
两种灌丛化草原小叶锦鸡儿的叶片化学计量特征
2021-11-09 00:46徐浩然俞富洋贾聪慧张国东
草地学报 2021年10期
徐浩然, 俞富洋, 贾聪慧, 张国东, 李 贺
(江苏师范大学地理测绘与城乡规划学院, 江苏 徐州 221116)
碳(Carbon,C)、氮(Nitrogen,N)和磷(Phosphorus,P)是生态系统的重要组成元素,在生态系统各个过程中起着至关重要的作用,同时也是植物体生长和代谢的必需元素[1-2]。C是植物体中含量最丰富的元素之一,植物通过光合作用将CO2转化为有机物,以建立植物基本结构并维持代谢功能;N是植物细胞蛋白质、核酸的组成元素并参与叶绿素的合成,与植物光合作用紧密联系;P是细胞酶和核酸的重要组成成分[3-4]。叶片C∶N和C∶P可反应植物利用养分以及同化碳的能力,也常用于表征植物N,P养分利用效率以及生长速率;N∶P能反映植物生长受外部环境限制的情况[5-7]。因此,研究植物叶片C∶N∶P的化学计量特征及其控制因素对阐明生态系统养分循环和植物适应环境的生态策略具有重要意义。
目前,已有许多研究表明水热和土壤条件是影响植物叶片化学计量特征的主要因素[8]。例如,樊江文等的研究发现内蒙古草地植物叶片N含量随着年平均气温的增加而增加;N,P含量随着年平均降水量的增加呈升高趋势[9]。Luo等对新疆荒漠植物的研究发现多数土壤因子对叶片N,P化学计量特征有直接影响,而气候因子年平均气温、年平均降水量则产生间接影响[10]。还有研究表明,不同地理单元中的气候、土壤因素差异是造成植物叶片N,P化学计量特征差异的重要原因[8]。然而,植物生长与气候和土壤条件的关系极为复杂,植物叶片的化学计量特征受遗传与环境的共同作用[11-13]。已有研究表明不同植物对环境条件的响应存在极大不确定性,而同种植物在不同环境条件下其化学计量特征也可能发生变化,这是植物化学计量特征的环境可塑性不同造成的[14-16]。而内蒙古典型草原降水比荒漠草原多,但年积温较低;土壤类型也与荒漠草原棕钙土不同,主要为栗钙土[17]。因此,两种草原类型中的同种植物叶片化学计量特征可能由于环境可塑性而存在差异,值得深入探究。
小叶锦鸡儿(Caraganamicrophylla)作为豆科(Leguminosae)灌木,是北温带草原典型灌木,具有耐寒、抗旱、耐贫瘠和耐高温等特点[18-19]。近年来由于气候变化等原因,小叶锦鸡儿的多度和密度在内蒙古草原地区有显著增加趋势,导致草原灌丛化现象,使草原生态系统从以草本植物为主的连续景观转变为木本植物与草本植物镶嵌的不连续景观,植被组成发生转变,对草原生态系统的结构与功能造成重要影响[20-22]。目前,关于小叶锦鸡儿叶片化学计量特征的生理特征、对人类活动的响应、对草原群落的影响、生态与营养价值以及幼苗化学计量特征等方面已有许多研究,但是对其叶片化学计量指标及其控制因素的研究还比较缺乏,小叶锦鸡儿叶片化学计量特征的环境可塑性也未有相关报道[18,23-27]。
综上所述,本研究通过分析不同区域小叶锦鸡儿叶片化学计量特征及其在不同水热与土壤条件下的差异,试图阐明植物叶片化学计量特征的环境可塑性和植物的资源利用策略,揭示植物资源利用与温度、降水、土壤等环境要素之间的相互关系,同时也能从资源利用的角度为小叶锦鸡儿在草原上多度和盖度增加的现象以及引起草原灌丛化发生发展的机制提供一定的解释,以期为我国北方灌丛化草原植被的保护和恢复工作提供参考。
