
橡胶树无性系光合特性及叶片解剖结构比较
2021-11-10 07:22邱彦芬亚华金李小琴张凤良
热带农业科技 2021年4期
邱彦芬,亚华金,杨 湉,李小琴,张凤良,李 玲,吴 裕
(云南省热带作物科学研究所,云南景洪 666100)
橡胶树(Hevea brasiliensis)原产于巴西亚马逊河流域,属于典型的热带雨林树种,也是目前种植规模最大的产胶植物[1]。光合作用作为植物同化物的代谢途径,是植物生长发育的基础和决定植物产量的重要生理过程,而叶片作为光合作用的主要器官,叶片的结构特征能体现环境对植物的影响和本身对环境的适应,具体表现为解剖结构、气孔结构的差异[2]。结构是功能的基础,叶片结构的改变必然会引起生理、生态功能的变化[3-4]。已有研究表明,种内的光合速率差异是由测定方法和叶片解剖结构的先决影响造成的[5]。栅栏组织的厚度在一定程度上决定叶片的光合速率,爵床科植物中的银爵,栅栏组织最小,但是净光合速率最大,表明栅栏组织厚度与净光合速率具有一定相关性[6-7]。小麦旗的叶片厚度与其蒸腾速率、净光合速率呈显著的正相关[8]。绿化树种光饱和点与叶表皮厚度显著正相关。而胞间CO2浓度与上表皮的气孔密度显著负相关[9]。大豆的叶片厚度和单位面积栅栏组织细胞数决定了大豆叶片的光合速率[10]。目前,针对橡胶树无性系间叶片结构与光合特性关系的研究可查阅的文献较少。鉴于此,本文选取7个橡胶树优良无性系测定其叶片解剖结构指标和光合生理指标,旨在比较分析不同无性系间叶片解剖结构变化与其光合生理特征的关系。
1 材料和方法
1.1 试验地自然概况
试验地设在云南省热带作物科学研究所管辖的“农业部景洪橡胶树种质资源圃”内,该资源圃位于西双版纳傣族自治州景洪市城区。东经100°46’~100°48’,北纬21°59’~22°01’,海拔500~600 m;区域内地形复杂,中山、低山、丘陵组合,具有立体气候特征,是云南省农业生产环境最为特殊的地区之一。区域内全年日照数达1 800~2 200 h,年降雨量为900~1 500 mm,年平均相对湿度71.3 %~85 %;6-10 月为雨季,占全年降雨量的80 %;11-2 月为冬季,低温干燥;3-5 月为旱季,高温干旱;干湿季节分明,年温差小于昼夜温差。
1.2 试验材料
选择2017 年定植的7 个优树无性系(plustree clones)为研究对象(表1),每个无性系选择3株,每株取3 片无病虫害、生长一致的叶片,取样部位均为植株中上部稳定叶蓬。
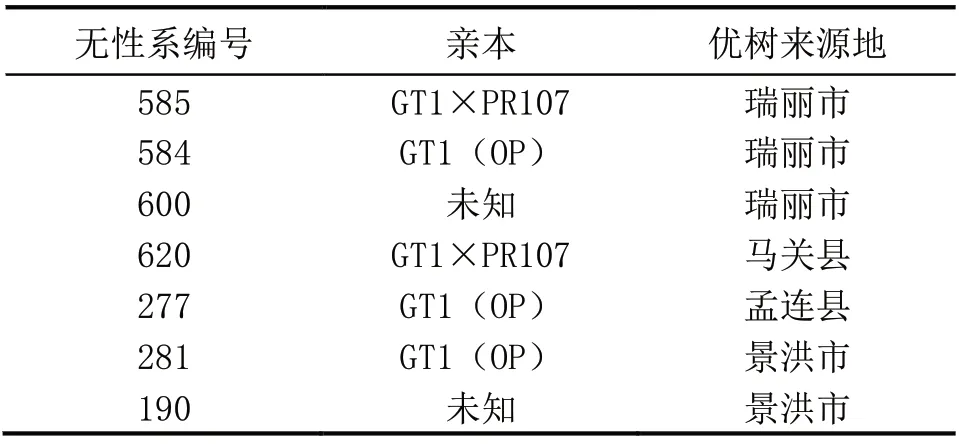
表1 橡胶树无性系基本信息
1.3 光合指标的测定
选择2020 年8 月生长旺季,在天气晴朗的早上(9:00—11:30)进行光合测定,使用Li-6400 便捷式光合测定仪(Li-COR 公司,美国)。活体测定不同无性系的净光合速率(Pn)、胞间CO2浓度(Ci)、气孔导度(Gs)和蒸腾速率(Tr)。为避免自然光强的不同和因环境因素导致的数据误差,人工控制将光合有效辐射设定为1 200 μmol·m-2·s-1,样品室流速500 μmol/s,叶室风扇为Fast 模式,控制混合器关闭。
1.4 叶片解剖结构的测定
光合指标测定完成之后,采集叶片带回实验室,剪取叶片中部,避开主脉及边缘,大小为5 mm×5 mm,然后立即放入50% FAA 固定液中于4℃冰箱固定72 h 以上,采用石蜡切片法[11]做成永久装片,在LEICA DM6 B 显微镜下拍照观察、测量数据。测量指标为上角质层厚度、上下表皮厚度、栅栏组织厚度、海绵组织厚度。
