
经济蟹类性腺脂质需求的研究进展
2021-11-05 03:59张凤英马凌波
动物营养学报 2021年10期
傅 愔 张凤英 赵 明 马凌波
(中国水产科学研究院东海水产研究所,农业部远洋与极地渔业创新重点实验室,上海 200090)
我国甲壳类水产品资源丰富,种类繁多。随着人民生活水平不断提高,水产品在动物性食品中的占有率日趋壮大,而蟹作为传统的水产佳肴一直备受喜爱,其养殖规模亦随之膨胀。性腺发育是蟹类繁殖的基础,良好的性腺发育水平可以提高蟹类的繁殖效率,同时为获得大量优质的苗种提供保证。性腺是蟹类最主要的食用部位之一,性腺发育水平也决定其食用价值。相比未成熟的扣蟹,性成熟的雌蟹和雄蟹滋味更为鲜美,营养更为丰富。蟹类通常在生殖蜕壳后快速发育,根据卵细胞组成与形态的变化,雌蟹第1次卵巢发育分为6个时期(Ⅰ~Ⅵ期),卵巢发育从生殖蜕皮后的Ⅱ期开始,到Ⅴ期基本发育成熟,直至产卵[1-2]。对于雄蟹,精巢发育主要划分为5个时期(Ⅰ~Ⅴ期),从Ⅰ期精原细胞期至Ⅳ期精子期精子成熟。
脂质作为蟹类性腺发育需要的主要营养素之一,直接影响其性腺的发育,关系到后续的交配繁殖、亲本生殖力,甚至是胚胎发育以及内源营养期。此外,脂质作为蟹黄与蟹膏的主要成分,决定了成蟹的营养与食用品质。蟹类性腺中含有丰富的脂质,其中与性腺发育密切相关的脂质涉及各类脂肪酸,特别是二十碳五烯酸(EPA)、二十二碳六烯酸(DHA)、花生四烯酸(ARA)以及磷脂、固醇等。而在实际生产中,脂质的缺乏、过量、不平衡以及氧化均会导致蟹类性腺发育出现一系列的问题,严重制约蟹类养殖业的发展。目前,我国最主要的经济养殖蟹类为中华绒鳌蟹(Eriocheirsinensis)、三疣梭子蟹(Portunustrituberculatus)和拟穴青蟹(Scyllaparamamosain)[3],其养殖产量逐年增高,2019年产量分别达到77.9万、11.4万和16.0万t[4]。养殖过程中脂质的不合理摄入导致亲蟹性腺发育不良、性早熟或者晚熟、肝胰腺白化症、抱卵率下降,甚至流产等问题[5],进而造成成蟹营养价值降低、性腺发育不良、胚胎死亡率高、幼体育成活率低、优质苗种不能满足产业需求等严重后果。因而,研究蟹类繁殖营养、关注性腺发育过程时脂质的调控作用是重点。本文聚焦中华绒螯蟹,同时综述三疣梭子蟹和拟穴青蟹以作对比,就蟹类性腺发育阶段脂质组成及变化规律、脂质在氧化供能与内分泌调控方面的生理作用,以及饲料脂质对性腺发育的影响3个方面作归纳总结,印证蟹类繁殖期科学合理地补充脂质营养对性成熟的促进作用。
