
桂林岩溶石山青冈群落植物功能性状的种间和种内变异研究
2021-11-18 02:42:26姚玉萍姚义鹏梁士楚李月娟梁欢欢赵庆年黄勇焙林才健
生态学报 2021年20期
何 雁,姚玉萍,姚义鹏,姜 勇,*,梁士楚,李月娟,梁欢欢,赵庆年,黄勇焙,林才健
1 广西高校野生动植物生态学重点实验室,广西师范大学,桂林 541006
2 广西师范大学生命科学学院,桂林 541006
3 桂林航天工业学院,桂林 541004
一个多世纪以来,生物多样性的形成和维持机理即群落构建机制一直是生态学研究的核心论题。相比大多数基于植物分类和数量的研究,植物功能性状考虑了共存物种的冗余和互补,反映了物种在功能属性方面的差异,已成为揭示群落物种共存和多样性维持机制的可靠途径[1- 2]。植物功能性状是指植物体具有的与其定植、生长、存活和死亡等紧密相关的一系列核心植物功能属性,且这些属性与环境适应后表现出的性状变异不仅能够显著影响生态系统功能,而且可以将群落结构与群落环境、生态系统过程等联系起来[3- 4]。通过植物功能性状可以探明环境变异与物种分布之间的机理性关联[5- 7],而探讨功能性状在种间和种内尺度上是否存在差异及其形成原因是理解群落构建的重要环节[8]。
目前,已有大量的研究表明种内变异对群落动态变化以及生态系统特征具有显著的影响,其与种间变异同等重要[9- 10]。Siefert等[11]对植物群落种内性状变异相对程度的全球荟萃分析表明,种内性状变异平均占群落内总性状变异的25%,占群落间总性状变异的32%。不少研究表明,种内性状变异可能是物种定居新环境或抵抗环境变化能力的主要因素[12- 13]。种内性状变异可通过遗传变异和表型可塑性来响应局部选择压力使物种内性状发生适应性变化,相较于利用物种性状均值进行的种间研究更能反映出植物种群的性状变异能力[14- 15]。而Niu等[16]对西藏高山草甸的研究也表明了,种内性状变异在植物群落中的功能重要性会随着环境的恶劣程度而增加。因此,将种内性状变异和种间性状变异相结合才能更真实地反映群落构建过程中物种对生境变化和资源竞争的响应,并更准确地理解物种多样性维持机制及生态系统特征[17- 19]。基于功能性状的群落构建机制通常将群落视为是确定性过程或随机性过程或两者结合的物种分选过程的结果[20- 22]。确定性过程侧重于两个选择性过程:环境过滤和相似性限制(生态位分化)[23]。环境过滤被视为定向选择的一种形式,即非生物环境选择特定的性状值,从而导致生境内的性状趋同[24]。相似性限制则被视为密度制约的一种选择形式,通过竞争和其他生物相互作用以避免与相邻物种的性征值过于相似[25]。而如果群落主要受到扩散限制的影响,则随机过程将起主导作用[21,26]。通常,检验群落内物种共存机制的方法可利用实际观测的性状分布与零模型随机模拟得出性状分布进行比较[27]。近几年来基于功能性状结合零模型探讨群落构建机制的研究已经有很多,但检测水平大多还停留在物种水平、种群水平[18,28],综合考虑种内变异基于个体水平的零模型检测还少有报道。