1.5 统计分析
采用Microsoft Excel 2007 对数据进行整理,SPSS 20.0 对无性系间解剖结构指标与生理指标作单因素方差分析,对方差分析差异具有统计学意义的指标再进行多重比较。
2 结果与分析
2.1 无性系叶片显微结构观察
橡胶树叶片的横切面结构由表皮、叶肉和叶脉3 部分组成,上表皮下有占比较大的栅栏组织,栅栏组织下面为细胞分散疏散的海绵组织,为典型的异面叶。叶片的表皮由一层细胞构成,细胞较大,呈椭圆形或矩形,排列比较整齐(图1);表皮细胞外覆盖着厚度不同的角质层(图A-G)。
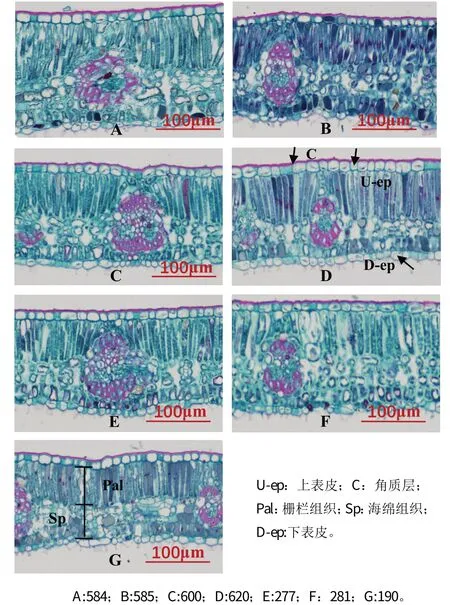
图1 橡胶树无性系的叶片解剖结构(标尺=100μm)
橡胶树叶片的栅栏组织紧贴上表皮,具有整齐清晰的层次,排列紧密而有序,在叶片的横切面中,栅栏组织占据了较大比例,由2~3 层细胞组成,叶绿体主要分布在栅栏组织中;海绵组织细胞排列疏松,且细胞形状不规则、大小不一,细胞之间有较大间隙;下表皮细胞横切面形状不规则,厚度差异较大。
2.2 无性系叶片解剖结构差异
比较7 个无性系的叶片解剖数据可以发现(表2),无性系角质层厚度、上下表皮厚度、栅栏组织厚度、海绵组织厚度、叶厚差异显著(P<0.05,下同)。7个无性系叶片的角质层厚度变异系数为28.35%,无性系584 角质层最厚,为4.67 μm,277、281 最薄,分别为2.11、2.23 μm。 上表皮厚度变异系数为13.28%,无性系277 最厚,为11.39 μm,无性系281 最薄,为9.00 μm。栅栏组织厚度变异系数为20.96%,无性系584最厚,为87.23 μm,无性系281 最薄,为48.66 μm。海绵组织厚度的变异系数为20.19%,无性系584 最厚,为67.89 μm,无性系281 最薄,为41.78 μm。下表皮厚度变异系数为17.16%,无性系584 最厚,为10.02 μm,无性系190 最薄,为8.14 μm。叶片厚度的变异系数为16.94%,无性系584 最厚,为176.41 μm,无性系281 最薄,为107.93 μm,无性系584、281 与其它无性系差异显著。
2.3 无性系光合生理参数的差异
从表3 可以看出,7 个无性系间叶片的净光合速率、气孔导度、胞间CO2浓度、蒸腾速率、水分利用效率差异显著(P<0.05,下同)。7 个无性系的净光合速率变异系数为23.17%,无性系620 净光合速率最大,为14.22 μmol·m-2·s-1,与其它无性系差异显著,无性系281 净光合速率最小,为8.50 μmol·m-2·s-1,与190差异不显著,与其它无性系差异显著。气孔导度变异系数为39.18%,无性系620、277 气孔导度最大,分别为0.25 、0.22 mol·m-2·s-1,与其它无性系差异显著,584 最小,为0.09 mol·m-2·s-1。胞间CO2浓度变异系数为18.09%,无性系277、281 胞间CO2浓度最大,分别为290.57 、283.38 μmol·mol-1,与无性系584、620、190 差异显著,无性系584 胞间CO2浓度最小,为177.86 μmol·mol-1,与其它无性系差异显著。蒸腾速率变异系数为38.98%,无性系620 最大,为3.24 mmol·m-2·s-1,无性系600 最小,为1.31 mmol·m-2·s-1,620、600 与其它无性系差异显著。水分利用效率变异系数为32.00%,无性系600 最大,为7.82 μmol·mmol-1,与其它无性系差异显著,无性系620、281 最小,分别为4.19、4.79 μmol·mmol-1。
