
矮化中间砧长度对红富士苹果苗木和幼树生长影响及生理机制研究
2021-11-10 13:11尹宝颖付晓雅朱玉宁梁博文李中勇徐继忠
河北农业大学学报 2021年5期
沈 震,尹宝颖,付晓雅,朱玉宁,梁博文,李中勇,徐继忠
(河北农业大学 园艺学院,河北 保定 071001)
苹果矮砧密植栽培具有矮化、早花早果、丰产优质、便于集约化栽培管理等优点,现已成为国内外现代苹果生产发展的主流和方向[1]。应用矮化中间砧是苹果矮砧密植栽培的一种方式。据研究显示,不同矮化中间砧[2]和同一矮化中间砧不同嫁接长度[3]对苹果树体生长结果均会产生较大的影响,且由于中间砧长度不一,致使建园后树体高矮有差异,严重影响了整齐度,植株生长发育不一致,影响了经济效益[4]。
郭金丽、Karlidag和高艺翔等[4-6]报道了不同中间砧长度对金红、金冠、澳洲青苹及长富2号树体生长的影响,结果表明随中间砧长度的增加,矮化效果增强。但杨廷帧、程万强等[7-8]对红富士、阳丰甜柿树体生长的研究则认为,随着中间砧长度的增加,嫁接苗的矮化性状均呈现先升高后降低的变化趋势。王海波、Marcon Filho、杜俊兰等[9-11]研究表明不同中间砧长度对泰山嘎拉、皇家嘎拉、长富2号果实品质及产量的影响差异显著。前人关于不同矮化中间砧长度的研究主要集中在树体生长结果上,而不同矮化中间砧长度对树体生长影响的生理机制,特别是对苹果树体酶活性及内源激素的影响尚缺乏系统的研究。
‘冀砧2号’是河北农业大学选育的苹果矮化砧木,2016年通过河北省林木品种审定,作为中间砧嫁接‘天红2号’矮化性状明显,嫁接亲和性好,性状稳定[12-13],目前关于冀砧2号中间砧长度方面的研究还未见报道。因此,本研究以红富士苹果幼树(天红2号/冀砧2号/八棱海棠)和苗木(天红2 号/冀砧2号/平邑甜茶)为试材,通过对不同矮化中间砧长度苹果幼树、苗木的生长以及苗木叶片内生理指标进行分析,旨在明确中间砧长度的作用及生理机制,以期为推广矮化中间砧集约化栽培模式提供理论和实践依据。
1 材料与方法
1.1 试验材料
不同中间砧长度对幼树生长的影响调查在河北省顺平县南神南苹果试验基地进行。砧穗组合为‘天红2号/冀砧2号/八棱海棠’,2017年定植,株行距为1.5 m×4 m,中间砧长度为(10±2)、(30±2)、(50±2) cm,标记为T-10、T-30、T-50。试验采用随机区组设计,每处理5株树,单株小区。2019、2020年调查树体生长指标。试验树进行常规管理。
不同中间砧长度对苗木生长的影响调查在河北农业大学农科教学实习基地进行,以‘天红2号/冀砧2号/平邑甜茶’半成品苗为试材,栽植于装有育苗基质的塑料花盆(盆口直径30 cm,盆底直径20 cm,盆高22 cm)。中间砧长度设计为(10±2)、(30±2)、(50±2)cm,标记为P-10、P-30、P-50。随机区组设计,每处理4株为1小区,重复4次。选取生长势基本一致、无病虫害的树体用于试验,进行常规管理。
1.2 试验方法
