
种子大小和原位埋藏对17种高寒草甸植物种子萌发率和死亡率的影响
2021-11-09 02:00任彦梅孙建平傅延锦宁鑫蕊周华坤张春辉
草地学报 2021年10期
李 聪, 马 真, 武 林, 任彦梅, 孙建平, 傅延锦, 孟 洁,宁鑫蕊, 周华坤, 张春辉,2*
(1.青海大学生态环境工程学院省部共建三江源生态与高原农牧业国家重点实验室, 青海 西宁 810016; 2. 中国科学院西北高原生物研究所青海省寒区恢复生态重点实验室, 青海 西宁 810008;3. 中国科学院高原生物适应与进化重点实验室,青海 西宁 810008; 4.青海大学农牧学院, 青海 西宁 810016)
种子萌发是植物生活史中不可或缺的阶段,一直是备受关注的研究热点[1-2]。种子的萌发特征影响着幼苗存活能力[3]、个体适合度[4]和植物生活史的表达[5]。种子萌发率是植物种群繁殖能力的表现,种子的成功萌发直接关系着物种的繁衍与生存[6]。因此,对种子萌发性状的研究已成为群落生态学和进化生态学研究的重要课题。
种子萌发是一个复杂的多步骤过程,容易受内外因素的影响,内因素包括种子自身的发芽力以及解除休眠的能力,外因素主要为环境条件,如光照、水分等[7]。相关的研究大多集中于光照[2,8]、温度[9]和水分条件对种子萌发特性的影响[10]。光照主要作为刺激信号,打破种子休眠使种子萌发[11],除光照条件以外,温度和水分也是种子萌发所必须的条件,萌发环境温度和水分含量可以决定种子萌发速率[12]。
自然条件下,种子成熟脱落后进入土壤,所以种子在土壤中埋藏的阶段是植物生活史的重要阶段。埋藏可以影响种子所处环境的温湿度和光照条件,这些环境条件与种子萌发[13]、幼苗存活[14]等关系密切。埋藏土壤的理化性质(例如土壤pH)和土壤微生物(特别是病原微生物)可以影响种子的休眠程度和存活状态[15],从而影响植物种群更新过程和种群动态。因此,深入探究埋藏对植物种子萌发率影响,可以为植被恢复和多样性保护提供科学依据。
种子大小是种子植物诸多性状中的一个重要性状,能影响种子的萌发能力以及群落的结构和动态[16-17]。种子大小决定种子内部储存营养物质的含量,一般而言,大的种子具有更强的竞争优势和存活能力[18-19],而小种子在数量和传播方式方面更具有优势[20]。种子大小与种子萌发率呈显著负相关关系[21],与幼苗的存活率呈正相关关系[22]。也有研究表明,种子大小与萌发率之间没有直接的相关性[23]。然而,关于种子大小和萌发率之间关系的研究一般没有涉及埋藏处理。因而在种子埋藏处理下,研究种子大小和萌发率的关系是种子生态学研究中的一块空白。
青藏高原是世界上海拔最高的地区,被称为“世界第三极”,是全球变化和人类干扰的敏感区,也是生物多样性热点区域[24]。高寒草甸是青藏高原分布面积最大且类型最多的生态系统,是适应高原隆起与长期低温环境形成的特殊产物,作为青藏高原典型的地带性植被,高寒草甸生态系统极为脆弱且部分地区退化严重[24]。种子是退化草地恢复的重要来源,研究青藏高原高寒草甸种子萌发对环境因子的响应及其适应局部环境的生态机制,可为高寒草地种质资源的利用和保护提供理论依据,并为高寒草地植物种群的保育、退化草地恢复等方面提供支持。
1 材料与方法
1.1 研究区域概况
