
城市公园春季声景观与植被结构的关系
2021-11-18 02:32:42赵伊琳白梓彤殷鲁秦孙振凯孙睿霖孙宝强
生态学报 2021年20期
赵伊琳,白梓彤 ,王 成,*,殷鲁秦,孙振凯,张 昶,孙睿霖,徐 诗,边 琦,孙宝强
1 中国林业科学研究院林业研究所,国家林业和草原局林木培育重点实验室,北京 100091
2 国家林业和草原局城市森林研究中心,北京 100091
3 天津财经大学艺术学院,天津 300222
“月出惊山鸟,时鸣春涧中”,“百啭千声随意移,山花红紫树高低”,正如古典文学中所描述的,声景观,尤其是生物声景观,早在古代就受到了人们的关注。在古典园林中,“声音”成为了人们营造意境的重要手段。无锡寄畅园的八音涧是古人营造声景观,表现自然幽谷溪涧声音之美的典型。可见,从古至今,人们对于景观的要求都不止局限在视觉上,优美的声景观能带给人更丰富而立体的体验[1]。
“声景观”——来自景观中所有声音的集合——这一术语在1969年首次被Southworth 在城市规划领域文献中使用[2-3]。声景观不是恒定的,其在空间和时间上都是异质的。这种变化在每个生态系统(自然和城市)中都很普遍,可以通过特定的声学特征来定义和表征[4]。“声景生态学”这一研究领域及其理论框架于2011年由Bryan C. Pijanowski等正式提出[7],该领域强调声音的生态特性及其时空模式[3]。声景生态学研究通过记录和量化声音信息为解析区域景观和生态特征、长期监测动物群落动态、评估环境生物多样性水平和生态完整性提供了新的研究理论与视角[5]。
将声学数据从时域转换为频域(如通过傅里叶变换),提供了研究景观中有机体信号流动的新思路。声学指标可以作为跨时空尺度的生态系统功能的表征,能够客观地描述或评估声景观总体的表现和特征[8],作为表征生态系统功能的度量,其比传统的物理调查具有更小的侵入性和更高的准确度[6]。Boelman等开发的生物声学指数(BIO),是录音中频谱振幅和频带数目的函数[9],该指数被证明与夏威夷森林中的鸟类数量密切相关[10-11]。声学多样性指数(ADI)[12]计算每个1 kHz频带信号功率占用的香农多样性指数。能够有效反映声学活动总体差异,例如黎明合唱和夜间活动之间或不同生境之间的区别[13];归一化声景指数(NDSI)[14]已被证明可以反映景观中的季节性和昼夜变化,能够描述动物与人类之间的相互作用[14],也被证明与生境中鸟类的存在有关[15]。
由于声景生态学仍处于起步阶段,因此存在许多研究空白,对这些部份的研究将使声景技术在长期而有效的生态系统功能和生物多样性监测以及城市或绿地声景观规划与管理方面从理论变成实际的应用。随着城市快速发展,越来越多的自然景观转换为城市景观,在人类主导的系统中保持生态完整性将是一项至关重要的管理挑战。因此,了解声景观时空动态及其在城市地区中应用于声景观监测的潜力至关重要[16]。
但迄今为止,大多数声景观研究在自然保护区或森林中进行,很少有研究描述高度城市化地区城市公园的声景观特征。此外,在城市尺度声景观的研究对植物特征的提取常常停留在景观类型[16-17]、归一化植被指数[17]或植被盖度[18]等较宏观的水平分布特征。而生境是生物活动的直接环境,乔木群落的垂直异质性与分层结构的复杂性能够极大的增加生物的栖息空间[19],目前仍缺乏对于植被结构的标准量化方法,这可能会影响对于植物环境与生物声景观关系的研究。因此,在本研究中,记录并描述了北京五环内二十个城市公园的声景观特征。此外,为了确定植被群落结构对生物声景观的影响,将植被因子降维后用于植被结构类型的划分,解析植被结构与生物声景观的关系。这些生态声学指数随后可服务于城市规划者、社区决策者与开发商,以实现科学管理绿地声环境及监测声景观变化。