1 蟹类性腺发育过程中脂质组成的变化
1.1 蟹类脂质的分类
蟹类中常见的脂质营养素包括不同饱和度、不同链长的脂肪酸,磷脂和固醇等。脂肪酸若依照链长分类,则12个碳以上均为长链脂肪酸。依照饱和度分类,可分为饱和脂肪酸、单不饱和脂肪酸(MUFA)和多不饱和脂肪酸(PUFA),其中PUFA也包含了有3个或以上双键,且碳链长度大于20的高不饱和脂肪酸(HUFA)。典型的长链饱和脂肪酸有棕榈酸、硬脂酸等,MUFA有油酸等,PUFA有亚油酸、亚麻酸、ARA等,而HUFA有EPA和DHA等。此外,根据不同的双键位置,PUFA还能被分为n-3系列和n-6系列,亚麻酸、EPA和DHA均属于n-3系列,而亚油酸、ARA属于n-6系列。类脂中,磷脂是磷酸复合脂,为生物膜的重要组成成分,主要有甘油磷脂与鞘磷脂两类。而环戊烷多氢菲衍生的胆固醇,是蟹类最主要的固醇类化合物。
1.2 蟹类性腺脂质的组成与变化
生殖期蟹类大量合成和积累高价值的脂质,用于构建生殖生理结构。中华绒螯蟹卵巢发育过程中,总脂含量呈先升高后降低的趋势,于Ⅲ期达到最高[6]。而其主要脂肪酸含量也于Ⅲ期达到峰值,但饱和脂肪酸、DHA和磷脂含量于Ⅳ期达到峰值(表1)。拟穴青蟹雌蟹的卵巢发育过程中,磷脂和总脂含量也在Ⅳ期最高,此后降低,而各脂肪酸和固醇含量均在Ⅴ期达到最大值[7-8](表2)。目前公布的数据中,还未有关于三疣梭子蟹不同性腺发育时期各种脂质成分的变化,这是值得今后研究补充的空白。
值得关注的是,完全成熟时,性腺的脂质组成因性别和种类而异(表3)。中华绒螯蟹雄蟹精巢中以饱和的棕榈酸、雌蟹卵巢以单不饱和的油酸较为突出[9-11],以储存为主。各PUFA中,雌性中华绒螯蟹中亚油酸含量较高,雄蟹ARA和EPA含量较高[9]。此外,中华绒螯蟹雄蟹成熟后副性腺异常发达,占据生殖系统的绝大部分,副性腺发育过程中长链PUFA、固醇等脂质的含量在Ⅱ期最高,而它们在性腺中的含量于Ⅳ期最高[12]。
与中华绒螯蟹相比,三疣梭子蟹[9]和拟穴青蟹[13-14]有更高含量的EPA、DHA和PUFA,而青蟹的PUFA比例最高,并且含有大量磷脂[15]。并且,PUFA的n-3/n-6比值在2种海水蟹中亦较高[16]。此外,3种蟹类雄蟹生殖腺中的ARA均高于雌蟹[9,13-14]。
