
早老素在日本血吸虫生殖发育中的作用
2021-11-09 11:59:24杨旭晗沈际佳
安徽医科大学学报 2021年10期
杨旭晗,杨 菲,沈际佳,刘 淼
血吸虫病是热带地区高发的寄生虫病之一,虫卵是其传播和致病最重要的因素。血吸虫雌虫性器官的发育成熟和产卵必须通过雌雄虫合抱方可完成。可能原因为雌雄合抱在虫体内启动了特殊基因的表达,这些基因可能参与了调节血吸虫雌虫的性成熟和产卵过程。但是,雌雄合抱对雌性生殖系统发育的影响及分子机制尚未完全确定。
早老素 (presenilin, PS) 在多种动物体内均可表达,具有多种生物学作用。但其在血吸虫虫体生殖发育中的作用尚未有报道。该研究首先运用荧光定量PCR技术分析早老素基因在血吸虫发育各个阶段的表达情况,随后通过RNA干扰(RNA interference,RNAi)成虫的早老素基因,初步证实早老素在日本血吸虫生殖系统发育中的作用。
1 材料与方法
1.1 实验动物
雌性6周龄昆明鼠由安徽省实验动物中心提供,阳性钉螺(含日本血吸虫尾蚴)购自湖南省血吸虫病防治研究所。1.2 主要试剂
Tween-20及DEPC购自美国Solarbio 公司; TRIzol购自美国Invitrogen 公司; ProteinaseK、LipofectamineRNAiMAX Reagent、荧光定量PCR试剂盒(SYBR Green)、10×PCR buffer、dNTPmix及逆转录试剂盒(PrimeScriptRT reagent Kit)购自日本TaKaRa&Clontech公司;三抗及引物委托上海生工生物公司合成; RPMI1640、无添加剂的M199培养基、OPTI-MEM及胎牛血清(FBS)、糖原购自美国Gibco公司;其余试剂均为国产分析纯。1.3 日本血吸虫雌雄虫的收集
准备加满纯净水的10 ml锥形瓶,并将感染日本血吸虫的钉螺放置于瓶中,于 25 ℃、通风条件下光照4 h,然后收集从钉螺中逸出浮于水面的血吸虫尾蚴。每只昆明鼠裸露腹部皮肤,给予80只左右的尾蚴经腹部皮肤感染,昆明鼠分别在感染血吸虫后16、18、24、28 d处死,收集虫体则使用肝门静脉灌注法,用PBS充分冲洗虫体3次,将虫体上黏附的其他组织清洁干净,分离雌虫及雄虫,并分别储存在-80 ℃、无RNA酶环境中备用。1.4 日本血吸虫雌虫卵巢分离
取50~60条28 d的雌性血吸虫放入离心管里,用PBS清洗,加入表皮消化剂,在37 ℃水平摇床消化5 min,弃去消化剂,消化4次,然后加入弹性蛋白酶,当雌虫整个虫体呈现松弛的团状时终止,将虫体倒入平皿中,加入PBS,移液器吹打使卵巢从虫体释放。洗涤去卵巢上的附着组织,将卵巢吸取收集,储存在-80 ℃备用。1.5 日本血吸虫总RNA的提取