1 研究方法
1.1 研究地点选择
北京(115°24′—117°30′E,39°38′—41°05′N)是中国的首都和世界第二大城市。平均海拔43.5m。在城市发展过程中,北京城区形成典型的同心环结构,城市化梯度随着环数增加而逐渐减小。截至2015年底,北京市常住人口达到2170.5万人[20],人口密度随着环数量的增加而逐渐减小。五环(城区)内的绿化面积占城市总面积的32.8%。城市公园是北京最重要的绿地。这些公园营建主要面向市民使用,同时兼具城市生态系统保护与生物多样性改善的作用。因此,有必要研究城市公园不同植被结构下生物声景观特征及城市绿化规划与管护策略。北京五环内,沿北-南、西北-东南、西-东、西南-东北截面,根据北京市环状分布格局从2环到5环(位于五环外沿的奥林匹克森林公园北园除外),选择20个公园(图1),面积从2 hm2到200 hm2不等。此后,基于公园面积聚类结果,在各公园内选择1—5个固定样点,共计60个。各样点所处的坡向、海拔高度相似,以便于后续比较分析。由于空间具有自相关性,选择的公园至少间隔1km。
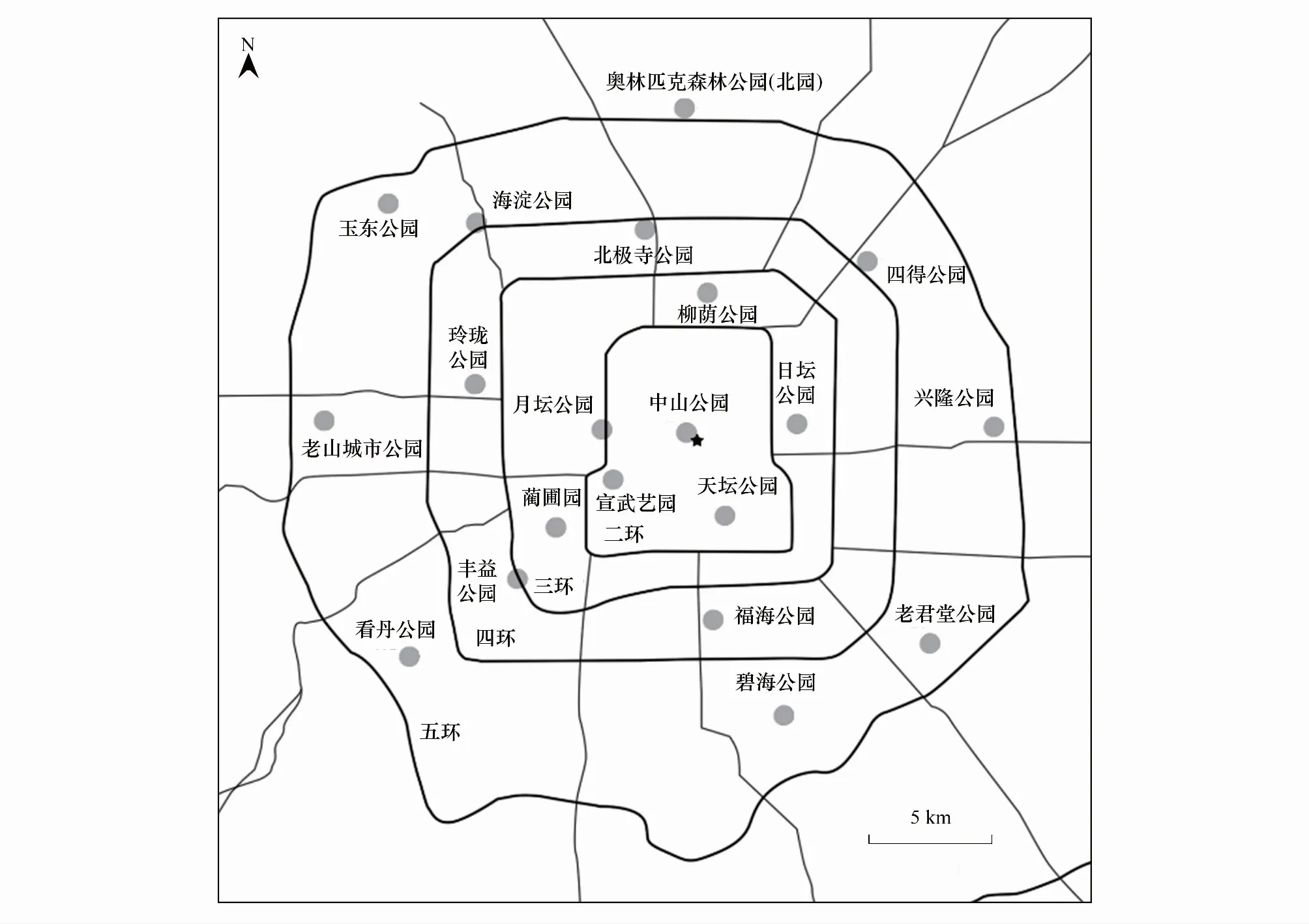
图 1 5环内20个城市公园位置Fig.1 Location of 20 urban parks inside 5th ring road灰色圆点为20个城市公园:AS:奥林匹克森林公园(北园);YD:玉东公园;HD:海淀公园;BJ:北极寺公园;SD:四得公园;LY:柳荫公园;XL:兴隆公园;RT:日坛公园;ZS:中山公园;YT:月坛公园;LL:玲珑公园;LS:老山城市公园;XW:宣武艺园;LP:蔺圃园;TT:天坛公园;FY:丰益公园;KD:看丹公园;FH:福海公园;LJ:老君堂公园;BH:碧海公园
1.2 声音数据采样方法
每处样点放置1台声音采集器(Zoom H5,Zoom Inc.,千代田区,东京,日本,System 2.40),于4月选择晴朗无风的4d进行24h连续监测(4月10日—4月14日),且所有样地测定均确保同步进行。
声音采集器(Zoom H5),采样率为44100Hz,采样位为16bit,立体声采样,音频格式为WAV。同时设置高通滤波器为220Hz,可消除部分电流杂音。Zoom H5是一款商用数字录音设备,带有两个内置单向电容麦克风,具有良好的声音可靠性,并已成功用于其他声景生态学研究[13, 15, 18, 21]。声音采集器应放置在群落中心位置,选择长势良好的乔木,固定在距地面2m位置。收音结束后,利用Python对声音样本进行系统随机抽样,每隔15min采集1min[22],共得到345600段1min音频文件。
1.3 样地植被调查