1.2.1 生长参数调查 苹果幼树分别于2019年10月、2020年10月植株停长时,利用卷尺测量幼树中间砧上接口至中心干延长梢顶端的长度、干周、外围新梢长度。苗木于2020年10月植株停长时,利用卷尺测量新梢长度,利用游标卡尺测量中间砧干径、品种干径,新梢粗长比为品种干径和新梢长度的比值。
1.2.2 取样 于2020年7月中旬至9月中旬进行采样测定苗木酶活性;于2020年9月中旬进行采样测定苗木激素。每处理4株为1小区,重复4次。用干净的纱布擦去叶片表面的灰尘,立即置于液氮罐内带回实验室,置于-78 ℃超低温冰箱中待测。
1.2.3 酶活性的测定 吲哚乙酸氧化酶(IOD)活性测定采用2, 4-二氯酚法,过氧化物酶(POD)活性测定采用愈创木酚法[13]。
1.2.4 同位素15N分配量、利用率的测定 2020年7月中旬每盆苗木施用1 g15N-尿素,施肥后立即浇水。10月植株停长时,将植物解析为根系、基砧(茎)、中间砧、中心干、叶片5部分,烘干至恒重后用磨样机磨成粉末,混匀后装袋备用。15N同位素通过元素分析仪结合稳定同位素比率质谱仪测定,各部位氮素分配量为各部位的干质量与元素质量分数的乘积[14-15]。氮肥利用率为植株15N总量与总施肥量的比值[16]。
1.2.5 叶片激素含量的测定 采用高效液相色谱法,参考卢颖林[17]的方法从叶片中提取内源激素(IAA、ZR、KT和GA3含量)。
1.3 数据分析
所得数据用 SPSS 20.0数据分析软件进行统计分析,采用Microsoft Excel软件制图表。
2 结果与分析
2.1 矮化中间砧长度对红富士苹果幼树和苗木生长的影响
2.1.1 对幼树生长的影响 2019、2020年2年试验结果表明(表1),在本试验设置的中间砧长度范围内,随着中间砧长度的增加,幼树中心干长度、中间砧干周、品种干周、新梢长度均呈递减趋势。中间砧长度为10 cm的中心干长度(381.75 cm)显著高于T-30和T-50;T-10处理的中间砧干周为22.50 cm,高于T-30,显著高于T-50;T-10处理的品种干周为24.63 cm,高于T-30、T-50;T-10处理的新梢长度(26.80 cm)显著高于T-30和T-50。表明随矮化中间砧长度增加,树体矮化效应逐渐增强。

表1 矮化中间砧长度对幼树生长的影响Table 1 Effect of dwarfing interstock length on growth of young trees
2.1.2 对苗木生长的影响 由表2可知,在本试验设置的中间砧长度范围内,随着中间砧长度的增加,中间砧干径、品种干径及新梢长度均呈递减趋势。P-10处理的中间砧干径为15.29 mm,显著高于P-30和P-50;P-10处理的品种干径(8.00 mm)显著高于P-30、P-50,P-30与P-50间则没有显著差异;P-10处理的新梢长度(73.67 cm)显著高于P-30和P-50(57.33和45.5 cm);P-30处理的新梢粗长比最大,为0.014,显著高于P-10、P-50。表明随矮化中间砧长度增加,树体矮化效应逐渐增强。
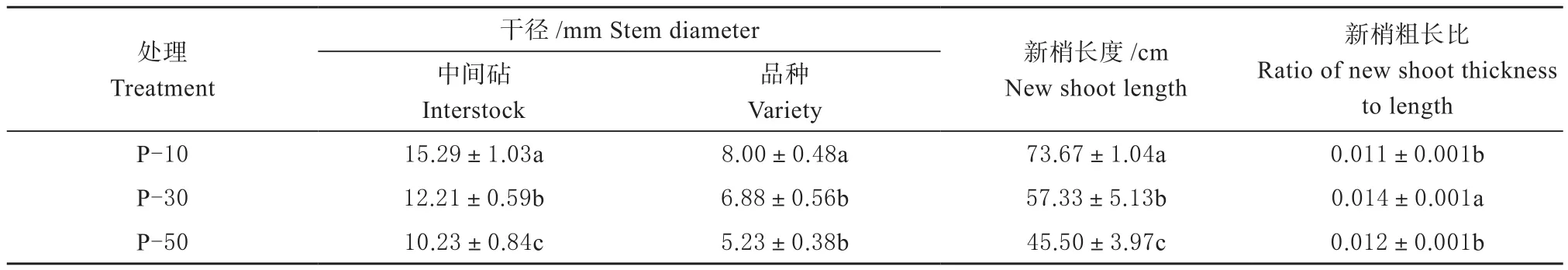
