
贵阳某野生动物园芽囊原虫感染调查及基因分型*
2021-11-05 03:00陈礼颖柴宜均姚静新陈善瑜周紫峣刘海峰钟志军彭广能
云南农业大学学报(自然科学) 2021年5期
陈礼颖,柴宜均,姚静新,陈善瑜,周紫峣,刘海峰,钟志军,彭广能
(四川农业大学 动物医学院,四川 成都 611130)
芽囊原虫(Blastocystissp.)是引起肠易激综合征(IBS)、炎症性肠病(IBD)和腹泻等临床症状的重要病原[1-3]。据估计,芽囊原虫在发展中国家对人的感染率高达30%~60%,其中在易与动物接触以及与粪—口传播有关的地区具有更高的感染率[1,4-5]。自首次描述芽囊原虫以来,现已报道芽囊原虫可在多种动物体内被检测到,其中包括野生动物、宠物和家畜等[6-9]。
目前已有关于圈养野生动物的芽囊原虫感染情况的相关研究。胡雄峰[7]调查显示:秦岭地区野生动物感染芽囊原虫中非人灵长类(75.9%)和有袋类(72.7%)感染率较高;乔继英等[10]对陕西地区圈养珍稀野生动物的肠道寄生虫感染情况进行调查,发现芽囊原虫感染率最高(46.7%);LI等[11]研究发现:孟加拉国家动物园内的圈养野生动物的芽囊原虫感染率为15.5%,其中灵长类动物感染率达到31.8%。但目前对于贵阳地区圈养野生动物芽囊原虫感染的研究调查较少。鉴于近期发现野生动物在传播芽囊原虫的过程中扮演重要角色[12]以及可作为带虫宿主对人类健康造成威胁,对贵阳某野生动物园的圈养野生动物进行芽囊原虫感染情况调查,并评估人畜共患风险。
1 材料与方法
1.1 粪便样本采集
收集贵阳某野生动物园野生动物的新鲜粪便样本共203 份,每份样品50~70 g,所采集的动物种类及数量见表1,且均为无腹泻症状的个体。每份粪便单独用无菌塑料密封袋包装,并记录动物种类及编号,低温保存并送至四川农业大学实验室检测。
1.2 粪样基因组DNA 提取与PCR 扩增
用蒸馏水从粪便标本中分离重铬酸钾,3 500 r/min 离心10 min。使用粪便DNA 提取试剂盒(购自Omega 公司)从每个粪便标本中提取200 mg 基因组DNA,于-20 ℃保存。
参照SANTÍN 等[13]的引物序列:上游引物RD5:5′-ATCTGGTTGATCCTGCCAGT-3′,下游引 物BhRDr:5′-GAGCTTTTTAACTGCAACAACG-3′,扩增目的片段长度约为600 bp。以提取的DNA 为模板,PCR 扩增SSUrRNA 基因序列。PCR 反应条件为:94 ℃ 5 min;93 ℃ 1 min;59 ℃ 1 min;72 ℃ 1 min;循环30 次;72 ℃ 3 min。PCR 经琼脂糖凝胶电泳检测所有阳性样品的PCR 扩增产物均交由上海生工生物技术有限公司测序。试验所用的2×TaqPCR Master Mix 购自天根生化科技(北京)有限公司;DL2000 Maker 和ExTaqPCR Master Mix 均购自日本TaKaRa 公司;琼脂糖凝胶购自法国 BLOWEST 公司;Tris-Base和溴化乙啶(EB)均购自万科化学试剂公司;饱和蔗糖溶液由四川农业大学实验室自制。
1.3 芽囊原虫的序列分析与亚型鉴定