以采样点为中心,设置20m×20m植被调查样地。植被结构调查分为2个垂直层次:(1)所有高于1.5m木本植物种类、胸径(cm)、高度(m)、树冠高(m)等[21];(2)灌丛和地被层种类和数量。此外,测量并计算了样地内叶高多样性(Foliage Height Diversity, FHD)[23-24]。为了避免主观估计误差,群落植被调查全部由2名调查员共同完成。根据Wunderle的研究,叶高多样性(FHD)在每一个样点的东、南、西、北四个方向每隔4m确定一点测量,每个方向3个测量点,每个样点共12个测量点。在各测量点记录各高度水平空间(距离地面高度)是否有植物叶片,包括<0.5,0.6—1.0,1.1—2.0,2.1—4.0,4.1—6.0,6.1—8.0,8.1—10.0和>10m。最后,利用Shannon多样性指数计算得到FHD[25]。其他计算的植被垂直变量还包含树高变异指数(TCV)、乔木平均高度(TAH)、平均树冠高(CAH)。植被结构的水平方向变量包括:乔木层盖度(TC)、乔木层α多样性(TD)、灌木和地被层盖度(SGC)及α多样性(SGD)。
1.4 数据分析
1.4.1声景观分析
声景观中的不同实体如鸟类、两栖动物、哺乳动物、风和机械等产生不同频率的声音。生态系统中声音主要包括三种:生物声(Biopony)——有机体发出的生物声音,通常为中高频率(2—11kHz);人工声(Anyhrophony)——人类或人造物体产生的声音,通常频率较低(1—2kHz);地球物理声(Geophony)——自然过程(如风和雨)产生的声音,可能超越整个声谱(1—11 kHz)[3, 7, 26]。
声景观分析使用R统计计算环境。通过soundecology包中的“multiple sounds”功能,在2—11,025HZ区间内计算和可视化了共23040条声音样本。已排除0—1 kHz中的数据,其很大程度上来自风[27]。
(1)声学指数计算
本研究中使用了声景观研究中被广泛认可的3个声学指数:声学多样性指数(Acoustic Diversity Index,ADI)[13]、生物声学指数(Bioacoustic Index,BIO)[9]、归一化声景指数(Normalized Difference Soundscape Index,NDSI)。ADI将声谱划分为多个频带,并计算每个频带中超过- 50db的声音所占比例。BIO计算各频段曲线下与分贝值相关的面积,即与声级和鸟类所使用的频带数目相关的函数。NDSI通过计算人工声与生物声的比率衡量生境人为干扰程度[14],公式为:
其中生物声被定义为2—11 kHz的频率范围,人工声则被定义为属于1—2kHz范围。NDSI的低值表明人工声优势,高值则表示生物声优势。研究针对每个样点的声音数据进行整理,分别对各样点声学指数的日变化进行可视化。黎明合唱是鸟类歌唱活动的高峰,被定义为从日出前2h开始到此后2h结束的时期[18]。
(2)声强度计算
基于R自动计算的功率谱密度(Power Spectral Density, PSD)。所有的音频文件被划分为10个1 kHz频率区间后计算- 50dbfs以上声音的归一化功率谱密度。计算每个频率间隔的PSD值,可定量的描述其相对贡献,反映声事件的功率随时间和频率的变化[28]。
1.4.2样地植被类型划分
为了识别和解释仅城市公园(n=60)样地中声景观与植被的关系,创建了声学指数与植被因子的斯皮尔曼(Spearman)相关矩阵,并指导后续分析。
为了进一步解释样地植被对声景观影响,选择相关性分析中统计学显著的因子进行进一步分析:对样地的6个植被变量进行主成分分析(PCA)和最大方差法归一化旋转以方便解释。PCA将6个变量转换为三个主要成分(前三轴(Dim1—Dim3)解释了总方差的70%以上)。确定了三个轴中绝对负载值> 0.5的植被变量.