表2 矮化中间砧长度对盆栽树体生长的影响Table 2 Effect of dwarfing interstock length on the growth of potted trees
2.2 矮化中间砧长度对苗木同位素15N分配量、利用率的影响
2.2.1 对叶片15N浓度的影响 由图1可知, 不同中间砧长度对叶片15N浓度有显著影响,叶片15N浓度由大到小依次为P-10>P-30>P-50,P-10处理的叶片15N浓度为0.12 mg/g DW,高于P-30(0.11 mg/g DW),显著高于 P-50(0.10 mg/g DW)。
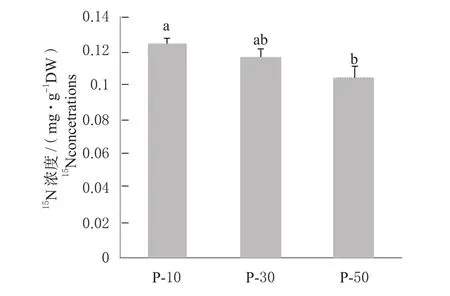
图1 矮化中间砧长度对苹果苗木叶片15N浓度的影响Fig. 1 Effect of dwarfing interstock length on 15N concentrations in apple seedlings leaves
2.2.2 对分配量的影响 由图2可知,不同中间砧长度对全株氮素分配量有明显影响,全株15N分配量由大到小的排序为P-10>P-30>P-50,P-10处理的全株分配量为6.62 mg DW,显著高于P-30、P-50。不同中间砧长度影响了苹果幼树各器官对氮素的吸收,各器官氮素分配量表现为根>叶片>中间砧>中心干>基砧(茎)。P-10处理的根内15N分配量为3.35 mg DW,显著高于 P-50,高于 P-30。P-10 处理的叶片、中心干内15N分配量显著高于P-30、P-50;P-50处理的中间砧内15N分配量(0.85 mg DW)是P-10的1.47倍,显著高于P-30;各处理基砧(茎)内15N分配量没有显著差异。

图2 矮化中间砧长度对苹果苗木各器官15N分配量的影响Fig. 2 Effect of dwarfing interstock length on the assigned amount of 15N in each organ of apple seedlings
2.2.3 对利用率的影响 如图3所示,不同中间砧长度对苹果苗木同位素15N利用率有显著影响,15N利用率由大到小依次为P-30> P-50> P-10,P-30处理的的15N利用率为6.62 %,显著高于P-10、P-50,P-10和P-50间15N利用率无显著差异。

图3 矮化中间砧长度对苹果苗木同位素15N利用率的影响Fig. 3 Effect of dwarfing interstock length on 15N availability of apple seedlings
2.3 不同矮化中间砧长度对苗木叶片酶活性的影响
2.3.1 对叶片内IOD酶活性的影响 由图4可知,3个时期不同中间砧长度苗木的叶片内IOD酶活性由大到小依次为P-50>P-30>P-10,与树体中间砧干径、品种干径、新梢长度长势排序相反。7月15日,P-5处理的叶片内 IOD 酶活性为 17.51 μmol/(h·g)显著高于其他2组处理;8月15日,P-50处理的叶片内IOD酶活性最高,为15.31 μmol/(h·g),显著高于P-10,高于P-30;9月15日,P-50处理的叶片内IOD酶活性最高,为11.63 μmol/(h·g),显著高于P-10,高于P-30。这表明,随矮化中间砧长度增加,叶片内IOD酶活性升高,树体矮化效应逐渐增强。