利用TRIzol试剂分别提取感染不同时期的血吸虫的总RNA。1 mg虫体加入1 ml TRIzol试剂,使用常规酚氯仿法对总RNA进行抽提。RNA浓度用NanoDrop2000测定并记录,按照TaKaRa公司的逆转录试剂盒说明书的步骤进行cDNA合成。利用Primer Premier5软件设计荧光定量PCR引物,设计好的引物由上海生工生物公司进行合成。设计合成的引物序列如表1。加样操作按照荧光定量PCR试剂盒(SYBR Green)的说明进行,而后PCR过程按95 ℃ 10 s,95 ℃ 15 s、60 ℃ 40 s(40 个循环),95 ℃ 15 s、60 ℃ 1 min、95 ℃ 15 s完成,结果分析通过ABI StepOne software进行。
表1 早老素及PSMD4的荧光定量PCR引物序列
1.6 RNA干扰
根据SjPS的mRNA,利用Thermo Fisher在线软件(https://rnaidesigner.thermofisher.com)设计小干扰RNA(siRNAs)的序列,送至上海吉玛基因公司合成,并合成1管小干扰RNA为阴性对照,序列见表2。首先处死感染血吸虫尾蚴28 d后的昆明鼠,利用肝门静脉灌注法取出血吸虫虫体并筛选出合抱的血吸虫至完全巴赫培养基培养,首先使用3个不同的siRNA对体外培养的成虫进行RNAi,同时设置DEPC水组(Control)、非特异性siRNA组(negative control, siNCTRL)。根据LipofectamineRNAiMAX Reagent说明书上的操作步骤转染siRNAs,3 d后收集培养的血吸虫虫体并提取总RNA,各组的干扰效率通过荧光定量PCR技术计算并分析。使用同样条件进行3次平行重复实验后,确定干扰效率最高的SjPS siRNA1。打乱所选取的小干扰RNA序列,使用NCBI中的BLAST比对证实其无特异性后,送至上海吉玛基因公司,合成错配的siRNA,序列见表1。分别设置DEPC水组(Control)、SjPS SiRNA1组与siRNA1错配组(Mock),并分别于体外作用成虫10 d,每天进行半量换液,培养10 d后于显微镜下计算合抱与未合抱的虫体数量和虫卵数量,后收集所有培养的虫体,一半提取总RNA,使用荧光定量PCR技术计算出siRNA的干扰效率,另一半虫体使用多聚甲醛固定后,利用中性树胶封片,使用激光共聚焦显微镜观察干扰后血吸虫虫体内生殖系统的形态学变化。
表2 siRNA的序列
1.7 激光共聚焦观测日本血吸虫生殖系统
从培养基中挑出培养的血吸虫完整虫体,并使用PBS清洗,将雌虫与雄虫分离后,分别置入多聚甲醛固定液中固定,使用盐酸卡红将固定好的虫体染色24 h,再脱色,待虫体呈现透明状态后,再转移到载玻片上,使用中性树胶进行封片。在反射模式下,将激光共聚焦扫描显微镜设置为470 nm波长的滤波器以及488 nm波长的He/Ne激光器,观察日本血吸虫两性生殖系统结构形态是否发生变化。1.8 统计学处理