试验所用植物种子采集及埋藏地点位于青藏高原东北部青海省门源回族自治县境内的中国科学院海北高寒草甸生态系统定位站(37°37′ N,101°12′ E),海拔3 200 m;年降水量561 mm,降水主要分布于6—9月;年平均气温0.8℃,全年无绝对无霜期,相对无霜期20 d;属于典型的高原大陆性气候。
1.2 供试材料
本试验以青藏高原高寒草甸的17种植物种子为研究对象并进行埋藏试验。详情见表1。
1.3 试验种子处理及称量
本试验所用17种植物种子(见表1)于2019年9—10月的种子成熟期采集。每一个物种采集选取2~3个种群,每一种群的种子随机在20株以上的个体中均匀的采集,采集后的种子充分混合。将采集到的种子带回实验室进行清理,风干后装入信封中在15℃室温下储藏待用。

表1 高寒草甸17种植物物种以及种子大小Table 1 Species name and Seed size of 17 plants in alpine meadow
将处理好的每种植物种子充分混合,随机选取饱满健康的种子进行试验,并用万分之一的电子天平称量每种植物种子的百粒重,重复3次。埋藏试验前,每一物种数出3份种子,每份50粒种子,进行四氮唑试验检验种子活性,并计算每一物种的活种子率。后续的种子萌发率和死亡率的计算都基于活种子率进行计算。
1.4 埋藏试验
试验包括两个处理(对照处理和原位埋藏处理),每个处理8个重复,每个重复30粒种子,30粒种子放于一个尼龙网袋内。每一物种包括16个尼龙网带,共计272个尼龙网袋,于2019年11月5日运往海北站。其中8包分别埋藏于8个地点,每一地点相隔10 m,埋藏深度为15 cm,称为(原位)埋藏处理。另外8包放在埋藏点附近的彩钢房内(可以挡雨,冬季无任何取暖措施),称为对照处理。第二年6月15日,把每一物种的8份种子运回实验室。
1.5 种子死亡率测定试验
在实验室内,随机取埋藏处理每一物种8份种子中的4份(剩余4份种子用于萌发试验),用蒸馏水清洗埋藏的种子,统计埋藏的每一尼龙网袋中腐烂的和消失的种子数,这些种子算为土壤中死亡的种子。其他种子进行四氮唑试验检验种子活性,得到剩余活种子数,进而得到种子死亡数[25-26]。这两部分数据相加,得到埋藏处理的种子死亡率。
随机取对照处理每一物种8份种子中的4份(剩余4份种子用于萌发试验),直接进行四氮唑试验检验种子活性,得到剩余活种子数,进而得到种子死亡数及对照处理的种子死亡率。
1.6 萌发试验
在实验室内,用蒸馏水清洗埋藏和未埋藏的种子,去除腐烂和不完整的种子,其他种子放在铺有2层滤纸的培养皿(直径9 cm)内,加入蒸馏水至滤纸充分湿润,放于培养箱内。培养箱温度设置为20℃/10℃,光照设置为14 h光照/10 h黑暗。每天检查培养皿缺水情况,补充适量蒸馏水,挑出发芽种子。萌发试验进行45天后,连续7天培养皿内没有发芽种子视为萌发结束。
1.7 数据分析
1.7.1萌发指标及计算方法: (1) 萌发率(Germination proportion)=(试验种子达到完全萌发时种子总数/试验处理活种子总数)×100%;
(2) 种子死亡率(Seed mortality rate)=(试验中种子死亡数/试验处理种子总数)×100%。
萌发率用试验前的活种子率和埋藏处理或对照处理的种子死亡率矫正;种子死亡率用试验前的活种子率矫正。