桂林岩溶石山地貌区生态系统特征典型,其岩溶发育强烈、生态系统稳定性差、岩石裸露率较高,地形破碎、土层浅薄、土壤贫瘠、水分渗漏严重和石漠化加剧,是世界岩溶区域演化最具典型性和普遍性的代表地之一[29- 31]。由于该区域的生态环境恶劣,植物的生长与繁殖受到了限制[32]。青冈(Cyclobalanopsisglauca)为壳斗科青冈属常绿阔叶树种,具有较强的耐贫瘠、耐旱性、石生性和嗜钙性等特点,容易成为岩溶生态系统顶极群落的建群种,在岩溶森林植被中具有重要的地位和作用[33- 34]。鉴于此,青冈群落为探讨岩溶森林植被的性状变异及其背后的生态学机理提供了良好的研究对象。通过以桂林岩溶石山青冈群落木本植物为研究对象,基于4个植物功能性状(叶干物质含量LDMC、叶厚度LT、叶面积SLA和木材密度WD),采用混合线性模型结合方差分解的统计检验分析功能性状在种间和种内2个尺度变异程度及比重;其次,针对其在种内和种间2个尺度的变异及比重的不同,基于零模型(null model)方法检验桂林岩溶石山青冈优势群落构建机制,主要探讨以下3个科学问题:(1)植物功能性状的种间和种内变异程度及比重如何?(2)结合个体性状值将种内变异纳入考虑的群落机制检测是否得到改善?(3)桂林岩溶石山青冈群落构建过程中的潜在驱动机制是什么?通过对以上问题的研究,希望能初步揭示桂林岩溶石山青冈群落种内种间性状分布模式和维持机制,为桂林岩溶石山森林保护与管理提供科学依据。
1 材料和方法
1.1 研究区概况
研究区位于桂林岩溶区处于演替后期的青冈群落内,地处广西壮族自治区东北部(110°14′—110°42′ E,24°43′—25°20′ N)。该区域属于中亚热带湿润季风气候,雨量充沛,气候温和,具有显著的岩溶地区特性,年平均气温为18—19 ℃,最冷1月份平均气温为8 ℃,最热8月份平均气温为28 ℃,全年无霜期达309 d,年降雨量为1856.7 mm,降雨量全年分配不均匀,春夏湿润多雨,年平均蒸发量为1458.4 mm[35]。该区域乔木层主要以青冈为建群种,伴生种主要有扁片海桐(Pittosporumplanilobum);灌木层主要以一叶萩(Flueggeasuffruticosa)、粗糠柴(Mallotusphilippensis)、红背山麻杆(Alchorneatrewioides)、干花豆(Fordiacauliflora)、檵木(Loropetalumchinense)等为主;木质藤本以龙须藤(Bauhiniachampionii)为主。
1.2 群落调查
2017年7—9月在桂林阳朔县葡萄镇碑头村(BTC)、桂林阳朔县白沙镇富里湾(FLW)、桂林市郊芦笛岩张家村(ZJC)和桂林市郊演坡山(YPS) 4个采样点共建立了20个20 m×20 m的样方,各样方基本概况如表1所示。调查时,将每个20 m×20 m的样方划分成4个10 m×10 m的小样方,对每个小样方进行常规群落学调查,内容主要包括:对乔木层植株进行每木检尺,记录种名、空间坐标、高度、胸径、基径、冠幅等指标;灌木层植株记录种名、空间坐标、高度、基径等指标。同时,记录各样方的土壤类型、海拔、岩石裸露率、砾石直径、干扰程度等生境特征。所有样方共记录了4190株植物个体,隶属36科,57属,105种。其中乔木2372株,灌木1818株。