将本研究获得的芽囊原虫基因序列经BLAST 比对,在GenBank 下载参考序列,利用Clustal X 2.0 程序(http://www.clustal.org/)对序列进行比较分析后,确定芽囊原虫基因亚型。利用MEGA 7.0 (http://www.megasoftware.net/)软件,采用邻接法(neighbor-joining,NJ)和Boostrap 分析(重复次数为1 000),以蝎虎鞭毛虫(Proteromonas lacertae) (GenBank 登录号:U 37108)作为外群,对PCR 扩增为芽囊原虫阳性的SSU rRNA基因序列构建种系发育进化树。
1.4 统计学分析
采用SPSS 20.0 软件进行数据处理。使用χ2分析芽囊原虫在类群间的感染率差异,P<0.05 表示差异有统计学意义。
2 结果与分析
2.1 SSU rRNA 基因 PCR 扩增结果
PCR 扩增结果显示:在203 份粪便样品中,有 22 份粪便样品成功扩增出大小约 600 bp的SSUrRNA 基因序列的阳性条带,总检出率为10.84%,即该园的芽囊原虫总感染率为10.84%。部分PCR 扩增结果见图1。
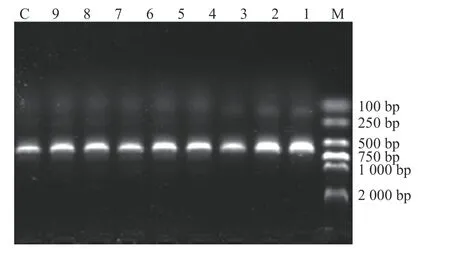
图1 芽囊原虫 SSU rRNA 基因的 PCR 扩增结果Fig.1 PCR amplification of Blastocystis SSU rRNA gene
2.2 芽囊原虫感染情况
由表1 可知:受芽囊原虫感染率的22 份样本包括灵长类(15.19%)、象类(30.77%)和有袋类(35.29%)野生动物;经SPSS 分析,它们之间差异不显著(χ2=0.44,P> 0.05)。在偶蹄类、食肉类和鸟类动物中未发现芽囊原虫。
2.3 芽囊原虫的亚型鉴定及分布
本研究从22 份芽囊原虫PCR 扩增阳性条带中共获得17 份测序成功的SSUrRNA 基因序列,将测序成功的序列与参考序列进行分析比对(图2)发现:5 份基因序列之间具有100%的同源性,并且与参考序列 AB107971 (ST8)位于同一进化支上;8 份基因序列之间的同源性为93.54%~98.44%,并且与参考序列KY929115 (ST1)位于同一进化支上;1 份基因序列与参考序列KX351992(ST2)位于同一进化支上,两者之间同源性为99.64%;2 份基因序列与参考序列MN338086(ST13)位于同一进化支上,两者之间同源性为99.13%;1 份基因序列与参考序列AB070988(ST3)位于同一进化支上,两者之间同源性为98.22%,表明本研究共检测出5种不同的芽囊原虫基因亚型,分别为ST1 (8/17)、ST2 (1/17)、ST3(1/17)、ST8 (5/17)和ST13 (2/17),分布于6种野生动物(表1)。
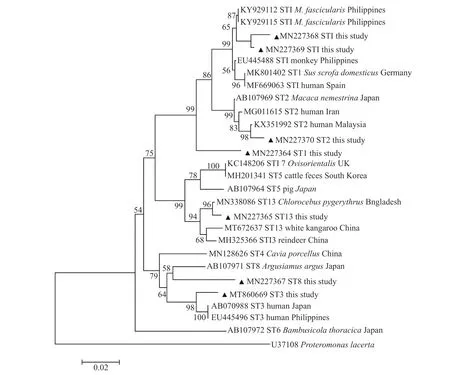
图2 基于芽囊原虫SSU rRNA 基因构建的系统发育树Fig.2 Phylogenetic tree based on SSU rRNA gene of Blastocystis sp.