1.7.2数据统计与分析方法 数据分析之前,萌发率和种子死亡率经过反正弦平方根转化;种子重量经过对数(log10)转化。使用SPSS 22.0 软件进行数据统计与分析,用T检验(T test)检验显著性水平,应用一般线性回归检验种子大小与萌发率和死亡率的相互关系。作图使用Origin 9.1软件。
2 结果与分析
2.1 埋藏对种子萌发率的影响
如图1所示,17种植物种子的萌发率存在差异(对照:F=27.17,P<0.001;埋藏:F=122.32,P<0.001)。在对照处理下,17种物种的平均萌发率为52.25%,变化范围在0(高山嵩草(Kobresiapygmaea)和扁蕾(Gentianopsisbarbata))到99.94%(冷地早熟禾Poacrymophila)之间。对照处理下有6种植物种子的萌发率在80%~100%之间,占物种总数的35.29%;2种植物种子的萌发率在60%~80%之间,占物种总数的11.76%;1种植物种子萌发率在40%~60%之间,占物种总数的5.88%;2种植物种子萌发率在20%~40%之间,占物种总数的11.76%;6种植物种子萌发率小于20%,占物种总数的35.29%(图1)。埋藏处理下,17种物种的平均萌发率为45.40%,变化范围在0(高山嵩草和扁蕾)到100%(美丽风毛菊(Saussureapulchra)和垂穗披碱草(Elymusnutans))之间。埋藏处理下有7种植物种子的萌发率在80%~100%之间,占物种总数的41.18%;0种植物种子的萌发率在40%~80%之间;3种植物种子萌发率在20%~40%之间,占物种总数的17.65%;7种植物种子萌发率小于20%,占物种总数的41.18%,其中扁蕾和高山嵩草种子发芽率为0(图1)。整体上,埋藏处理没有显著改变种子的萌发率(t=1.74,P=0.101)。
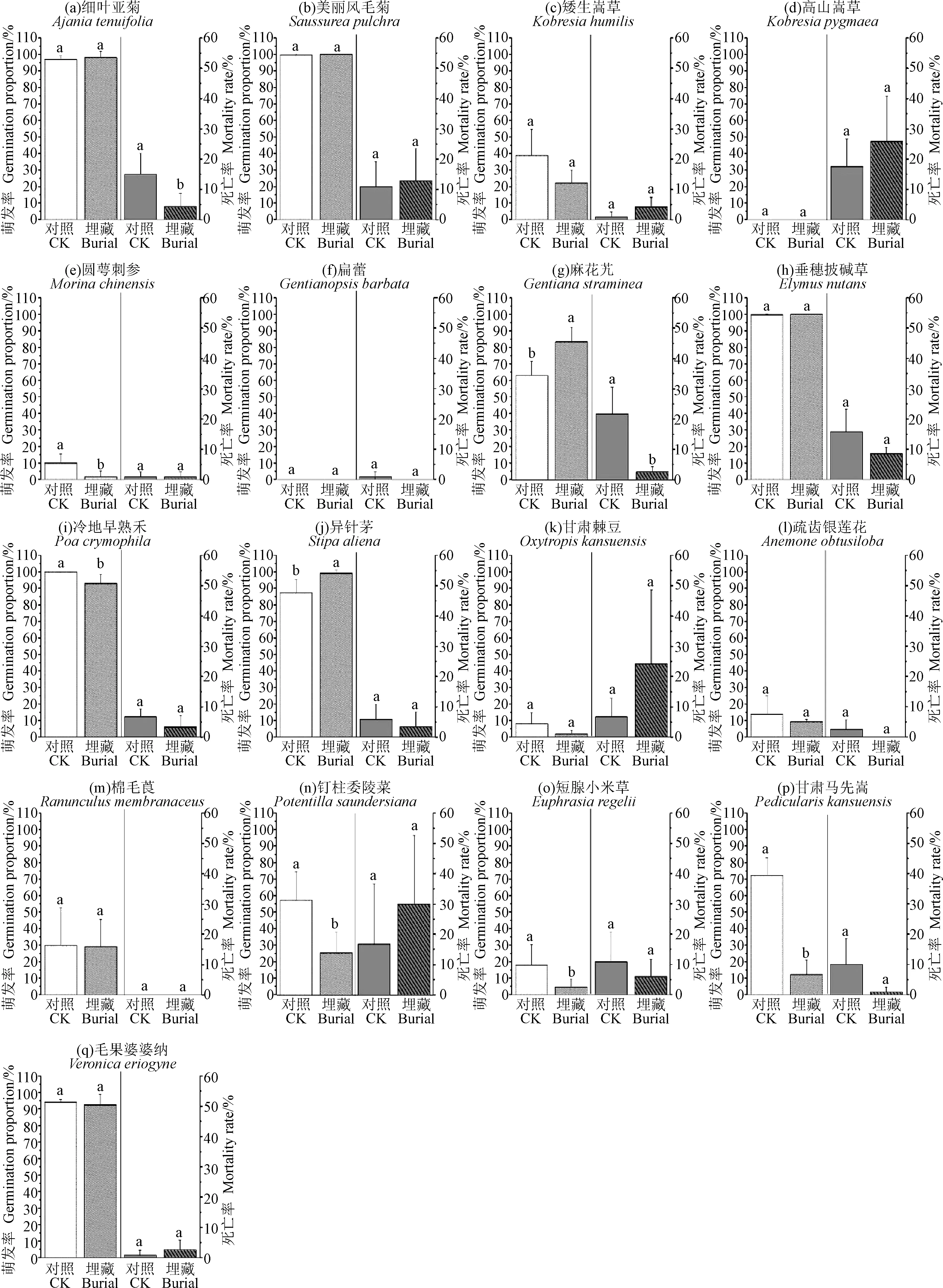
图1 17种高寒草甸植物种子萌发率和死亡率Fig.1 Seed germination proportion and mortality rate of 17 species of alpine meadow 注:误差棒为标准误;不同字母表示差异显著(P<0.05)Nome:The error bars represent standard error;Different letters indicate significant difference at the 0.05 level
埋藏处理可以显著提高麻花艽(Gentianastraminea)和异针茅(Stipaaliena)的种子萌发率(P<0.05),与对照相比,分别提高了20.50%和11.65%。埋藏处理显著降低了圆萼刺参(Morinachinensis)、冷地早熟禾、钉柱委陵菜(Potentillasaundersiana)和甘肃马先蒿(Pediculariskansuensis)的种子萌发率(P<0.05),与对照相比,分别降低了8.33%,6.84%,32.12%和59.94%。另外,埋藏处理边际显著地降低了短腺小米草(Euphrasiaregelii)的种子萌发率(t=2.31,P=0.06),与对照相比,降低了13.57%。埋藏处理对于其他物种的种子萌发率没有显著影响。
2.2 埋藏对种子死亡率的影响