统计分析使用GraphPad Prism软件(6.0版本)进行。所有试验全部独立重复 3 次及以上,组间比较采用单因素方差分析,事后两两样本之间多重检验采用Tukey法,将各基因的相对表达量、虫卵量以及雌雄合抱对数表示为均数±SEM,P
<0.05为差异有统计学意义。2 结果
2.1 早老素在合抱前期、初期及后期的血吸虫雌虫表达情况
应用荧光定量PCR技术分析比较早老素在血吸虫感染16、18、24 d的雌性日本血吸虫虫体内的表达情况。选择PSMD4为管家基因,标准化反应,运用2方法分析早老素mRNA的相对表达量,结果见图1,从结果分析可知合抱后期的雌虫体内早老素相对表达量(89.26±6.31)高于其他两组,差异有统计学意义(F
=196.0,P
<0.001)。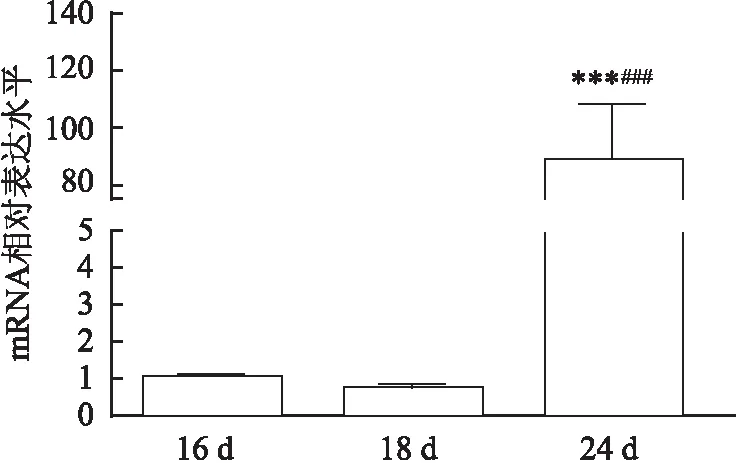
图1 早老素在合抱前期、初期及后期的雌虫表达水平与16 d比较:***P<0.001;与18 d比较:###P<0.001
2.2 早老素在血吸虫雌性成虫、雄性成虫及雌性成虫卵巢的表达情况
应用荧光定量PCR技术分析比较早老素mRNA在血吸虫雌性成虫、雄性成虫及雌性成虫卵巢的表达水平。结果发现,早老素蛋白在雌性血吸虫的卵巢中相对表达量(3.00±0.19)高于雌性及雄性血吸虫虫体,并且与其他两组的差异有统计学意义(F
=118.4,P
<0.001)。而雌性血吸虫虫体相对表达量(1.04±0.05)及雄性血吸虫虫体相对表达量(0.76±0.04)之间差异无统计学意义,见图2。
图2 早老素在雌性血吸虫成虫虫体、雄性血吸虫成虫虫体及雌性血吸虫成虫卵巢中的表达水平与雌性成虫虫体比较:***P<0.001;与雄性成虫虫体比较:###P<0.001
2.3 利用siRNA干扰血吸虫成虫早老素基因
利用siRNA干扰技术,对体外培养的4周龄血吸虫成虫的早老素基因进行干扰。首先,使用针对早老素表达设计的3个不同的siRNA对体外培养的血吸虫成虫进行干扰,干扰3 d后收集虫体,利用荧光定量PCR技术计算出各组早老素的表达水平,从结果分析中能够发现,在设计的3个siRNA中,SjPS siRNA1干扰后早老素的相对表达水平(0.34±0.028)远低于其他两组,差异有统计学意义(F
=45.18,P
<0.001),见图3,故选择干扰效率较高的SjPS siRNA1进行下一步实验。然后利用SjPS siRNA1对体外培养28 d成虫的早老素基因进行干扰,于10 d后收集虫体,同时提取总RNA并逆转录后,通过荧光定量PCR技术,分析结果发现SjPS siRNA1组的效率与之前筛选实验的结果一致; 同时发现SjPS siRNA1组的雌雄合抱数(10.40±0.51)低于Control组(12.80±0.73)和Mock组(12.80±0.66),差异有统计学意义(F
=4.645,P
<0.05),见图3;虫卵计数的结果显示SjPS siRNA1组的产卵量(1 308.0±75.47)比Control组(2 872.0±101.7)和Mock组(3 043.0±149.7)均下降(F