1.3 脂质的转移促进性腺成熟
性腺发育时期,脂质从蟹的肝胰腺转运至性腺,积累以促进性腺成熟。蟹类性腺和肝胰腺中脂质组成及含量随着性腺发育阶段的变化有一定的规律性。肝胰腺脂质在中华绒螯蟹雌蟹性腺发育Ⅲ期达到最大值29.85%,然后开始下降,直至产卵[6]。就卵巢脂质含量而言,中华绒螯蟹卵巢脂质则从Ⅱ期到Ⅳ期一直保持增长,并且在Ⅳ期达到最大值19.05%,然后在产后(Ⅴ期)降到最低值6.59%[6]。远海梭子蟹(Portunuspelagicus)肝胰腺Ⅲ期的EPA含量增高,仅次于饱和脂肪酸[17],而三疣梭子蟹卵巢中的脂质含量也呈现出类似中华绒螯蟹的规律[18]。榄绿青蟹(Scyllaolivacea)卵巢中总脂、饱和脂肪酸、MUFA和PUFA的含量从Ⅰ期增加到Ⅲ期,Ⅵ期呈下降趋势[19]。根据以上试验结果,可以认为在卵巢发育过程中,Ⅲ期雌蟹卵母细胞迅速增长,为内源性卵黄合成期,此时肝胰腺脂质含量也达到高峰。而卵巢的脂质则在完全成熟期,当卵巢体积接近最大时才达到高峰。
性腺成熟与肝胰腺中脂肪酸转运蛋白、脂肪酸结合蛋白、酰基转移酶等的表达有关[20],表明其可能参与了脂质从肝胰腺到性腺的转运。就雄蟹而言,发现脂肪酸结合蛋白在中华绒螯蟹睾丸快速生长中参与了脂质的转运[21]。而在中华绒螯蟹雌蟹卵巢快速生长期脂肪酸结合蛋白也起到了类似的转运作用[22]。此外,中华绒螯蟹卵巢发育过程肝胰腺和卵巢中催化三酰甘油酯合成的甘油-3-磷酸酰基转移酶的表达水平从Ⅰ期开始升高,到Ⅳ期达到最高,然后急剧下降[23]。三疣梭子蟹的肝胰腺中发现至少4种脂肪酸结合蛋白用于介导脂质转运[24]。青蟹肝胰腺转运脂肪时,首先将吸收或存储的脂质转换为氧化稳定性更好、更易消化吸收的磷脂,同时伴随着肝胰腺中的游离脂肪酸逐步增加,及时转运到发育中的卵巢[16]。
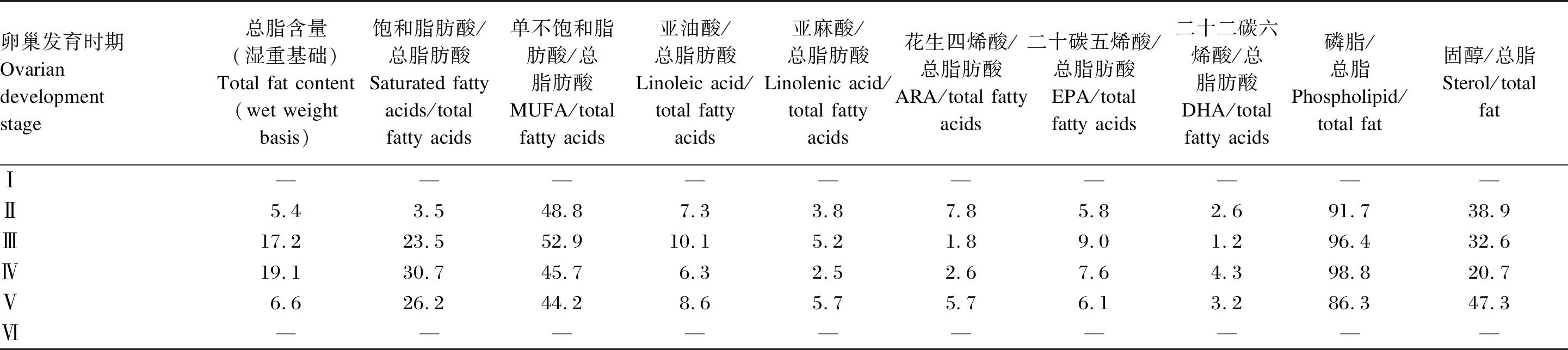
表1 中华绒螯蟹雌蟹卵巢发育过程中各脂质组成的变化
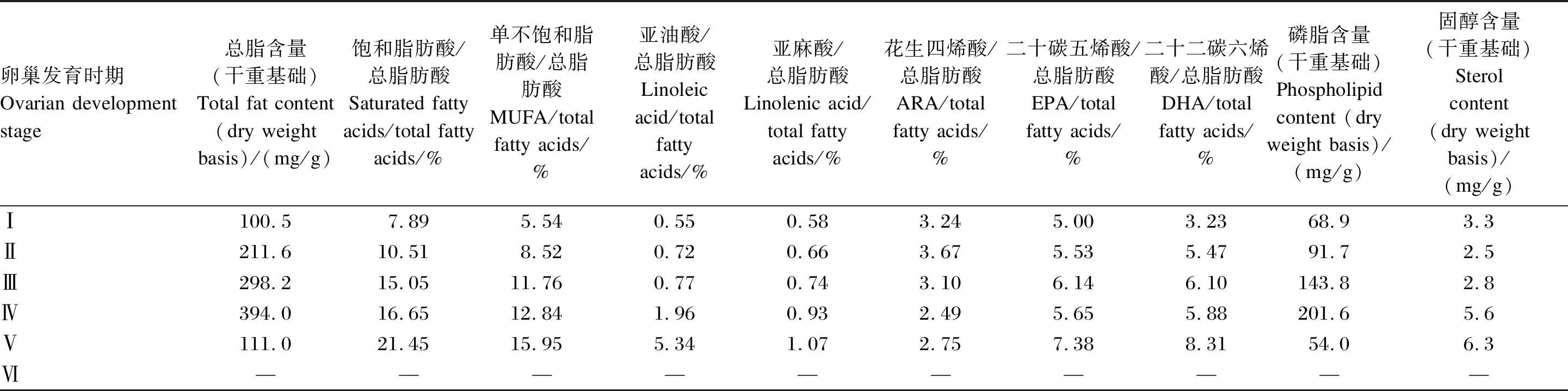
表2 拟穴青蟹雌蟹卵巢发育过程中各脂质组成的变化