如图1所示,17种植物的种子死亡率存在差异(对照:F=3.85,P<0.001;埋藏:F=3.85,P<0.001)。在对照下,17种物种的平均死亡率为8.43%,变化范围在0(棉毛茛Ranunculusmembranaceus)到21.67%(麻花艽)之间。对照处理下有1种植物的种子死亡率在20%~30%之间,占物种总数的5.88%;7种植物的种子死亡率在10%~20%之间,占物种总数的41.18%;9种植物的种子死亡率小于10%,占物种总数的52.94%(图1)。埋藏处理下,17种物种种子的平均死亡率为7.60%,变化范围在0(扁蕾、疏齿银莲花(Anemoneobtusiloba)和棉毛茛)到30%(钉柱委陵菜)之间。埋藏处理下有3种植物的种子死亡率在20%~30%之间,占物种总数的17.65%;1种植物种子死亡率在10%~20%之间,占物种总数的5.88%;13种植物的种子死亡率小于10%,占物种总数的76.47%,其中扁蕾、疏齿银莲花和棉毛茛的种子死亡率为0(图1)。在整体上,埋藏处理边际显著地改变了种子死亡率(t=1.85,P=0.083)。
埋藏处理显著降低了细叶亚菊(Ajaniatenuifolia)和麻花艽的种子死亡率,与对照相比,分别降低了10.65%和19.14%(图1)。对于其他物种,埋藏处理对于种子死亡率没有显著影响(图1)。
2.3 对照和埋藏处理下种子大小与萌发率和死亡率的关系
在对照处理下,种子大小与种子萌发率和死亡率均无显著相关关系(表2);在埋藏处理下,种子萌发率和死亡率与种子大小也均无显著相关关系(表2)。

表2 对照和埋藏处理下种子大小与萌发率和死亡率的回归分析Table 2 Regression analysis of seed size with germination proportion and mortality rate under the control and burial treatments
3 讨论
种子萌发阶段是植物生活史的关键阶段,种子萌发阶段的特征是植物生活史的重要特征。种子休眠是种子在适应环境过程中产生的特殊机制[26-27]。种子存在休眠现象的意义在于延长种子的存活时间,更容易形成土壤种子库。埋藏对种子萌发会产生3种影响:1)种子萌发;2)种子没有萌发,且可能受到其他生物破坏;3)种子休眠,成为土壤种子库的一部分[28]。青藏高原海拔较高,年平均温度低,降水分布不均匀,紫外线强,空气氧含量低,且各种环境因子的变化具有随机性和无法预测性。植物生活在这样的特殊环境之中,会产生不同的生态萌发对策[2,8]。本研究表明,埋藏对种子休眠程度和种子存活率具有一定的影响,进而影响植物的生态适应策略及其更新机制。
3.1 萌发率
植物种子在合适的条件下发芽,或是在不利的环境中处于休眠状态,这是植物适应逆境并保持物种延续的一种生态策略[26,29-30]。不同物种对埋藏的响应存在差异。埋藏处理下萌发率增高的物种为麻花艽和细叶亚菊,从萌发格局分类,它们属于埋藏促进型。这可能是由于土壤的理化性质和微生物环境促进了种子休眠状态的打破。而埋藏处理萌发率减少的物种包括冷地早熟禾、短腺小米草、甘肃马先蒿、圆萼刺参和钉柱委陵菜,从萌发格局分类,它们属于埋藏抑制型。这可能是这些物种具有不同的适应机制或者生态对策,它们萌发率降低表示这些物种具有一定的风险平摊对策[2,31]。