1 材料与方法
1.1 研究区概况
本研究在内蒙古自治区锡林郭勒盟开展(北纬38.67~44.91°,东经108.28~119.02°,海拔800~1 200 m),共调查了19 个灌丛化草原样地。该地属于半干旱、干旱区,气候为典型的温带大陆性季风气候,夏季温暖湿润,冬季寒冷干燥,降水主要集中在气温较高的夏季。研究区年平均气温—0.7~4.8℃,气温年差较大,年平均降水量200~385 mm,湿度指数0.14~0.48,蒸散量463~600 mm。按照中国草地区划,本研究调查的灌丛化草原有11个样地为典型草原,8个样地为荒漠草原[28-29]。其中,典型草原区中的主要建群和优势物种有大针茅(Stipagrandis)、克氏针茅(Stipakryovii)、羊草(Aneurotepidimuchinense)等,而荒漠草原区中的建群种主要为小针茅(Stipaklemenzii)、沙生针茅(Stipaglareosa)、短花针茅(Stipabreviflora)等[30]。
图1 样本点示意图Fig.1 Diagram of sample sites
1.2 研究方法
1.2.1样地选择及样品采集 野外调查和采样于2019年进行,选择以草本植物为基质、灌木镶嵌的灌丛化草原(灌木盖度低于30%)为样地。根据自然分布情况,选取了无人类活动影响的11个典型草原区样地、8个荒漠草原区样地,每个样地之间相隔约30 km。为避免异质性导致的误差,每个样地设置3个20 m×20 m重复样方,每个样方的四周和中心选取自然生长状态相近、中等冠幅约60~80 cm左右的小叶锦鸡儿5~10 株,每株收集植株中部老枝上的成熟叶片10 g左右混合。在每个样方内分别使用五点法(样方四周和中心),用直径5 cm土钻采集5钻0~10 cm深度土壤样品。
1.2.2养分测定 野外采集的植物叶片在65℃烘干至恒重后研磨。土壤样品风干后过2 mm筛,仔细将里面的根系挑出后研磨。通过元素分析仪(Vario EL III,Elementar,Germany)测定叶片和土壤中的全碳、全氮含量,采用钼锑抗比色法测定全磷含量。
1.2.3气象数据的选择及处理 气候原始数据来自中国气象局国家气象信息中心(http://www.nmic.gov.cn),选取了1980—2019年各台站降水量和温度等数据,将台站数据进行克里格插值后得到了本研究使用的气候指标[年平均气温(Mean annual temperature,MAT)/℃、年平均降雨量(Mean annual precipitation,MAP)/mm和相对湿度(Relative humidity,RH)]。
1.3 数据处理与分析
采用单因素方差分析比较典型草原和荒漠草原小叶锦鸡儿叶片化学计量学特征的差异;采用皮尔逊相关性分析研究小叶锦鸡儿叶片化学计量学特征与土壤养分之间的关系,以及叶片化学计量学指标之间的关系;采用线性回归研究小叶锦鸡儿叶片化学计量学特征与MAT和MAP之间的关系。本研究所有统计分析与绘图均通过R4.0和其相关程序包实现[31]。
2 结果与分析
2.1 小叶锦鸡儿叶片化学计量特征