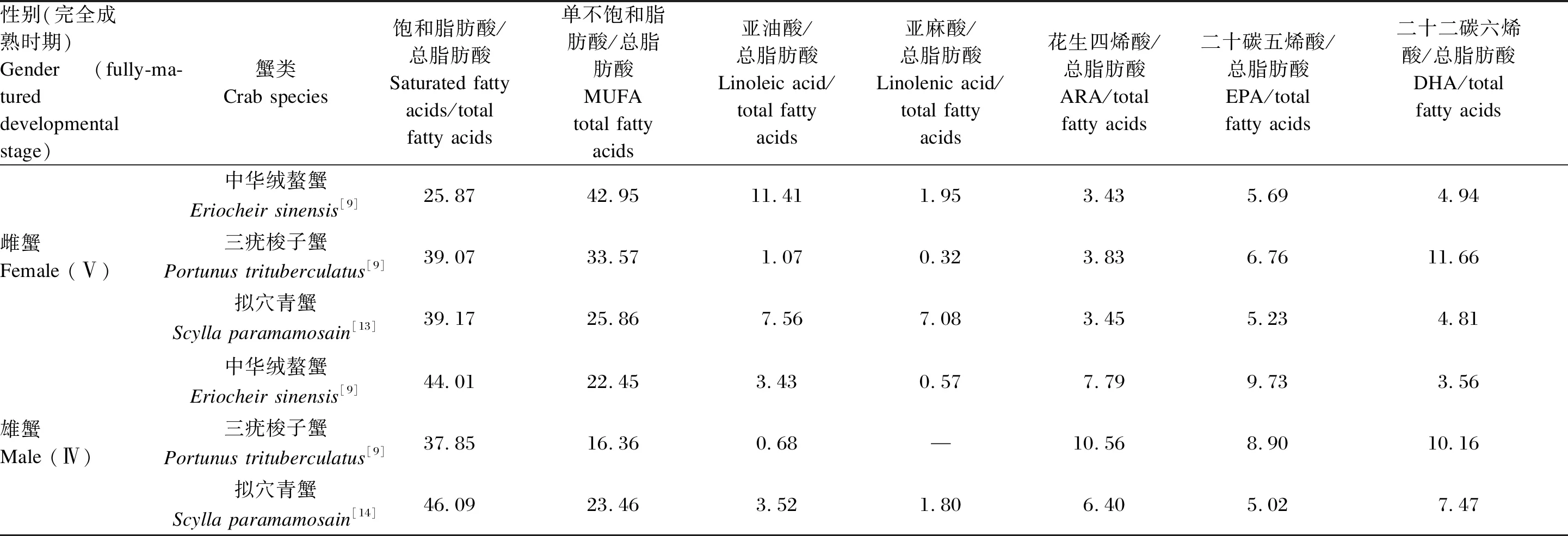
表3 3种经济蟹类的雌、雄蟹成熟性腺脂肪酸的相对含量
参与转运的脂质之中,PUFA是促进性腺发育的关键脂肪酸类别,其大量转运主要集中在中华绒螯蟹雌蟹卵巢发育Ⅲ期后[25]。成永旭等[26]发现抱卵中华绒螯蟹肝胰腺中EPA和DHA含量明显降低,而卵巢中的长链PUFA含量增加[27],这是由于抱卵蟹肝胰腺中高含量的DHA、EPA、ARA等部分转移至卵巢,用于卵巢成熟、受精卵的发育、卵膜卵柄的形成等[10]。研究认为,不抱卵和流产蟹肝胰腺的PUFA未能有效转运至卵巢[28]。在不抱卵的中华绒螯蟹的卵巢中发现EPA和DHA含量较抱卵蟹低,而抱卵蟹流产后肝胰腺PUFA的含量明显上升,其中变化最显著的为DHA与EPA的含量之和,大约上升3.7倍[10],推测其滞留在肝胰腺未能转移至卵巢,使卵巢发育不良,且胚胎得不到足够的营养和能量。
1.4 性腺脂质积累的抗氧化保护
生殖期亲蟹性腺对其自身的抗氧化保护需求要高于其他时期,维生素、矿物质和类胡萝卜素等都对积累的脂质起到一定的抗氧化保护作用。维生素C和维生素E能抑制中华绒螯蟹雌蟹卵子脂质过氧化发生,降低脂质过氧化物丙二醛含量,提高血清超氧化物歧化酶和谷胱甘肽过氧化物酶的活性[29]。维生素C、维生素E的协同作用还可防止细胞和亚细胞膜中磷脂和固醇等不饱和脂肪酸的过氧化反应,使中华绒螯蟹体内自由基达到动态平衡。虾青素能帮助维持中华绒螯蟹[30]卵巢的总脂含量,使之呈上升趋势,这种抗氧化作用同样适用于三疣梭子蟹[31]。矿物质的抗氧化保护主要体现在硒元素上,能激活拟穴青蟹的谷胱甘肽过氧化物酶,保护性腺脂质[32]。此外,β-胡萝卜素在性腺成熟过程中于青蟹卵巢中的沉积也能有效清除自由基,阻止生物膜上含PUFA的脂质和蛋白质氧化[33]。由此可知,适量补充维生素C、维生素E、虾青素、硒、β-胡萝卜素等抗氧化营养素有利于保护亲蟹性腺的脂质营养组成。
2 脂质在性腺发育过程中的生理功能
2.1 脂质氧化为性腺发育供能
脂质的消化分解为蟹类性腺发育提供能量。中华绒螯蟹肝胰腺的脂肪酶活性在卵巢成熟期逐渐上升[34]。三疣梭子蟹肝胰脏中三酰甘油脂肪酶的mRNA表达水平随着卵巢的发育也有所提高[24]。三酰甘油等中性脂肪是主要提供能量的脂质,这点可在大量表达的脂质代谢基因中得到证实。性腺发育期,在三疣梭子蟹肝胰腺转录组中发现与线粒体脂肪酸氧化供能有关的酶基因包括酰基辅酶A脱氢酶、四醇脱氢酶、线粒体样D-β-羟基丁酸脱氢酶和短链脱氢酶等[24]。雄蟹精子能量的主要来源也被认为是脂质的氧化作用[35]。