为了进一步划分植被结构类型,通过k-means聚类算法对Dim1—Dim3进行聚类分析。通过计算不同样本间植物信息的欧式距离来判断个案间的相近关系,依据解释变量方差的积累值的阈值(“手肘法”,Elbow Method)进行k值的选择。使用V值,即簇内距与簇间距的比值作为聚类结果的评价标准,通过比较不同k值对于V值的影响,选择最佳K值。据此,将采样点进行聚簇,并按聚类分布图统计出所有样地植被类别。使用Kruskal-Wallis(K-W检验)与多重比较,分析各样地类型中声景观的特征与差异。本节数据分析与可视化基于Python 3.9.0平台。
2 结果与分析
2.1 城市公园声景观日动态
2.1.1城市公园声学指数日变化特征
在声景观音频录制期间,绘制每天24h内NDSI、BIO、ADI指数的变化图(图2)。为方便比较,指数已进行归一化处理。总体来说,三种声学指数的日变化模式呈现相似的模式,揭示了与之相对应的生态过程[7, 29]。具体变化过程来看,NDSI和ADI变化趋势都呈现单峰型,BIO则呈现多峰分布。所有声学指数在黎明鸟类合唱时上升,在5:00—6:00达到最高峰,之后逐步下降,直到鸟类傍晚合唱时再次略微上升。其中ADI逐步下降,直到第二日清晨的黎明合唱前至最低值,此后迅速上升。NDSI则于傍晚7:00左右降至最低点,此后波动上升,在黎明合唱前出现短暂的小幅下降,此后继续上升至峰值。

图2 归一化声学指数(NDSI、BIO、ADI)的日变化 Fig.2 Daily variation normalized acoustic indices (NDSI, BIO, ADI)
所有声学指数都能准确反映出黎明合唱——在温带国家,这一春季最明显的鸟类保卫繁殖区域、试图吸引伴侣或召集同伴的生态活动。正如归一化植被指数(NDVI)评价植被健康一样,NDSI综合考虑生态系统生物声和人为干扰,因此被用来评价生态系统健康状况[18]。NDSI显示了北京城市公园内人类活动在傍晚20:00达到一天中的高峰,这反映了北京公园人类活动的实际情况,也符合其他关于城市公园公众行为研究[30]。夜间较高的NDSI反映了该指数对夜间生物活动敏感,例如直翅目昆虫等[31]。黎明前NDSI的小幅下降可能代表了从夜间声学信号向白天不同信号种类的过渡。综合来看,NDSI的日变化模式为声学生态位假说提供了支持[32]:两栖动物和昆虫倾向于在夜间发信号以规避鸟类及掠食者,因为声学生态位是动物争夺声音维度空间的有限资源[15]。
BIO在黎明合唱后小幅下降后仍保持较高数值,直到正午。这可能与人群在城市公园内的活动相关。这点验证了[33]在英国城市地区的研究结果。BIO基于频谱幅度的变化来计算的,反映了景观中声音的强度。BIO被证明与鸟类发声次数[34]和鸟类物种丰富度相关[9]。但当直接在城市地区应用BIO时,其结果可能被人声干扰。因此,本文在进行后期分析时,只采用未受到人声干扰的黎明期间数据。ADI测量声谱熵多样性[13],与鸟类多样性和物种活动有关[35]。BIO和ADI在16:00—19:00的小幅上升,反映了北京城市公园内鸟类群落的傍晚合唱,而二者较低的夜间值表明其对北京城市公园的夜间生物声不够敏感。
总体而言,北京城市公园生物声的日变化显著,由鸟类鸣声主导,具有明显的黎明合唱特征。