表1 样地基本概况Table 1 Basic information of different sampling sites
1.3 功能性状数据采集和测定
1.3.1功能性状数据采集
由于植株的叶片生长状况、个体发育阶段、冠层位置以及荫蔽度等会对植物的叶性状差异造成影响,因此,本研究中采集了样方内所有胸径≥1 cm的木本植物个体树冠顶端3片完全展开的健康向阳叶片,为避免年份和季节等时间因素造成植物的功能性状变异,研究数据均于2017年7—9月采集完成。木材样本采自与叶片样本相同的个体,使用枝剪截取树冠外围3小段非当年生小枝。为保持叶片和枝条的水分饱和,室内功能性状测定均在样本采回8小时内完成。本研究共计有12570片叶片和木材样品,来自105个种的4190株植物个体。
1.3.2功能性状数据的测定
本研究选取叶干物质含量(LDMC)、叶厚度(LT)、比叶面积(SLA)和木材密度(WD) 4个功能性状进行植物功能性状的种间和种内变异研究。其中,SLA和LDMC是叶片经济谱的核心性状,代表植物获取资源的能力;LT决定着叶片物理抵抗能力;WD是与机械稳定性、生长速率以及耐旱和耐荫策略相关的关键性状[15, 36]。功能性状各指标测定方法如下:LDMC用精度为0.0001 g的电子天秤(BSM- 220.4,Zhuojing,China)称量叶片的鲜重,然后将叶片样品编号置于80 ℃烘箱中烘48 h 至恒重,称量并记录其叶片干重,根据叶片干重(g)与叶片鲜重(g)的比值获得叶干物质含量;LT用精度为0.01 mm的电子数显卡尺(SF2000,Guilin,China)测量其叶厚度,每个叶片避开叶片主脉测量其前端、中端和末端,然后将三者的平均值作为所测叶片的叶厚度值;SLA由便携式叶面积仪(Yaxin- 1241,Yaxin,China)测得叶片叶面积,由电子天秤测得烘干后的叶片干重,根据叶片面积(cm2)与叶片干重(g)的比值获得比叶面积;WD根据Swenson和Enquist的研究[37],利用测定的枝条密度代替木材密度,即取回的枝条将其表面的树皮削去后,用电子天平称量并记录枝条鲜重,利用量筒排水法测其体积,然后对其编号并放入80 ℃烘箱中烘干至恒重后,称量并记录枝条干重,根据枝条干重与体积的比值计算枝条密度(g/cm3)。
1.4 土壤取样和理化性质测定
本研究选取土壤含水量(SWC)、有机质(SOM)、pH值(pH)、全氮(TN)、速效氮(AN)、全磷(TP)、速效磷(AP)、全钾(TK)、速效钾(AK)等9个土壤理化性质进行测定。在20 m×20 m的样方中心位置处用环刀采集0—20 cm的土样用于测定土壤含水量。土壤化学成分则采用“梅花五点法”采样,在每个样方的四个顶点和中心位置清除表层枯落物和腐殖质层后,用土钻钻取0—20 cm表层土样1 kg左右,运用四分法混合均匀后装入密封袋带回实验室用于土壤化学性质的测定。将采集的新鲜土样放置于室内阴凉通风处自然干燥,待风干后,剔除其中的根系、石块、钙核及动植物残体等杂物后倒入研钵中研细,过不同孔径(0.15、0.25、1.4 mm)的土壤筛,对预处理后的土样进行分析测定。测定方法参照《土壤农业化学常规分析方法》[38]。每个土壤样品重复测定3次后取其平均值作为本研究分析的数据。