内蒙古草原小叶锦鸡儿叶片C,N,P含量均值分别为449.14 g·kg-1,34.39 g·kg-1,1.46 g·kg-1,变化范围分别为395.30~484.20 g·kg-1,22.70~43.80 g·kg-1,0.88~2.46 g·kg-1;叶片C∶N,C∶P,N∶P均值分别为13.26,322.94,24.29,变化范围分别为10.23~18.95,185.90~523.60,16.50~41.40。
两种草原类型中,小叶锦鸡儿的叶片化学计量特征差异显著(图2)。方差分析显示典型草原中小叶锦鸡儿叶片C含量显著低于荒漠草原,N,P含量显著高于荒漠草原,叶片C∶N,C∶P,N∶P均显著低于荒漠草原(P<0.05)。

图2 两种草原类型小叶锦鸡儿叶片化学计量特征Fig. 2 Leaf stoichiometry traits of Caragana microphylla in two grassland types注:不同小写字母表示叶片化学计量特征在两种草原类型间差异显著(P<0.05)Note:Different lowercase letters indicate significant differences in leaf stoichiometry traits between 2 grassland types at the 0.05 level
2.2 小叶锦鸡儿叶片-土壤的化学计量特征相关性分析
小叶锦鸡儿叶片与所处土壤化学计量特征存在显著相关性(表1)。叶片N含量与土壤C,N,P含量显著正相关;叶片C含量与土壤C,N,P含量及C∶P显著负相关;叶片P含量与土壤N,P含量显著正相关,与土壤C∶N显著负相关;叶片C∶N,C∶P均与土壤N,P含量显著负相关,叶片C∶P还与土壤C∶N显著正相关;叶片N∶P与土壤N含量及N∶P显著负相关,与土壤C∶N显著正相关(P<0.05)。
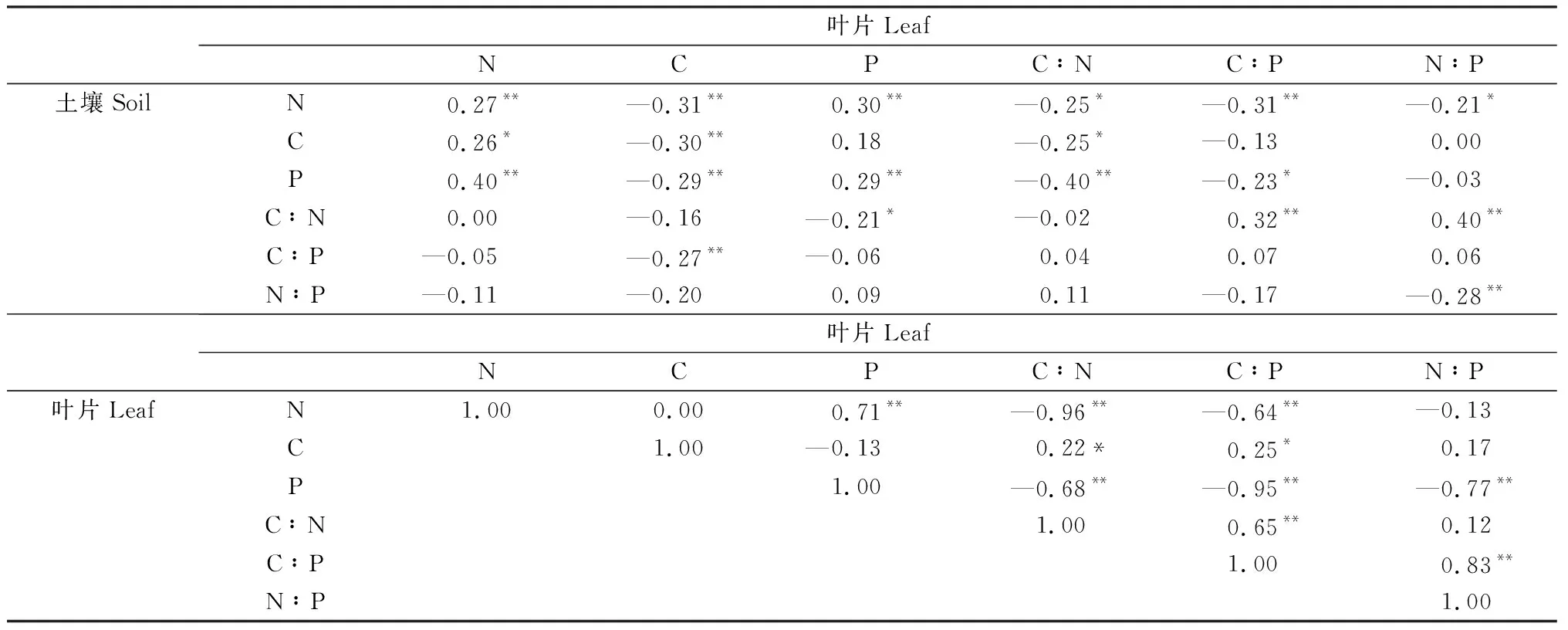
表1 小叶锦鸡儿叶片-土壤化学计量特征的相关性Table 1 Correlation between leaf-soil stoichiometry traits of C. microphylla
小叶锦鸡儿叶片化学计量指标之间也存在显著相关性。叶片C含量与叶片C∶N,C∶P显著正相关;叶片N含量与叶片P含量显著正相关;叶片C∶N,C∶P均与叶片N,P含量显著负相关,叶片C∶P还与叶片C∶N显著正相关;叶片N∶P与叶片P含量显著负相关,与叶片C∶P显著正相关(P<0.05)。
2.3 小叶锦鸡儿叶片化学计量特征与降水和温度的关系
由图3、图4可知,小叶锦鸡儿叶片C含量随年平均降水量(MAP)和年平均气温(MAT)的增加先上升后下降,回归方程均显著;叶片N,P含量均随MAP上升而显著增加,随MAT上升而显著下降;叶片C∶N,C∶P,N∶P均随MAP上升而显著下降,随MAT上升而显著上升(P<0.05)。

图3 小叶锦鸡儿叶片化学计量学特征与年平均气温的关系Fig.3 The relationship between leaf stoichiometry traits of C. microphylla and mean annual temperature