2.1.2城市公园声景观强度特征
将PSD在24h内的变化情况进行汇总,可视化沿频率区间的声强(图3)。最低频率功率值(1—2 kHz)在日出后波动上升,在晚上8时达到峰值,并在黎明前保持稳定,然后在日出前下降到最低点。与1—2 kHz相比,2—11 kHz(生物声)的PSD日变化呈现相反模式。2—3 kHz的声景强度峰值出现在凌晨5—6时,这段时间正好是日出时间。之后急剧下降,然后随着波动缓慢下降,直到第二天黎明。3—4 kHz除在傍晚有相对更明显的波动上升外,与2—3 kHz具有非常相似的模式。4—5 kHz则重点突出了鸟类的傍晚合唱。这可能反映了春季雌性到达繁殖地时,雄性开始减少黎明合唱,而在黄昏合唱期间具有更明显的歌声。该部分声音变化特征可能反映了雄性鸟类生殖期鸣声规律变化[36]。总体来看,2—5 kHz变化规律符合鸟类活动。5—11 kHz区间具有相似的模式:在夜间持续波动上升,直到黎明前(凌晨4时)达到最高值,在日出后急剧下降,然后在6—7时之间保持相对平稳。
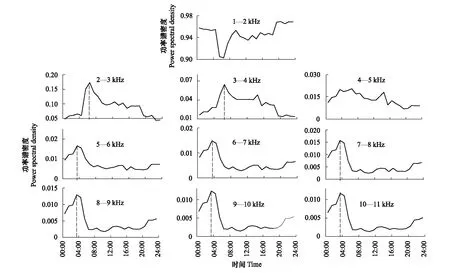
图3 共10个频率区间内声功率谱密度日变化Fig.3 Power Spectral Density daily variation of all 10 frequency intervals图中虚线标注了每个频率区间一天中的强度峰值及其所对应的时间节点
2.1.3城市公园声学生态位特征
基于ANH假设,声景观内不同频率的声强变化可能反映了不同声学群落的信息。因此,为了进一步揭示不同类型声音的变化,反映声音生态位,本研究可视化了1—2 kHz,2—5 kHz, 5—11 kHz频率区间在一天中占全部声景观的比例,以及其中声学生态位的变化(图4)。最低频率功率值(1—2 kHz)在一天中一直占据最大比重,最高达到90%以上,表明大部分时间人工声主导了城市公园声景观,主要包括车辆噪音、交谈声、广场舞等。此外,本研究发现声学生态位存在重合现象。在1—2 kHz发现了黑斑侧褶蛙(Pelophylaxnigromaculatus)和长耳鸮 (Asiootus)在夜间出现,但最低频区间的主要音源仍是人工声。
生物声中2—5 kHz的鸟类鸣声占据了主导地位。鸟类的听觉敏感频率在1—4 kHz[37],而城市鸟类最主要的鸣声频率在2.3和2.4 kHz之间[38],例如北京最常见的鸟类喜鹊(Picapica)和四声杜鹃(Cuculusmicropterus)。他们的合唱为黎明前后生物声在一天中的最高占比贡献了主要力量。而4—5 kHz的傍晚合唱由于此时也处于高强度的人工声,其在整体声景观中未能得到明显表现。5—11 kHz所贡献的绝对声音虽然能量非常低,但他们仍能共同反映了在夜间主导声景观的昆虫的鸣声规律[18],例如声音最响亮的蟋蟀(Grylloidea)、螽斯(Tettigoniidae)等。昆虫发出的声音比鸟类的声音频率高,也出现不同的峰值时间段,说明在城市背景下各自占有不同的声学生态位(图4)。

图4 北京城市公园声强(watts/kHz)相对比例及其生态位日变化图Fig.4 Percent of total soundscape power (watts/kHz) and acoustic niches variation for frequency intervals