1.5 统计与分析
首先,本研究筛选出个体数大于3的71个主要物种进行植物功能性状的种间和种内2个尺度上的变异分析。考虑数据需要满足方差齐性和正态性,本研究分别对叶干物质含量(LDMC)、叶厚度(LT)、比叶面积(SLA)和木材密度(WD)这4个性状进行对数转换。利用混合线性模型和方差分解方法分析其在种间和种内2个尺度上贡献程度。该模型通过限制最大似然来拟合,相应的性状作为响应变量,两个尺度(即种间和种内)作为嵌套随机因素。方差成分之间的比例表示每个尺度导致的变化的比例贡献。针对4个性状在种间和种内2个尺度上贡献的不同,本研究采用①性状的范围(Range)矩阵(作为检验环境筛选的标准)和②性状轴上最近邻体距离变异系数(CV_NND)矩阵(作为检验相似性限制的标准)在种间和种内2个尺度上进行观测值与零期望9999次随机模拟值比较分析。预计环境过滤和相似性限制会使观测到的性状值范围和最近邻体距离变异系数低于零期望值,本研究采用Wilcoxon符号单侧秩检验法比较观测群落与随机群落是否存在显著差异性,进而从统计学角度验证群落构建机制理论。考虑到环境过滤对相似性限制检验可能产生的附加影响,在随机筛选性状数据时执行一个“限制零模型”,即从受约束的物种库中随机筛选物种性状的分布范围与指定群落性状分布范围相同。理论上,与随机分布相比,环境过滤在理论上会改变实际群落的性状均值以及引起性状的范围变窄;而相似性限制则会使物种沿性状轴分布得更加均匀,因而会降低实际群落的最近邻体距离变异系数[22, 39]。对于每个群落,利用SES值(standardized effect size)量化了实际群落性状分布的标准值对随机群落性状分布零期望偏离的程度:
SES=(Iobs-Inull) /σnull
(1)
其中,Iobs代表实际群落的观测值,Inull和σnull分别代表随机群落的平均值和标准差,而SES值的正负则刻画了实际群落性状分布的标准值对随机群性状分布零期望偏离的方向。
功能性状与土壤因子的关系采用多元逐步线性回归分析,以土壤含水量(SWC)、土壤有机质(SOM)、土壤pH值(pH)、全氮(TN)、速效氮(AN)、全磷(TP)、速效磷(AP)、全钾(TK)、速效钾(AK)等9个土壤理化性质指标为自变量,以叶干物质含量、叶厚度、比叶面积、木材密度的群落加权平均性状值(CWM)为因变量,根据AIC值、模型决定系数(R2)及P值大小选择最优模型,筛选出影响桂林岩溶石山青冈群落植物功能性状的关键环境因子。以上所有数据统计分析与制图均在R 4.0.2软件中完成,其中混合线性模型的拟合用“nlme”包,方差分解用“ade”包的“varcomp”函数,性状的群落加权平均计算用“FD”包。
2 结果和分析
2.1 植物功能性状的分布和变异