图4 小叶锦鸡儿叶片化学计量学特征与年平均降水量的关系Fig.4 The relationship between leaf stoichiometry traits of C. microphylla and mean annual precipitation注:*和**分别表示在0.05和0.01水平差异显著,ns表示无显著差异Note:* and ** indicate significant differences at the 0.05 the 0.01 level,respectivrly, ns indicate no significant difference
3 讨论
3.1 小叶锦鸡儿叶片的化学计量特征
通过比较发现,本研究小叶锦鸡儿叶片C含量高于内蒙古和阿拉善荒漠典型草原植物叶片均值[32-33]。小叶锦鸡儿叶片C含量较高的原因可能有两个:第一,可能是由于小叶锦鸡儿叶片C储存能力和光合作用能力较强,也可能与植物在叶片中积累含C化合物以适应土壤养分匮乏的环境有关[34]。第二,与草本植物相比,灌木植物具有大而深的根系,有研究表明天然小叶锦鸡儿根系可达1.8 m并有着广泛的水平分布范围,这可以使小叶锦鸡儿从周围环境吸收更多的养分元素以更好地供应植物生长[21,34-36]。
本研究中,小叶锦鸡儿叶片N含量高于内蒙古典型草原植物叶片均值(28.08 g·kg-1)和西北干旱区植物叶片均值(21.20 g·kg-1)[32,34]。小叶锦鸡儿叶片N含量较高的原因可能有如下几个:第一,小叶锦鸡儿是多年生豆科植物,具有根瘤固氮的特性[35];第二,小叶锦鸡儿个体较草本植物大,可能通过投入大量含N蛋白质酶以促进生长[36];第三,小叶锦鸡儿可能通过提高叶片非光合器官和组织N的投入,增加细胞内部的渗透压来提高水分利用效率,以适应草原干旱少雨的气候[37];第四,高叶片N含量会影响光合酶的活性和含量,提高CO2的同化速率,这可能使小叶锦鸡儿有更强的光合能力[38]。
小叶锦鸡儿叶片P含量在典型草原中略高于内蒙古典型草原植物叶片均值(1.56 g·kg-1)和西北干旱区植物叶片均值(1.40 g·kg-1),而在荒漠草原中低于这两个区域植物叶片均值[32,34]。这可能是因为叶片P含量主要受土壤P含量影响,而荒漠草原降水量少、气候干旱,降低了矿质的风化速率,使得土壤P含量相对较低[39-40]。
小叶锦鸡儿叶片C∶N均低于内蒙古典型草原植物叶片均值(16.67)和科尔沁沙地主要植物叶片均值(17.80)[32,41]。根据生长率假说(GRH),高生长速率的植物通常具有较低C∶N,从而将更多的N分配到核糖体RNA中以支持植物快速生长,这可能说明小叶锦鸡儿生长速率快[42-43]。小叶锦鸡儿叶片C∶P高于内蒙古典型草原植物叶片均值(302.6)和科尔沁沙地主要植物叶片均值(226.80)[32,41]。C∶P表征植物P利用效率,小叶锦鸡儿叶片C∶P较高说明其P利用效率高,这可能是因为植物需要提高P利用效率以适应草原土壤养分贫瘠的环境[44-45]。而荒漠草原中小叶锦鸡儿叶片C∶P显著大于典型草原。这可能与本研究中荒漠草原土壤P含量(0.18 g·kg-1)低于典型草原土壤P含量(0.21 g·kg-1),导致在荒漠草原中小叶锦鸡儿的生长受P限制程度更高有关。
3.2 小叶锦鸡儿叶片-土壤化学计量特征间的相关性