2.2 脂质参与内分泌调控
生殖期,亲蟹体内的部分脂质充当了性腺发育过程中激素合成的前体物质参与体液调节。比如,胆固醇是性激素、蜕皮激素、肾上腺皮质类固醇激素等生物合成的前体。因此,在卵巢发育阶段,中华绒螯蟹的总胆固醇含量以及肝胰腺中胆固醇酯的含量呈现上升的趋势[20]。再如,ARA通过环氧合酶、前列腺素合成酶转化为前列腺素。前列腺素抑制脂肪的分解,使肝胰腺中脂肪的积累大于分解,并部分转移至卵巢,促进亲蟹卵巢成熟。随着卵巢进一步发育和卵子的产出,抱卵蟹肝胰腺中ARA的含量降低[10],此时脂肪的分解加强,给受精卵提供其余必需的脂肪酸。另外,除了作为前体物质,过量的EPA和DHA还能与三疣梭子蟹前列腺素的中间产物结合[36],抑制其对卵巢发育的生物活性。青蟹卵巢和血淋巴中的固醇类激素含量在卵黄发生初期表现为类似的上升趋势[37],这种上升促进了性激素的合成和性腺的增长与成熟。
此外,脂质本身对生殖内分泌也存在一定的调控作用。脂质可以影响中华绒螯蟹眼柄软组织蜕皮抑制激素(MIH)的基因表达,但不同脂质对其表达的作用相反[25]。当中华绒螯蟹饲料脂肪含量由6%增加到15%时,MIHmRNA的表达明显受到抑制,并且鱼油(n-3 PUFA)也能明显抑制眼柄中MIHmRNA的表达,利于性腺发育。反之,猪油(饱和脂肪酸)和磷脂促进中华绒螯蟹MIHmRNA的表达,磷脂可以提高雌二醇的含量[25]。非基因组的作用机制中,胆固醇能通过长时间孵育作用调控中华绒螯蟹眼柄神经分泌细胞的钙电流,通过对钙离子内流的抑制,影响动作电位的传播,调制神经肽激素分泌[38],影响生殖发育。对于三疣梭子蟹来说,饲料中添加4%的大豆卵磷脂可以提高血清中类固醇激素(孕酮和雌二醇)的含量,并增大卵母细胞直径[39],这可能是类固醇激素生物合成代谢途径基因表达上调所致[40]。再者,磷脂的摄入能降低雌激素硫酸转移酶的表达,提高雌激素活性[40]。
因此,不同类型的脂质可以通过转录水平、神经水平和体液水平等多种途径来调节性激素的分泌,进而影响蟹类性腺的成熟。
3 脂质营养对蟹类性腺发育的影响