=71.30,P
<0.001),见图4。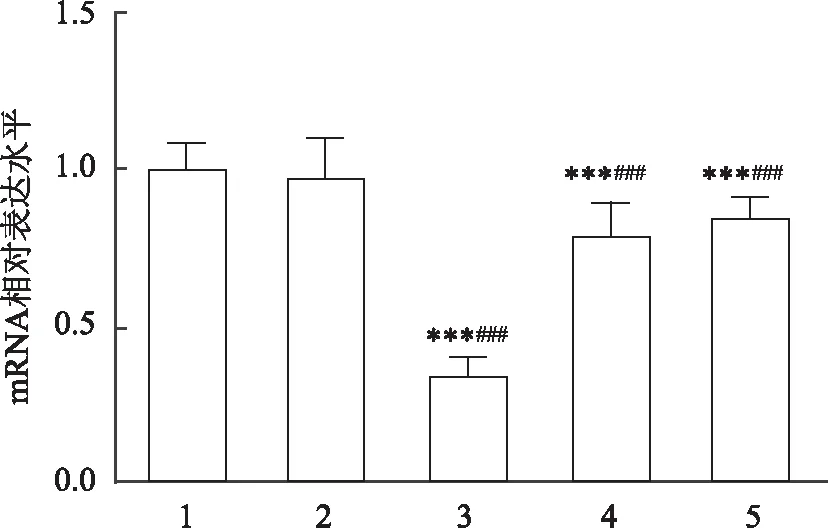
图3 体外筛选干扰效果最好的SjPS siRNAi1:Control组;2:siNCTRL组;3:SjPS siRNA1组;4: SjPS siRNA2组;5: SjPS siRNA3组; 与Control组比较:***P<0.001;与siNCTRL组比较:###P<0.001

图4 SjPS siRNAi1体外干扰对血吸虫产卵量及雌雄合抱的影响A:干扰后早老素基因相对表达水平;B:干扰后雌性成虫产卵数量;C:干扰后雌雄成虫合抱对数;与Control组比较:*P<0.05,***P<0.001;与Mock组比较:#P<0.05,###P<0.001
2.4 RNAi对日本血吸虫生殖系统的影响
形态学分析显示:经过siRNA干扰后的日本血吸虫雌虫与雄虫的生殖系统形态均发生异常。Control组和Mock组均显示雄性生殖系统由数个睾丸组成,且睾丸之间衔接平滑,同时睾丸内含有多个不同发育阶段的雄性生殖细胞,并且比较饱满,形态规则,呈圆形,包括精原细胞和精母细胞。同时睾丸周围的基质结构紧凑。然而,SjPS siRNA1组的血吸虫睾丸形态发生了明显的变化。睾丸的尺寸变小,睾丸间的衔接处出现褶皱,睾丸内精原细胞和精母细胞的数量开始逐渐减少,且形态不规则,细胞间出现较明显的间隙。除此之外,睾丸周围的基质也出现较多的空洞结构。在Control组和Mock组雌虫中,卵巢后部有大的成熟卵母细胞,前部有小的未成熟的卵母细胞。然而,SjPS siRNA1组血吸虫雌虫卵巢内成熟和未成熟卵母细胞界限被打破,卵母细胞分布杂乱,且数量减少,甚至在卵巢内部出现了空洞与裂隙。见图5。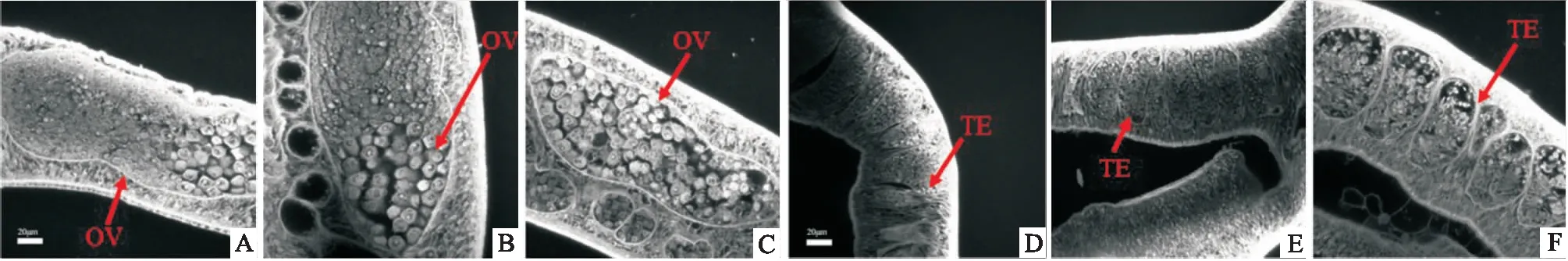
图5 日本血吸虫SjPS被特异性敲降后卵巢及睾丸的形态学变化A~C:雌虫的卵巢;D~F:雄虫的睾丸;A、D:Control组;B、E:Mock组;C、F:SjPS siRNA1组;OV:卵巢;TE:睾丸
3 讨论
在我国主要流行的是日本血吸虫病,其治疗在很长一段时间内只能依靠吡喹酮药物,而长期使用吡喹酮这种单一的药物存在化疗药物抗性的风险。血吸虫为雌雄异体。雌雄合抱、处于繁殖期的日本血吸虫能够持续不断地产生大量的虫卵。虫卵沉积在肝脏和肠系膜静脉,后者经肠黏膜损伤而排出体外。沉积在脏器的虫卵能够形成虫卵肉芽肿,导致纤维化。
昆明鼠感染后16 d,虫体发育成合抱前期,18 d为合抱初期,24 d为合抱后期。在前期实验中,利用抑制性消减杂交技术筛选并鉴定日本血吸虫雌虫在合抱前期、合抱初期和合抱后期的差异表达基因,发现早老素在合抱后期表达量显著升高,推测早老素可能在血吸虫生殖发育中起到一定作用,值得进一步研究。