如表2所示,4个功能性状的变异程度存在很大的差异,其中SLA的变异最大(60.81%),其次是LT(37.58%)和LDMC(25.13%),WD的变异最小(21.03%)。而方差分解和混合线性模型结果表明:4个植物功能性状在种间和种内2个尺度变异的大小表现出一致的规律(图1),均体现种间>种内。三个叶性状(SLA,LT和LDMC)的变化主要来源于种间(50.42%—71.90%),种内的贡献较小(23.65%—42.61%);而木材性状(WD)的变化在种间(47.82%)和种内(44.86%)的贡献相近。总体来看,桂林岩溶石山青冈群落植物功能性状的种间变异(59.48%)要高于种内变异(34.61%),种间变异是植物功能性状变异的主要来源,但种内变异亦不容忽视。

表2 4个植物功能性状的分布特征Table 2 Distribution characteristics of 4 plant functional traits
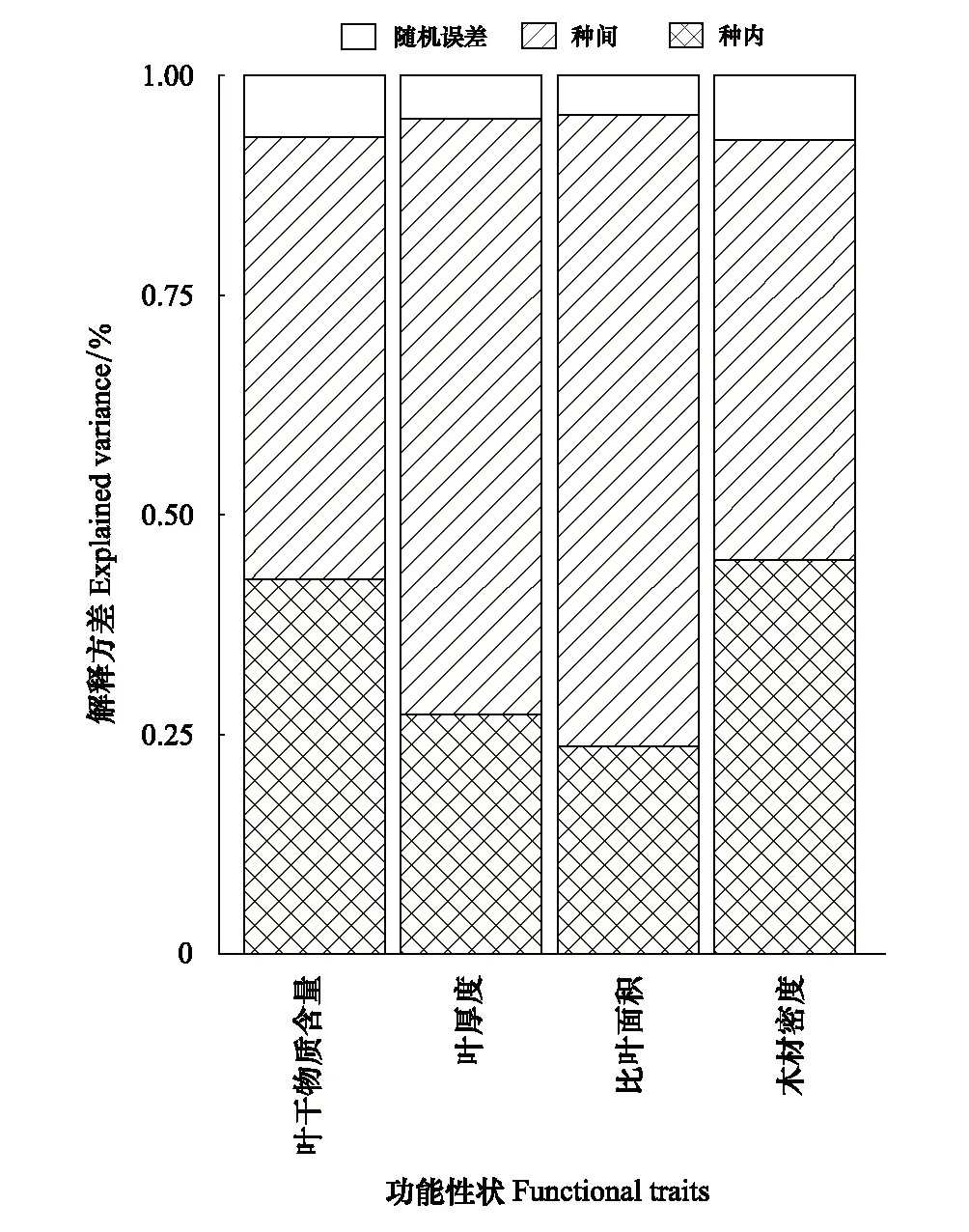
图1 不同功能性状在种间和种内尺度上的方差分解图 Fig.1 Variance partitioning of different functional traits at interspecific and intraspecific scales
2.2 零模型检验
如图2所示,在结合物种性状均值种间尺度上的环境过滤检验中,SLA和WD的分布范围与随机零模型相比显著减小,呈现聚集分布,表明环境因子对植物的性状分布具有限制作用;但是LDMC和LT的分布范围与随机零模型相比差异性不显著。在种间尺度的相似性限制检验中,仅WD的最近邻体距离变异系数与随机零模型相比显著减小,呈现均匀分布,这与相似性限制理论预测的结果一致;但其余性状与随机零模型相比差异不显著,呈现随机分布。

图2 功能性状在不同群落的零模型比较Fig.2 Comparison of null models of functional traits in different communities*代表观测群落与零模型随机群落在Wilcoxon符号秩检验下呈显著差异,* P <0.05,*** P <0.01,ns代表差异不显著
在结合个体性状值的种内和种间尺度的环境过滤检验中,LDMC,LT和SLA的分布范围与随机零模型相比显著减小,呈现聚集分布;WD的分布范围与随机零模型相比差异不显著。在种内和种间尺度上的相似性限制检验中,LDMC,LT,SLA和WD的最近邻体距离变异系数与随机零模型相比差异都不显著,呈现随机分布。
2.3 功能性状与土壤因子的关系
多元逐步线性回归分析结果表明,桂林岩溶石山青冈群落植物功能性状与土壤有机质、含水量、pH和氮磷钾含量等环境因子密切相关(表3)。其中,叶干物质含量与土壤含水量、全磷和全钾含量显著相关;叶厚度与土壤全磷、含水量、有机质和全钾含量显著相关;比叶面积与土壤pH显著相关;木材密度与土壤全磷、全氮和全钾含量显著相关。

表3 功能性状与土壤养分多元逐步回归分析Table 3 Multiple linear stepwise regression analysis of function traits with soil nutrients
3 讨论
3.1 植物功能性状的种间和种内变异分析