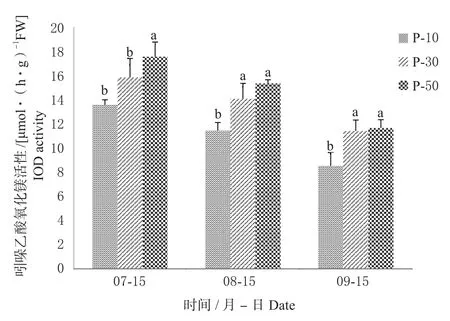
图4 矮化中间砧长度对叶片内IOD酶活性的影响Fig. 4 Effect of dwarfing interstock length on IOD activity in leaves
2.3.2 对叶片内POD酶活性的影响 由图5可知,3个时期不同中间砧长度苗木叶片内POD酶活性由大到小依次为P-50>P-30>P-10,与树体中间砧干径、品种干径、新梢长度长势排序相反。7月15日,3个处理的叶片内POD活性差异显著,P-50处理的叶片内POD活性为1 860.00 U/(mim·mg),显著高于P-10、P-30;8月15日,P-50处理的叶片内 POD 活性为 1 240.00 U/(mim·mg),显著高于P-10、P-30;9月15日,P-50处理的叶片内POD 活性最高,为 1 093.33 U/(mim·mg),显著高于P-10、P-30。这表明,随矮化中间砧长度增加,叶片内POD酶活性升高,树体矮化效应逐渐增强。
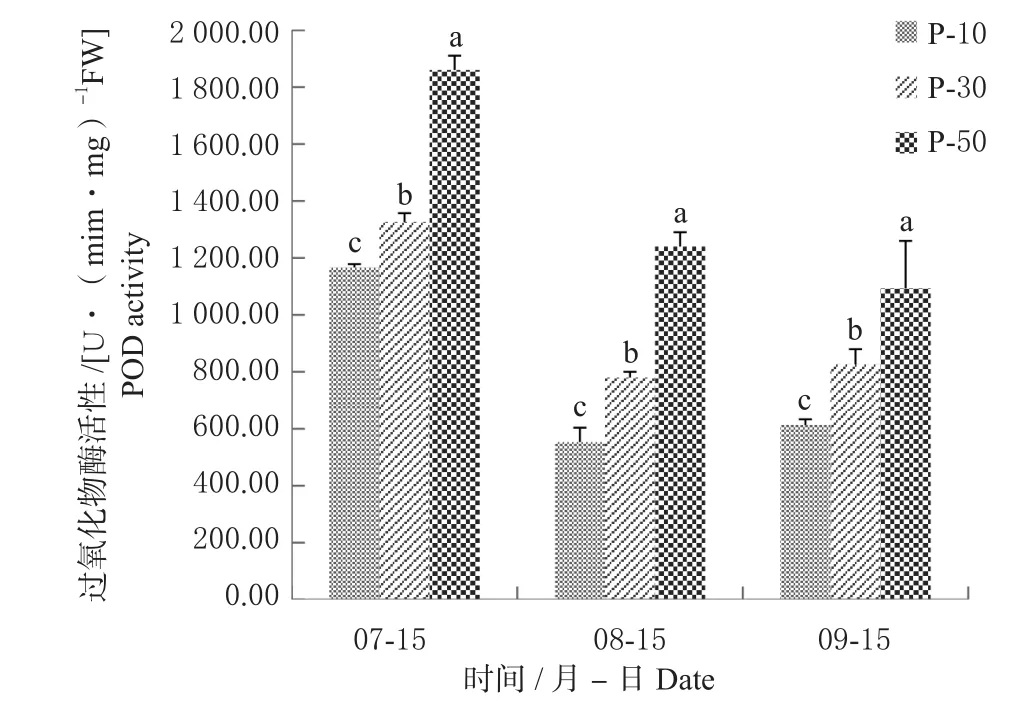
图5 矮化中间砧长度对叶片内POD酶活性的影响Fig. 5 Effect of dwarfing interstock length on POD activity in leaves
2.4 不同矮化中间砧长度对苗木叶片内激素含量的影响
不同矮化中间砧长度苗木的叶片内激素含量如表3所示,不同中间砧长度对红富士苹果叶片内激素含量具有显著影响。随矮化中间砧长度的增加,叶片内IAA含量减少,P-10处理的的IAA含量为57.66 ng/g,显著高于其它2组处理;叶片内ZR含量以P-30处理最高,为8.47 ng/g,显著高于P-10、P-50;叶片内KT含量以P-30处理最高,为14.53 ng/g,显著高于P-30、P-50;叶片内GA3含量结果表明,P-30处理的GA3含量最高,为15.62 ng/g,显著高于P-50。
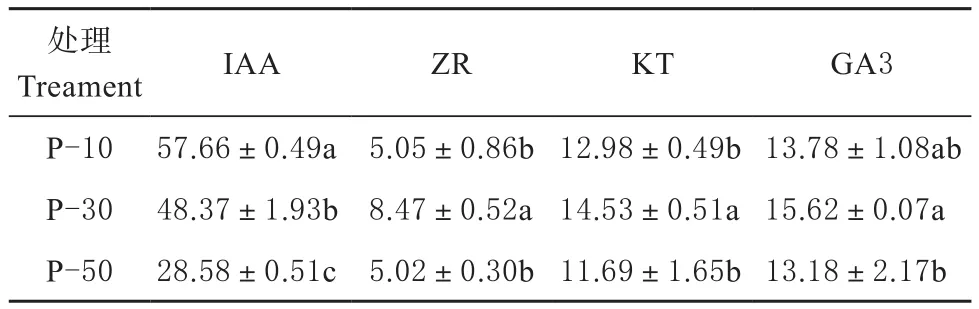
表3 矮化中间砧长度对叶片内源激素含量的影响Table 3 Content of endogenous hormones in leaves of interstock length ng/g FW
3 讨论与结论
本研究结果表明,无论是红富士苹果幼树还是苗木,其生长势均随着矮化中间砧长度的增加而减弱,一些学者在GM256、M9、MM106等砧木上也得到了相似结果[4-7],适当增加中间砧长度可增加矮化强度,至于中间砧长度对红富士苹果结果的影响还需进一步研究。