研究表明,如果植物生长受某种元素限制,那叶片内该元素浓度会与土壤提供此养分的能力呈正相关关系[41,46-47]。本研究中小叶锦鸡儿叶片P含量与土壤P含量、叶片N含量与土壤N含量显著正相关,表明小叶锦鸡儿生长受N,P限制[48-49]。而Gusewell等提出N∶P对植物生长受元素限制的情况同样具有指示意义:一般认为当N∶P<14时,植物生长主要受N限制;当N∶P>16 时,植物生长主要受P限制[49]。近期也有研究表明,相比于传统的临界比14和16,使用临界比10和20的误差风险更低[50-51]。本研究小叶锦鸡儿叶片N∶P大于20,表明小叶锦鸡儿生长受P限制,这与上文推论出的结论不一致。这可能是因为虽然研究区土壤N,P元素的含量(0.70 g·kg-1,0.20 g·kg-1)远低于全国平均水平(1.06 g·kg-1,0.65 g·kg-1),但豆科植物的固氮特性可以解决植物生长中N供应缺乏的问题[52-53]。因此,本研究认为基于N∶P的结果更为可靠,即小叶锦鸡儿的生长主要受P限制。
叶片C含量与N,P含量显著负相关,是陆生植物叶片化学计量的普遍性特征[33]。小叶锦鸡儿叶片C含量与N,P含量无显著相关性,有可能是因为研究区气候干旱导致N,P的吸收与C的同化不同步,即C和N,P的解耦[33,54]。叶片C含量与C∶N,C∶P呈显著正相关关系,这可能是因为叶片在C物质积累的过程中同时需要吸收和积累一定的N,P[55]。叶片N与P含量呈显著正相关,说明叶片中N,P元素二者耦合密切,这可能是由于植物在生长过程中需要消耗大量的ATP来合成植物的蛋白质,这符合植物元素计量关系的普遍规律,也是植物对生境适应良好的特征[56]。
3.3 小叶锦鸡儿叶片化学计量学特征的环境可塑性
尽管植物的叶片化学计量特征主要由遗传因素控制,但是环境因素的变化(如温度、降水等)也会影响植物的营养元素利用以及生理特性,而化学计量特征的可塑性可以使植物通过调整元素分配和养分利用效率等方式以适应生境变化[57-58]。
小叶锦鸡儿叶片C含量随MAP的增加先在荒漠草原中上升,后在典型草原中下降。叶片C含量在荒漠草原中上升可能因为植物主要通过光合作用获得C,干旱会抑制光合作用、减少C固定,而降水增加可以提高光合速率,使叶片积累更多C[55,59]。叶片C含量在典型草原中下降,可能因为植物常通过增加C在叶片组成化合物中的投资以适应干旱环境,随着降水增加叶片中的C投资相应减少[60]。
在两种草原类型中,小叶锦鸡儿叶片N,P含量均随MAP的增加而显著上升。这可能是因为在水分匮乏时,灌木植物对N,P的吸收利用率减慢,而降水增加可以提高植物对N,P的吸收利用率[61]。叶片P含量的上升,也可能与降水增加后土壤中P元素的供给增加有关[10]。由于N和P紧密相连,快速生长的植物需要增加叶片N,P含量来支持蛋白质等的合成;干旱地区植物叶片N含量的上升能提高叶片气孔导度对水势的敏感性,从而提高光合作用速率[10,17,60]。
小叶锦鸡儿叶片C含量随MAT的增加先在典型草原中上升,后在荒漠草原中下降(P<0.05)。叶片C含量在典型草原中上升,可能是因为温度上升使光合酶等的活性提高,植物光合作用增强,导致叶片积累更多的C[61]。叶片C含量在荒漠草原中下降,可能是因为植物在相对低温下具有更高的光合速率,到达最大速率之后随着温度的上升光合速率下降[61]。
小叶锦鸡儿叶片N,P含量,在两种草原类型中均随MAT的下降而显著上升(P<0.05)。可能有如下两个原因:第一,根据Reich和Olesykn的“温度-植物”生理假说,植物的N,P调节机制是温度敏感型[62],在低温下,小叶锦鸡儿可能会通过提高叶片N,P含量,来抵消生物化学反应速率降低的效应[42,60,62];第二,根据生物地球化学假说,低温会抑制土壤有机物的分解和矿化,减少N,P的可利用性,一般认为会降低叶片N,P含量,但宋璐璐等的研究表明,植物也会通过“奢侈消费”来抑制营养的流失以适应贫瘠的土壤[62-63]。