表1 贵阳某野生动物园芽囊原虫感染情况的检测结果Tab.1 Blastocystis infections of wild animals in a Guiyang wildlife park
3 讨论
本试验将研究对象分为 6 个类群,总感染率为10.84%,其中非人灵长类(15.19%)、有袋类(35.29%)和象类(30.77%)存在感染。研究表明:芽囊原虫感染率在具有感染的动物类群之间差异不大,然而在偶蹄类、食肉类和鸟类动物中未发现芽囊原虫。说明芽囊原虫感染情况在不同动物类群之间存在一定差异,这可能与动物种类采集到的样本数量较少或宿主差异性有关。此外,感染因素可能与感染类群所处地区的饲养管理条件、类群免疫力、卫生管理条件、食用被污染的水源和食物等有关,其中经粪—口途径[4,14]食用受污染的水源或食物是主要影响因素。
在世界范围内,芽囊原虫在各地区的感染率有所差异。调查结果显示:贵阳某野生动物园的总感染率(10.84%)低于秦岭部分野生动物总体感染率(40.2%)[7],但高于中国部分地区人类感染芽囊原虫的总感染率(4%)[15];园区内偶蹄动物并未发现有感染芽囊原虫的迹象,而在陕西部分地区犊牛[1]存在感染该原虫的情况;贵阳该动物园的亚洲象为芽囊原虫阳性,与澳大利亚动物园的亚洲象芽囊原虫[16]感染阳性相似;山魈和骆驼并未发现感染,而秦岭地区的山魈和骆驼分别存在25.0%和50.0%的感染率[7];狒狒和袋鼠在贵阳和秦岭两地都存在感染芽囊原虫的情况;另外,园区袋鼠感染率(35.29%)低于澳大利亚袋鼠感染率(50%)[16]。这些不同国家地区的同种野生动物中芽囊原虫感染情况的差异提示:该原虫的分布除地理因素外,也有可能存在样本的偶然性。同一地区不同动物之间变化较大的感染率也反映了该原虫的宿主特异性。
本研究在贵阳某野生动物园共获得5种亚型(ST1、ST2、ST3、ST8 和ST13),其中ST1(36.36%)是优势亚型,分布于园区内狒狒、长臂猿、黑猴和亚洲象中,表明贵阳园区该亚型的广泛分布。在世界各地也有过多例ST1 感染人的报道[1,4],本研究首次在中国黑猴体内检测出芽囊原虫ST1 亚型,表明黑猴在向人类传播芽囊原虫的过程中可能发挥潜在作用。动物与饲养人员的密切接触可能会增加人患芽囊原虫病的患病率。西澳大利亚动物园和澳大利亚昆士兰州东南部(SEQ)的大型集约化猪场的工作人员芽囊原虫病患病率分别为63%和76.7%[14-16],并且在SEQ 猪场工作人员中,83.3%是芽囊原虫携带者[15]。还有研究表明:法国某动物园的大多数哺乳动物常感染芽囊原虫人畜共患亚型,因此无法确定其传播方向,说明控制芽囊原虫病具有重要的公共卫生学意义[16]。但目前尚未在贵阳该园区发现饲养人员感染芽囊原虫病。ST8 (22.73%)是本研究中感染率第二的亚型,但它只感染袋鼠,分布范围有限。ST13 只分布在金丝猴中,该亚型在澳大利亚的袋鼠中也有报道[17],但在该园区的袋鼠中并未出现ST13,这一结果可能由于亚型具有宿主和地区特异性或所检查的样本数量少所致。长臂猿和狒狒中存在ST1 亚型与其他亚型混合感染的情况,说明该原虫在2种物种间可能存在相互感染。据报道,在芽囊原虫的17种基因亚型中,ST1~ST8 为人畜共患基因亚型[1],提示应加强监测该园区ST1、ST2、ST3 和ST8 芽囊原虫感染人的风险和危害。
4 结论
本研究对贵阳某野生动物园的圈养野生动物芽囊原虫的感染情况进行调查,首次在黑猴中发现芽囊原虫感染,扩大了已知的芽囊原虫宿主范围。本次共发现5 个芽囊原虫基因型(ST1、ST2、ST3、ST8 和ST13),其中人畜共患亚型(ST1、ST2、ST3 和ST8)的发现提示芽囊原虫在该地区具有人畜共患风险。本研究结果可帮助了解动物园动物及其对人类健康的潜在影响具有公共卫生学意义。
猜你喜欢
红领巾·探索(2021年4期)2021-09-14
中国动物传染病学报(2021年3期)2021-07-21
新农业(2020年18期)2021-01-07
知识窗(2020年8期)2020-08-27
知识窗(2020年2期)2020-03-25
快乐作文(1.2年级)(2019年3期)2019-09-10
艺术品(2019年4期)2019-05-30
乡村地理(2018年3期)2018-11-06
艺术品(2018年9期)2018-10-23
小学生导刊(低年级)(2016年8期)2016-09-24