矮化中间砧长度在一定程度上抑制了营养物质的转运过程,中间砧长度越长,抑制品种生长的作用越大[4]。氮素是植物生长发育所必需的营养元素之一,在植物生长进程中具有极其重要的作用,被称为“生命元素”[18]。本研究中,随矮化中间砧长度的增加,中间砧干径、品种干径及新梢长度减少,与前人研究结果一致[4-7],当中间砧长为10 cm时,氮素分配量表现为中心干>中间砧,全株氮素分配量最高,树体长势最好,但对15N的利用率最低;当中间砧长为30 cm时,氮素分配量表现为中间砧>中心干,全株氮素分配量次于P-10,树体长势低于P-10,但新梢粗长比最大,能保证适宜的树体长势,15N利用率最高;当中间砧长为50 cm时,氮素分配量表现为中间砧>中心干,全株氮素分配量最低,树体长势最弱。这可能是由于矮化中间砧长度影响了苹果幼树各器官对氮素的吸收,中间砧长度过长,氮素在中间砧产生滞留,一定程度上阻碍了氮素在体内的运转,使其向上运输养分消耗增加,叶片与中心干所吸收的氮素减少;中间砧长度过短,降低了根系向上输送氮素的消耗,叶片与中心干所吸收的氮素增加。
徐继忠等[19]对苹果的研究及周怀军等[20]对李树的研究表明,IOD酶活性的高低与树体生长势呈显著负相关,认为IOD酶活性越高矮化作用越好。多数研究者认为叶片中过氧化物酶活性与树体生长势呈显著负相关[19-21],POD酶活性越高矮化作用越好。植物内源激素是植物适应环境必不可少的信号分子,对于调节植物的各种生长发育过程具有十分重要的作用[22],IAA含量与树体的生长势呈正相关关系。本研究中,随矮化中间砧长度的增加,叶片内IOD酶活性、POD酶活性呈上升趋势,叶片内IAA含量呈下降趋势,可能是高水平的IOD加强了对内源激素IAA的氧化分解,减轻了其对生长的刺激,使植物表现矮化,或者是高水平的POD一方面参与IAA的降解或失活,使IAA浓度下降,抑制了树体的生长,另一方面加速了木质化进程,使细胞的伸长过早停止所致。李巧霞等[23]的研究结果表明,GA3是控制树体大小最重要的激素。本研究中,随矮化中间砧长度的增加,叶片内GA3含量由大到小依次为P-30> P-10> P-50,当中间砧长度为50 cm时,GA3含量最低。CTKs是一类促进胞质分裂的物质,促进多种组织的分化和生长,较高含量的细胞分裂素有利于‘丹尼斯’凤梨的花芽诱导及孕育[24],与植物生长素有协同作用,调节植物细胞生长和发育。本研究中,随矮化中间砧长度的增加,叶片内ZR、KT含量由大到小依次为P-30> P-10>P-50,当中间砧长度为50 cm时,激素含量最低。可能是高水平POD可以使刺激植物生长的CTK浓度下降,抑制植物生长;同时使木质化程度加大,使植物表现矮化。
猜你喜欢
作物学报(2022年10期)2022-07-21
作物学报(2022年9期)2022-07-18
安徽农学通报(2022年9期)2022-06-14
作物学报(2022年3期)2022-01-22
今日农业(2021年17期)2021-11-26
鸭绿江·华夏诗歌(2021年6期)2021-09-13
落叶果树(2021年6期)2021-02-12
果农之友(2017年9期)2018-02-06
农村农业农民·B版(2017年11期)2017-12-06
农家科技下旬刊(2017年5期)2017-06-20