从研究结果来看,不同性状的变异程度表现出了较大的差异,这表明不同功能性状受到的环境作用的影响程度是存在差异的。通过4个性状在种内和种间水平上的变异检测,进一步表明了桂林岩溶石山青冈群落植物功能性状的主要变异来源是种间变异。其次,相比木材性状(WD),三个叶性状(LDMC,LT和SLA)拥有更高的种间变异。植物性状是由遗传因素与环境条件共同决定的[40],不同遗传背景的物种(分类单元不同)其性状在种间差异较大,尤其是叶性状[41]。本文叶性状(SLA,LDMC和LT)在种间变异分析结果符合上述客观规律,说明其主要受到遗传因素限制,具有稳定的变化特征。另一方面,本研究发现性状的可塑性降低可能是由于倾向不利生境的专性化而引起的,表明低变异的物种更常见于恶劣的生境[42]。桂林岩溶石山相对恶劣的生境特点,导致了其更倾向于选择性状可塑性低的物种[10, 33],这意味着植物对特殊环境的变化敏感,因而可能存在特殊的适应环境方式。但木材性状WD种内变异的贡献接近其种间的贡献,根据生态位理论,植物功能性状的变异程度受环境过滤和相似性限制作用的共同影响[43- 44]。环境过滤作用能够降低性状的变异程度,使得具有相似性状的物种在局域生境中共存,共存物种的性状在功能上往往表现出趋同的一面,而相似性限制作用使这些共存物种通过种内变异来增强生态位的分化,从而呈现某些生态策略的趋异性来降低生物竞争强度[28, 45]。对比本研究中的4个植物功能性状,种间尺度的变异虽然占据了主导地位,但也有如WD种内变异贡献(44.86%)接近其种间贡献(47.82%)的性状。这表明了性状的种内变异并非是无足轻重,而是重要且不容忽视的变异来源之一。故在今后的基于性状的生态学研究中,不能简单地使用物种水平的功能性状平均值代替个体数据而忽略种内变异,应基于个体水平采样并整合功能性状的种内和种间变异来研究植物对环境的适应策略,以期更好地揭示植物群落构建和生物多样性维持机制。
3.2 种内种间变异对群落构建的响应
通过比较零模型结果来看,种内变异对改善群落构建机制的检测有着一定的影响。基于个体水平将种内变异纳入考虑的环境过滤检测确实得到了改善,三个叶性状都表现出更强的收敛模式,极显著低于零模型模拟。但与Siefert等[19]在美国纽约废弃农田的植物群落中的实验结果相似,在使用个体性状数据对相似性限制检测没有得出相似的结论,性状在局域群落中更趋向于随机分布而不是均匀分布。这一研究结果表明在桂林岩溶石山青冈群落种内和种间尺度上性状分布除了受到环境过滤这个确定性过程的主要影响外,还受到一定的以扩散限制为主的中性随机过程的影响。这可能与桂林岩溶石山相对特殊(极端)的生境有关。一方面,研究表明物种在所处生境受到的胁迫性越高所需的耐受性也就越高,那么其性状专化程度亦会相应的表现得较高[42, 46],因此岩溶区的植物群落容易表现出较强烈的环境过滤作用;另一方面,正如Conradi等[47]在温带草原上的田间试验发现:随机过程是营养贫瘠土壤上群落物种组成的有效驱动力,因此在岩溶石山相对贫瘠的土壤环境上随机过程必然也在起着一定的作用。同时,通过多元逐步线性回归的功能性状与土壤因子的关系分析,发现土壤有机质、含水量、pH和氮磷钾含量等环境因子与功能性状之间密切相关,这一结果也很好地佐证了环境过滤在桂林岩溶石山青冈群落构建中的驱动作用。