由上述结果可知,气候因子(MAP,MAT) 变化时,小叶锦鸡儿叶片C含量随MAT,MAP的变化趋势与N,P含量不同,这可能是由于干旱胁迫导致C和N,P的解耦;而叶片N和P含量随MAT,MAP的变化趋势相似,说明它们耦合密切[60]。
在两种草原类型中,小叶锦鸡儿叶片C∶N,C∶P均随MAP下降而显著上升(P<0.05)。可能有如下两个原因:第一,小叶锦鸡儿会提高N,P利用效率以适应干旱生境,也可能将更多N,P分配至植物根系以促进水分吸收[28,51];第二,随着MAP的变化,小叶锦鸡儿C,N,P之间的耦合关系发生改变,体现了其对降水的弹性适应能力[64]。叶片N∶P随MAP上升而显著下降P<0.05)。根据生长速率假说,植物生长速率随植物N∶P的降低而增加[43,63,65]。叶片N∶P下降,可能与MAP增加后,小叶锦鸡儿的生长速率得到提高有关。
在两种草原类型中,小叶锦鸡儿叶片C∶N,C∶P均随MAT增加而显著增加(P<0.05)。这可能表明MAT上升后,小叶锦鸡儿对N,P养分的利用效率提高[42]。叶片N∶P随MAT下降而显著下降(P<0.05)。Gong等的研究提出:在MAT较低时,叶片N∶P较低的植物可以匹配较高的生长速率[34]。N∶P升高,也意味着小叶锦鸡儿更容易受P限制[58]。此外,叶片N∶P与水分利用效率呈正相关,N∶P升高也可能与MAT增加后,植物水分利用效率提高有关[66]。
综上所述,小叶锦鸡儿可通过调整自身的化学计量特征来适应气候因子的变化,具有环境可塑性[57-58]。根据IPCC《气候变化2021:物理科学基础》和《全球升温1.5℃报告》,中纬度干旱和半干旱地区的降水量可能会减少,升温速度可能达到全球平均变暖速度的1.5到2倍,而亚洲东部等地的干旱程度会进一步加剧[67-68]。小叶锦鸡儿可能会采取增加C投资、提高N和P利用效率以及水分利用率、提高光合速率和生长速率等方式以应对气候变化,这有可能会继续加剧干旱区、半干旱区的草原灌丛化现象。
4 结论
研究区小叶锦鸡儿叶片C,N含量以及C∶P,N∶P较高,P含量和C∶N较低;叶片化学计量特征在两种草原类型中存在差异;叶片化学计量特征与所处土壤化学计量特征之间存在显著相关性(P<0.05);叶片C与N,P含量无显著相关性,叶片N与P含量呈显著正相关关系(P<0.05);叶片N和P含量随气候因子(MAP,MAT)的变化趋势相似,而叶片C含量在两种草原类型中表现出相反的变化形式;叶片N∶P>20,小叶锦鸡儿生长主要受P限制。
猜你喜欢
种子(2022年5期)2022-06-27
小学生优秀作文·时尚版·中年级(2022年2期)2022-02-18
散文诗(2021年22期)2022-01-12
散文诗(2021年22期)2022-01-12
High Technology Letters(2021年4期)2022-01-09
河北果树(2020年4期)2020-11-26
防护林科技(2020年5期)2020-07-21
水土保持研究(2020年1期)2020-04-27
海峡姐妹(2019年1期)2019-03-23
中成药(2018年9期)2018-10-09