早老素在种属之间高度保守,可分为早老素-1和早老素-2两种异构体,但早老素在血吸虫中没有异构体。早老素主要通过构成γ-分泌酶在高等哺乳动物体内参与多种有关生殖发育的信号通路,包括与生殖腺卵泡的发育、排卵及黄体化等有关的Wnt通路,以及与胚胎发生和卵母细胞发生在内的多种发育过程有关的Notch通路。有研究表明,针对体外培养的曼氏血吸虫,使用γ-分泌酶制剂DAPT处理后,可以观察到其虫卵发育被抑制,产卵量减少。
本研究证实早老素在血吸虫合抱后期表达量急剧升高,说明雌雄合抱启动了该基因的表达,同时证实该基因在卵巢高表达,提示早老素在血吸虫的生殖发育中,尤其在雌虫的生殖发育中起到重要作用。通过对早老素基因的干扰,早老素被敲降后,抑制了血吸虫的雌雄合抱和产卵量,说明早老素对维持日本血吸虫雌雄合抱和产卵起到重要的调控作用,为进一步研究其调控机制奠定了基础。本研究显示,利用siRNA干扰过日本血吸虫早老素基因表达后,从中挑取培养10 d后仍然合抱的血吸虫成虫,将雌雄虫分离后分别用卡红染色,制作成玻片标本,然后在激光共聚焦显微镜下观察发现,在雄虫中,睾丸内精原细胞和精母细胞的数量显著减少,睾丸的小叶明显小于对照。在雌虫中,卵巢内成熟和未成熟卵母细胞界限被打破,且数量减少。这些结果表明,早老素被特异性抑制后,导致雄性和雌性的生殖细胞分化与增殖受到抑制,而这些细胞的分化与增殖对于成熟生殖细胞的起始和连续产生具有至关重要的作用,在血吸虫体内,若成熟生殖细胞减少,那么卵子的数量也将不可避免地减少,从而使虫卵的发育和产生得到抑制。本研究同时证实RNA干扰组生殖系统相比于空白组雌虫有明显的形态学改变,多数早老素被敲降后的血吸虫睾丸和卵巢中出现空洞与裂隙等异常结构,提示日本血吸虫的生殖系统发育和分化受到了影响,从而抑制生殖细胞的增殖。本研究显示,早老素受到抑制后雌虫的产卵量也明显下降。接下来将进一步研究以早老素为中心的调节生殖系统发育的信号传导途径,将有助于更好地了解日本血吸虫的生殖生物学,发现调控日本血吸虫生殖发育信号通路中的重要分子,为寻找可以抑制血吸虫生殖发育的关键靶点奠定基础。
猜你喜欢
动物医学进展(2024年5期)2024-05-09 08:09:50
热带生物学报(2022年6期)2022-11-29 14:14:44
锦绣·下旬刊(2022年1期)2022-05-16 07:30:45
中国工作犬业(2021年3期)2021-03-27 15:10:41
热带病与寄生虫学(2021年2期)2021-02-28 15:15:11
环境昆虫学报(2020年5期)2020-11-11 08:51:06
中国感染与化疗杂志(2020年1期)2020-01-12 14:07:30
百科知识(2016年13期)2016-08-05 18:23:30
吉林大学学报(医学版)(2015年1期)2015-12-17 07:47:23
中国人兽共患病学报(2015年9期)2015-05-09 01:03:45