此外,本研究发现在种间尺度的零模型检验中,WD不仅体现环境筛选,同时体现限制相似性作用,但在种内和种间尺度的零模型检验中WD却显示随机的分布格局。根据Albert等[48]提出的“性状空间变异分割假说”,随着空间尺度的增加,环境异质性和个体数量逐步增加,表型可塑性也随之增加,从而导致种内变异的程度逐步增大,而种内变异与环境过滤过程呈负相关,与相似性限制作用成正比。因此,本研究推断这可能是由于考虑了种内变异,相似性限制作用的检测得到了加强,其与环境过滤的作用力相互抵消,导致了随机格局的产生。尽管确定性机制似乎是群落构建的重要因素,但在自然系统中,只要其他因素施加有意义的影响,它们的信号就可能被稀释和掩盖,要令人满意地解释在自然群落中观察到的群落构建模式,必须综合考虑确定性过程和随机过程的共存模式[49]。共存理论的最新发展认识到,随机性过程和基于生态位的确定性过程在塑造群落中同时起作用[43, 50],但是要恒量这两个过程在不同物种组,不同生境和生物群落以及不同空间尺度的相对贡献仍然是一个困难。因此,在今后基于功能性状的生态学研究中,注重在个体水平采样将种内性状变异和种间性状变异相结合的同时,亦要注重构建生态位分化的确定性过程和中性作用的随机性过程的耦合关系模型,量化环境过滤、相似性限制和扩散限制作用的相对贡献率,结合空间结构、环境因子和系统发育深入探究植物功能性状变异的来源与潜在驱动机制,以期更好地揭示植物对环境的适应策略以及植物群落构建和生物多样性维持机制。
4 结论
综上所述,本研究的结论如下:(1)桂林岩溶石山青冈群落植物功能性状的种间变异最大,但种内变异亦不容忽略;(2)基于个体性状值来考虑种内变异是有必要的,虽然没有提高相似性限制作用的检测能力,但环境过滤作用的检测明显得到了改善;(3)对于桂林岩溶石山青冈群落,环境过滤作用确实是群落构建的重要驱动机制,与相似性限制作用和随机作用共同塑造了以青冈为优势物种的群落构建,不过要具体量化不同过程在群落构建过程的相对重要性还是一个困难。因此,未来的研究中,应注重在个体水平采样将种内性状变异和种间性状变异相结合,进一步构建生态位分化的确定性过程和中性作用的随机性过程耦合关系模型,不断引入和开发新模型新方法来量化环境过滤、相似性限制和扩散限制作用的相对贡献率,加强群落构建机制的定量分析研究,为揭示植物对环境的适应策略和理解群落构建的具体过程提供深刻的见解。
猜你喜欢
歌海(2024年2期)2024-06-06 05:54:00
故事作文·高年级(2023年10期)2023-10-23 11:21:22
植物研究(2023年5期)2023-09-09 08:01:22
河南师范大学学报(自然科学版)(2022年5期)2022-08-08 14:07:42
湖南水利水电(2021年6期)2022-01-18 06:07:02
中华建设(2019年2期)2019-08-01 05:57:58
水利科技与经济(2016年9期)2016-04-22 01:07:50
杂草学报(2015年2期)2016-01-04 14:57:55
小主人报(2015年1期)2015-03-11 19:40:59
大众文艺(2015年3期)2015-02-22 09:15:09