2.4 叶片解剖结构与光合生理参数的相关性分析
通过表4 可以看出,7 个无性系的角质层厚度与胞间CO2浓度(Ci)呈极显著(P<0.01,下同)负相关,相关系数为-0.913。叶片厚度与胞间CO2浓度(Ci)呈显著(P<0.05,下同)负相关,相关系数为-0.755。胞间CO2浓度(Ci)与上表皮厚度、栅栏组织厚度、海绵组织厚度、下表皮厚度相关性均没有达到显著水平(P>0.05,下同)。净光合速率、气孔导度、蒸腾速率、水分利用效率,与角质层厚度、上下表皮厚度、栅栏组织厚度、海绵组织厚度、叶厚相关性均未达到显著水平。

表4 无性系叶片解剖结构与光合生理参数的相关性分析
3 讨论与结论
橡胶树虽然是热带雨林中的成员,但是也具有一定程度的旱生结构特征:不同无性系间叶片解剖结构存在相似性,具有较为发达的角质层,叶片上下表皮的形态基本一致(呈近长方形或椭圆形),上下表皮细胞排列紧密、整齐清晰,且有占比较大的栅栏组织。
7 个橡胶树无性系间叶片的解剖结构指标(角质层厚度、上表皮厚度、栅栏组织厚度、海绵组织厚度、下表皮厚度、叶片厚度)、光合生理指标(净光合速率、气孔导度、胞间CO2浓度、蒸腾速率、水分利用效率)均存在显著差异。对7 个无性系间叶片解剖结构指标与光合参数相关性分析显示,无性系的角质层厚度与胞间CO2浓度(Ci)呈极显著负相关。已有研究表明,较厚的角质层可以保住植物水分,减少水分向大气蒸发,能适应和抵御外界环境的胁迫,维持植物正常的生理机能[12]。因此,较厚的角质层增强了对太阳的反射,间接减少了对太阳能的吸收利用,细胞间的CO2浓度量减少,可能降低了胞间的CO2浓度。赵雪[13]、庞杰[14]研究表明,叶片厚度与气孔导度及胞间CO2浓度呈极显著正相关。本研究中,叶片厚度与胞间CO2浓度(Ci)呈显著负相关,与赵雪、庞杰等研究结果不一致,可能是树种之间存在一定的差异,还需要进一步研究论断。
本研究中,光合生理指标与栅栏组织厚度、海绵组织厚度、上下表皮厚度均无相关关系。但有研究表明,文冠果净光合速率与上表皮厚度及海绵组织厚度呈极显著正相关[13];油橄榄栅栏组织厚度、栅海比与净光合速率极显著正相关。这些指标在一定时期和范围内可判断植物光合能力[15]。光合特性存在基因型及种的差异,但空气中CO2、湿度,叶片类型、叶龄也对其有影响[16]。
本研究结果表明,在无性系之间,叶片的解剖结构影响其生理活动,对叶片结构的研究可以更好地理解光合速率、呼吸速率和蒸腾速率的变化。值得注意的是,本研究选择的是处于稳定期的橡胶树无性系叶片,对于其各个发育时期的叶片结构、生理指标会如何变化,差异会大还是小则还不清楚,还需进一步做研究。
猜你喜欢
热带作物学报(2022年8期)2022-09-16
农业工程学报(2022年4期)2022-04-24
作文周刊·小学二年级版(2021年40期)2021-01-05
农业与技术(2020年12期)2020-07-04
伴侣(2020年4期)2020-04-27
伴侣(2020年2期)2020-04-13
农民致富之友(2019年29期)2019-10-21
散文选刊·下半月(2018年2期)2018-02-23
新课程·下旬(2017年7期)2017-08-14
健康必读(2016年10期)2016-11-14