另一方面,研究发现埋藏处理可以减少种子的死亡率,从而使种子处于一个相对低死亡率的环境中,进而促进了种子休眠。另外,原位埋藏使种子处于一个湿润、低氧和相对恒定的环境中,这样一个环境可能促进了种子休眠。对于其他10个物种,埋藏没有显著影响它们的萌发率,这10个物种在萌发格局上属于埋藏不敏感型。总之,整体水平上埋藏对种子萌发率无显著影响,但不同物种对埋藏有着不同的响应。这也反映了高寒草甸物种面对埋藏有着多样性的生态对策,进而促进高寒草甸群落物种共存。
3.2 死亡率
研究发现埋藏处理整体上降低了种子死亡率,其中,麻花艽和细叶亚菊的种子死亡率对埋藏处理的响应最为明显。这说明埋藏处理更有利于保存种子,从而促进土壤种子库的形成。植物种子长期在土壤种子库中保持活力,可以减小物种在恶劣环境中灭绝的风险[23,32]。李海静等[33]研究埋藏对沙芥(Pugioniumcornutum)和斧翅沙芥(Pugioniumdolabratum)种子死亡率的影响,发现种子在埋藏处理下可能受到真菌感染或者是遇到不适宜其存活的微环境,导致其死亡率偏高,和本研究结果不同,这可能是因为高寒生态系统的温度较低,影响种子存活的病原微生物较少。在高寒生态系统,埋藏处理有利于种子的长期保存,可以促进土壤种子库的形成,种子进入土壤(类似埋藏)是形成土壤种子库的第一个步骤,也是最为关键的一个步骤。
3.3 种子大小与萌发率和死亡率的关系
种子大小是植物生活史的重要性状,与植物的竞争能力或种群扩散能力密切相关。植物群落中不同物种会产生不同大小的种子(有的群落中种子大小差异高达106)[31],而种子大小反映了种子内部储存营养物质的含量,因而一般认为不同种子大小的物种萌发率也不相同[34]。本研究发现种子大小与萌发率无相关关系,和宗文杰等[35]对青藏高原51种菊科植物的研究结果一致。在张玲卫等[36]对荒漠化生态系统的研究中也得到了类似的结果,但朱雅娟等[37]的研究表明荒漠化生态系统的种子大小与萌发能力呈现负相关的关系。一般认为,小种子更容易形成土壤种子库,也就是说小种子应该在土壤中有着高的存活率。然而,本研究结果也不支持这一观点。因此,种子大小与种子萌发和死亡之间的关系需要进一步的详细研究。
4 结论
对埋藏处理下17种高寒植物种子休眠程度和死亡率的影响研究表明:种子大小与种子萌发和死亡不相关;埋藏处理可以促进或打破种子休眠,是促进土壤种子库形成的潜在机制。但不同的物种对埋藏有着不同的响应,这反映了高寒草甸物种面对埋藏有着多样的生态对策,进而促进了高寒草甸群落物种共存。
猜你喜欢
防护林科技(2020年11期)2020-12-30
江苏农村经济(2019年5期)2019-01-14
中国食品学报(2019年3期)2019-01-13
小学生学习指导(低年级)(2018年9期)2018-09-26
读者·校园版(2018年1期)2017-12-16
数学大王·低年级(2017年10期)2017-10-31
数学小灵通(1-2年级)(2017年3期)2017-04-16
蔬菜(2016年10期)2016-03-27
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15
少年科学(2015年2期)2015-08-12