2.2 样地植被特征对声景观的影响
样地植被特征因子与声学指标的相关性分析证实了植被与声景特征之间的显著关系(表1)。在植被结构复杂的样地中,NDSI较高,这些植物群落具有复杂的分层结构,无论是乔木层或灌草层,都具有较高的覆盖度。BIO则在具有高大乔木以及更高乔木树冠的样地中更高。相似地,在具有高大乔木的植被类型中,声学多样性(ADI)也更高。这反映了城市生境中大树对声学群落的具有重要的正面作用。大树能够提供更多的结构复杂性[39-40],包括较大的树冠,枯死的树枝和空洞,这些小树所不能具有的结构能够为无脊椎动物、爬行动物、鸟类和哺乳动物提供重要的食物和生境资源[41]。但灌草层的植物多样性则对ADI具有一定负面影响。这可能是因为有较多灌草多样性的地块往往种植草坪,具有很高的人为管护强度,包括随季节更替地被,高频率的修剪、除草、农药的使用。这些人为干扰影响城市野生动物活动,尤其是地面筑巢物种,因此其声学多样性也相应地减少。
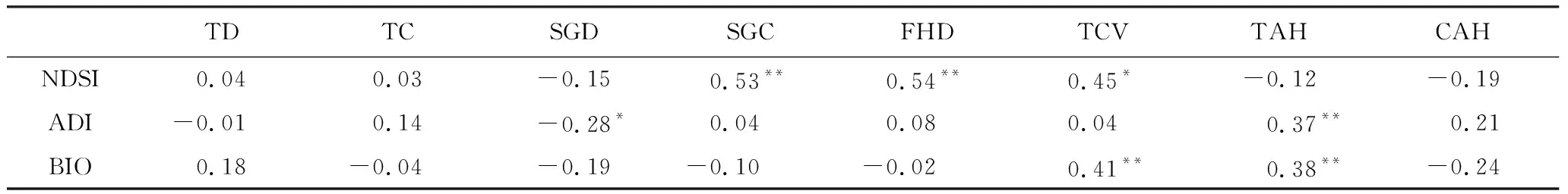
表1 声学指数和样地植被变量的斯皮尔曼秩相关矩阵(n=60)Table 1 Spearman rho correlation of habitat factors with acoustic indices (n=60)
2.3 植被与声景观的关系
2.3.1植被类型组成
根据相关分析结果,选择对声学指数具有影响的六个样地植被变量进行进一步分析(虽然变量TC也在统计学上显著,但由于其相关系数过小,因此予以剔除)。PCA将六个变量转换为三个主要成分(其解释了总方差的70%以上)。确定了三个PC中绝对负载值> 0.5的植被变量(表2)。

表2 样地植被变量的主成分分析结果Table 2 Principal component analysis of the variables that responded to the vegetation structure
变量TAH和CAH控制第一轴(Dim 1,绝对载荷>0.6,表1),并且它们的组合得到相关性的支持(r=0.73,P<0.001)。因此,Dim 1主要区分了样地植被中乔木层高度。变量SGC、FHD和TCV区分了第二轴(Dim 2,绝对载荷>0.5)。TCV主要定义了样地中乔木层的垂直复杂度,SGC衡量灌木和地被层,FHD则综合衡量了样地乔灌草层的垂直结构。因此本研究确定了Dim 2主要与样地的垂直复杂度相关。Dim 3反应了灌草层多样性信息。
进一步,依据PCA结果,通过 k-means算法对植物群落进行划分。聚类最佳k值为3,即60个样地被划分为3种具有植被结构特点的植被类型,样地聚簇结果如图5。划分的4种样地植被类型分别为:灌草层发达型(SG型,n=30)、垂直结构发达型(V型,n=22)、高大乔木型(T型,n=8)。
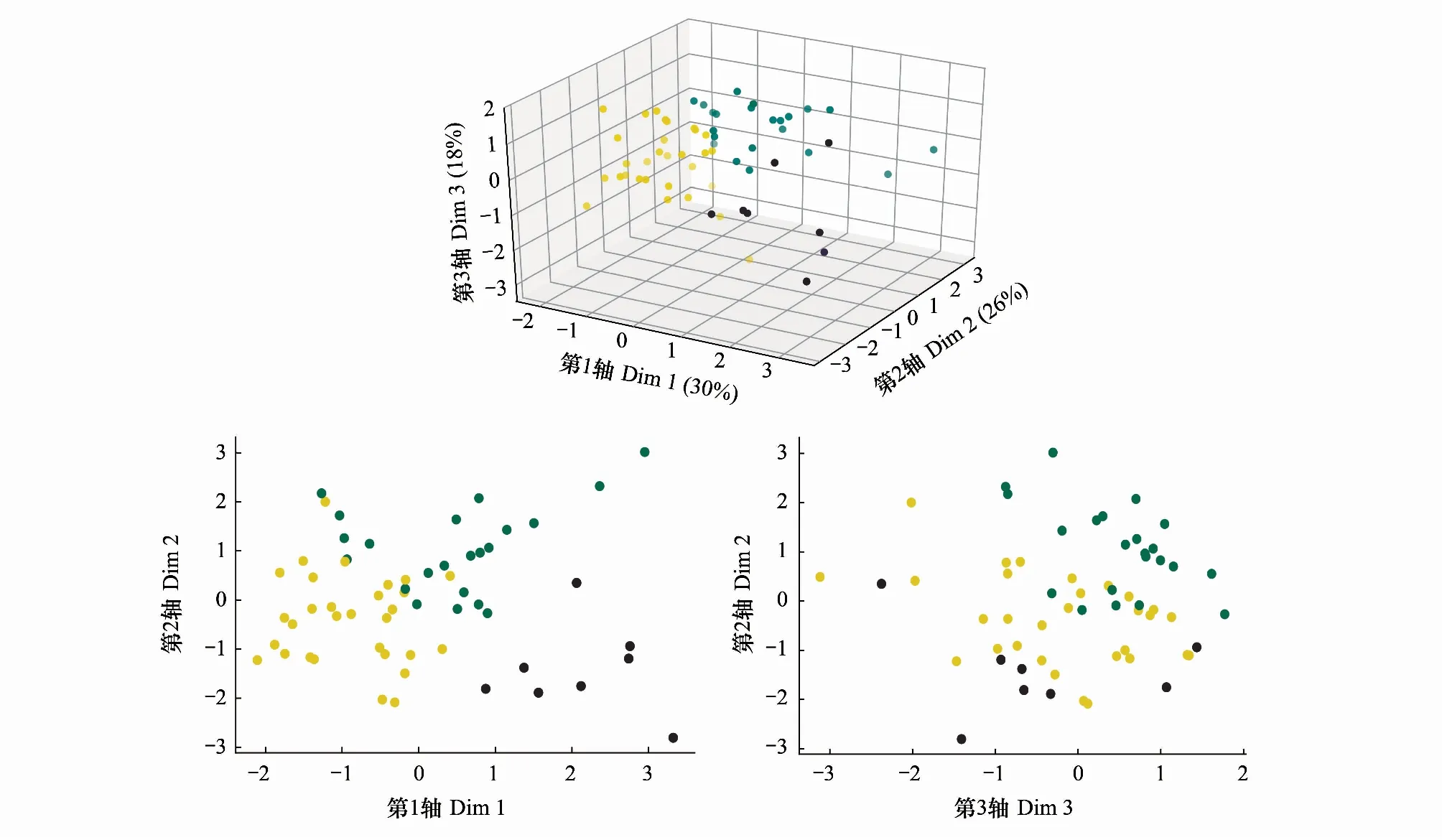
图 5 样地在PCA排序轴(Dim1—3)上的2维和3维展开图Fig.5 The 2-D and 3-D distribution of sampling sites in PCA图中三种颜色代表聚类分析结果:黄色为SG型、绿色为V型、紫色为T型
