杂草类所占比例。
3.3 火烧草地群落结构对氮添加和水添加的响应
连续火烧使土壤养分通过土壤加热、植物吸收和潜在淋溶等方面流失[31-32],火烧后凋落物中的绝大多数氮以气态形式损失掉[33],土壤肥力下降,从而降低群落地上生物量的积累。在火烧草地上添加氮肥和水分,补充连续火烧引起的土壤干旱和养分流失,从而显著增加群落地上生物量和物种多样性。从2017年和2018年数据可以看出,氮添加和水添加对物种多样性指数影响较小,但显著提高群落地上生物量。氮添加+水添加使群落地上生物量和物种多样性增幅最为显著,表明氮添加和水添加的交互作用具有累加效应。然而,不同功能群对氮添加和水分添加响应因年份不同存在显著差异。例如,2017年氮添加+水添加显著增加莎草科和杂草类所占比例,而2018年氮添加+水添加显著降低莎草科所占比例,可能原因是2017年和2018年降水的差异造成土壤水氮失衡程度不同,从而导致不同功能群对水氮添加的响应有所不同。
3.4 群落多样性和功能群组成与地上生物量的关系
目前,关于物种多样性与生产力关系开展了大量研究[34-36],但植物群落多样性和生产力之间的相关格局没有统一的结论,大致争议为正相关、负相关和不相关关系。在全球气候变化背景下,大气元素循环和降水格局的改变往往会影响草原生态系统植物物种多样性及其生产力[37],导致草原生态系统植物群落物种多样性及其生产力间关系发生变化。由于群落间养分水分受限差异不同,有研究认为养分添加后地上生物量与群落多样性呈负相关[4, 8],也有研究认为植物群落地上生物量与物种多样性指数和物种总丰富度呈显著正相关[19]。在本研究中,在封育草地上,氮添加和水添加群落地上生物量与群落多样性指数无显著相关,但与杂草类所占比例显著正相关,说明长期封育草地群落多样性受群落结构组成的影响。刈割草地氮添加和水添加后,物种多样性指数与地上群落生物量和杂草类所占比例呈显著正相关,说明长期封育草地刈割后,氮添加和水添加通过增加杂草类所占比例,提高物种多样性,从而增加地上群落生物量,可能是因为物种丰富度会增加物种功能性状的空间维度,提高光或者土壤养分资源的利用率,从而促进地上生物量的积累。火烧草地氮添加和水添加后,物种多样性指数和地上群落生物量呈显著负相关,但与莎草类所占比例呈显著正相关,说明长期封育草地火烧后,氮添加和水添加通过增加莎草类所占比例提高物种多样性。
4 结论
氮添加和水添加对草地群落结构和功能的影响因管理措施不同存在显著差异。在封育草地上,氮添加显著降低物种多样性,而水添加显著增加物种多样性指数;物种多样性指数均与地上生物量无显著相关,与不同功能群所占比例显著相关。在刈割草地和火烧草地上,氮添加和水添加显著提高草地群落地上生物量,氮添加和水添加交互作用具有累加效应;然而,氮添加和水添加对群落多样性指数与功能群组成的影响差异显著。在刈割草地上,Shannon-Weiner多样性指数与地上生物量和杂草类植物所占比例呈显著正相关;在火烧草地上,Shannon-Weiner多样性指数与地上生物量呈显著负相关,与莎草科所占比例呈显著正相关。结果表明,刈割和火烧改变长期封育草地养分水分含量和群落构成,在刈割草地和火烧草地氮添加和水添加后,物种多样性指数和群落功能群的响应存在显著差异,从而影响物种多样性指数和群落结构的关系。
本试验研究半干旱黄土区不同管理措施下草地群落结构对短期氮添加和水添加的响应特征,旨在为恢复草地生态系统的后续健康和可持续管理提供参考价值。由于试验时间较短,氮添加和水添加对草地群落结构的影响具有滞后性,因而存在一定的局限性。因此,未来研究时持续关注恢复草地不同管理措施下对氮添加和水添加的响应机制,同时注意功能群组成和管理措施的差异对生产力和群落多样性关系的影响。
猜你喜欢
天津农林科技(2022年2期)2022-04-19 10:48:14
河南畜牧兽医(2022年3期)2022-04-13 09:12:58
绿色科技(2021年10期)2021-06-23 03:13:06
草业学报(2019年2期)2019-02-25 01:57:16
天然产物研究与开发(2018年10期)2018-11-06 07:43:46
江苏农业科学(2016年3期)2016-05-03 15:00:58
当代化工研究(2016年5期)2016-03-20 16:21:30
湖南城市学院学报(自然科学版)(2016年4期)2016-02-27 14:02:41
农产品市场周刊(2015年39期)2015-10-31 19:12:27
山东农业科学(2014年8期)2014-10-10 11:24:32