2.3.2不同样地植被类型中声景观特征
所有样地植被类型中,黎明时段的声学指数都显著高于全日值(图6),反映了鸟类黎明合唱(dawn chorus)充满活力,声音更大,频率更高的特点。对声学指数进行非参数独立样本克鲁斯卡尔.沃利斯(K-W)检验和多重比较结果表明,在95%置信区间内,BIO和NDSI在三种样地植被类型中具有显著差异(BIO:χ2=14.8,P<0.001; ADI:χ2=20.7,P<0.001)。NDSI在黎明合唱期间数值受到样地植被垂直结构变化的最大影响。NDSI在具有发达植被垂直结构的V型样地植被类型中最高,而在具有高大乔木,疏林草地的T型样地植被类型中最低。与NDSI不同,BIO则在T型中最高,在灌草层发达但乔木层低矮的SG型中最低。ADI则在不同植被类型间未发现显著差异。结果表明不同结构的植被,可能导致不同的生物群落,同时形成种类丰富的声景观。

图6 声学指数在不同样地植被类型间的差异Fig.6 Differences of acoustic indices in different vegetation types
3 讨论
本研究量化(图2、3)并可视化(图3)了北京二十个城市公园内60个样地的声景观特征与动态,可以将其视为北京城市公园声景观研究的基线记录。随着声音数据被量化为声景观模式并通过这些声指数的可视化,可以更好地总结出一种有效的方法,以跟踪和评估随时间和空间变化生态系统的健康状况,并更好地理解声景观在生态过程中的作用[7]。
本研究对1—11kHz所有频率段在24h的变化模式的可视化能够反应包括鸟类、昆虫等声学群落的活动与相关的生态过程,其中几乎所有频率段都强调了非常清晰的黎明合唱,其具有一天中最高的功率,这与以前的研究非常一致[15, 17, 21, 42]。此外,每个频率段不同的模式为声学生态位假说提供了支持。这是声学群落分配声音资源的结果[12]。研究表明,动物可以调节其呼叫频率以适应嘈杂和复杂的声学环境,从而减少噪声干扰。
长期以来,植被结构在鸟类组合的决定中起着重要作用[43]。大多数关于声景观的研究都在自然保护区或森林中进行研究,这是首个探讨北京城市地区声景观与植被关系的定量研究。研究结果证明了在城市环境中,植被结构同样对声景观具有重要作用,这与前人的发现一致[15, 28, 44]。此外,相比于水平特征,植被垂直结构对声景观具有更明显的影响。当垂直异质性越强,冠层高度不均匀或植被多层时,生物声强度以及声学多样性也越强。除了为鸟类和其他野生动物提供更丰富的栖息地和食物资源,公园中复杂的垂直植物空间也可能因为限制公众进入,为城市野生动物提供躲避人类活动的避难所。NDSI是衡量生物声景观和人类干扰强度的综合指标,其在V型植被类型中的显著升高可以证明这一点。在更开放的植被环境中较低NDSI,也反映了人们对公园中散布着高大树木的开放景观的偏好[45]。大树是城市公园的关键结构[39]。随着大树数量的增加,它们对声学群落的积极影响也增加了。Chitra等人提出与其他植被结构相比,乔木是影响公园整体声景观的最重要变量[46]。大树能为无脊椎动物和脊椎动物提供更更富的食物和微生境资源,比如更多的花朵、花粉,花蜜,种子等[39, 40]。根据统计结果,高大树木T型植被类型在所有公园中最少,反映了北京城市公园中大树的损失。而城市环境中大树的损失可能产生深远的生态后果,是生态可持续性的巨大威胁[47]。
因此,在城市公园建设中,保留原生植被群落以及“近自然”的植物规划理念是可取的。然而,当前大部分公园的植物规划仍以“审美”为主要目的。本研究表明,设计公园时,植物的选择以及丰富的植被群落结构对于维持城市野生动物群落至关重要。由于乔木是最重要的植被结构之一[39, 48],因此其树种、规格的选择,以及食源性树种的合理种植至关重要。
北京的深度绿化工程,例如“百万亩平原造林”为候鸟提供了更多的栖息地,但是如果没有合理的城市森林管理策略,城市生物多样性的收益预期则会大大降低[49]。因此,除了不断推进绿化工程,本文建议重视城市森林的有效管理。
高强度的管护和随季节更替地被植物在城市公园中很常见,以这种方式创造的优美景观确实可以吸引游客并达到增加绿色娱乐价值的效果[50]。此外,北京城市公园常使用农药控制昆虫爆发,这将严重影响城市野生动物,尤其是鸟类的繁殖[51]。地被层也是鸟类食物来源地之一[52],建议在保留供游客休憩草坪区域的基础上,在公园的部份区域减少人为管护强度,允许地被长高、缓扫落叶、有机地表覆盖的使用[53],将有利于例如,地面巢鸟类如普通燕鸥,或地面捕食鸟类,如家麻雀(Passerdomesticus)等鸟类筑巢与觅食[54]。此外,这些措施也可以一定程度上改善除鸟类外的其他生物,例如蝴蝶或昆虫的生境质量[49]。这些举措具有促进城市生物多样性的潜力,但是需要系统的监测和评估,以确保实现积极的效果。本研究为利用声学指数作为植被状况对生物多样性价值的替代方法提供了证据,显示了声景观方法作为评估城市生态系统健康状况的快速工具的潜力。
4 结论
对城市绿化的监控不足,无法满足其对生物多样性供应和生态系统支持的关键作用。在北京没有声景观历史数据的情况下,本文首次对城市公园声景观数据进行系统地收集与统计,分析了春季声景观的时间动态变化与跨频率区间变化特征,结果反映了不同声学群落的存在与活动,验证了声学生态位假说。本研究结果可以基准,进一步与物种数据结合。相关和主成分分析表明,不同结构的植被,导致不同的生物群落,同时形成种类丰富的声景观;其中,植物群落的垂直结构对声景观质量和多样性具有更重要作用。这种关系可以用于景观规划以及衡量管理策略对生物多样性的进展和影响。本研究在发展低成本城市生态系统资源评估,和低侵入性生物多样性管理方面具有重要的潜在应用价值,能够指导相关战略规划和决策制定。
猜你喜欢
现代园艺(2021年23期)2021-12-01 07:47:44
疯狂英语·初中天地(2021年6期)2021-08-06 09:03:46
林业勘查设计(2020年1期)2021-01-18 02:40:48
新农业(2020年18期)2021-01-07 02:17:08
家庭影院技术(2020年6期)2020-07-27 01:37:54
家庭影院技术(2019年1期)2019-01-21 02:25:04
家庭影院技术(2018年11期)2019-01-21 02:20:50
家庭影院技术(2018年10期)2018-11-02 05:35:26
现代园艺(2017年22期)2018-01-19 05:07:04
现代园艺(2017年23期)2018-01-18 06:58:07