蟹类的天然饵料包括鱼、虾、贝、蚬、螺、沙蚕等,种类繁多。但品质不稳定,且受季节影响较大,经常缺乏必需脂肪酸、HUFA、磷脂等,导致亲蟹质量降低。因此,脂质的合理摄入能够改善亲蟹的性腺发育水平。国内外研究表明,亲蟹各组织尤其是肝胰腺、性腺和胚胎的脂肪酸组成与摄入饲料的脂肪酸组成呈正相关[41-43]。饲养试验在揭示摄入脂质与性腺成熟关系的同时,也有助于了解亲蟹对不同脂质的需求量、平衡比例、来源的选择,以及脂质摄入过量和不平衡导致的发育不良、性早熟、肝胰腺白化症等不良后果。一般来说,性腺指数(GSI)和肝胰腺指数(HI)能在一定程度上反映甲壳动物卵巢和肝胰腺中营养积累和发育的状况,因此可以作为评价性腺发育水平的指标。
3.1 脂质摄入量对性腺发育的影响
亲蟹在性腺发育中需要大量磷脂和PUFA,但从头合成的能力有限[44],因此必须从外界摄取。饲养试验揭示了不同雌蟹繁殖期性腺成熟对PUFA和磷脂的需求和适宜添加量(表4)。有关雄蟹繁殖期对PUFA和磷脂的需求量尚未有具体研究。一个有趣的现象是,虽然HUFA是性腺和胚胎正常发育的重要保证,但试验表明中华绒螯蟹亲蟹对DHA、EPA、ARA的需求量并不高,性腺发育只要满足其最低需求量[45],这可能是因为性腺和胚胎中的脂肪酸组成具有较强的保守性。但也有研究认为适量添加DHA和EPA对中华绒螯蟹卵巢发育作用显著[25,41],而且可以显著提高初孵幼体的成活率、变态率和抗逆境能力[46]。
相反,过多摄入脂质,特别是固醇、饱和脂肪酸等,以及生产中投喂动物性饵料过多,则会导致养殖中华绒螯蟹的性早熟。胆固醇在中华绒螯蟹体内积累可直接促进性腺促进激素和法尼酸甲酯及17-β雌二醇、睾酮等甾醇类性激素的合成和分泌,造成性腺发育过快,成熟过早[47]。另外,高蛋白质摄入也可能导致中华绒螯蟹的胆固醇摄入增加[48],诱导肝胰腺和全身的脂质沉积,并且过多的营养转移到性腺导致早熟。此外,含饱和脂肪酸较高的花生油能显著提高中华绒螯蟹类固醇类激素含量,猪油次之,但过量添加也极可能造成过早发育[25]。饱和脂肪酸摄入量超过34%时中华绒螯蟹有早熟现象发生[49]。据此,控制饲料的脂质含量,并适当补充磷脂和HUFA对中华绒螯蟹性早熟存在一定的反作用,能够在转录水平上促进脂肪氧化输出,减轻高脂饲料造成的代谢负担[50-51]。
因此,中华绒螯蟹性腺成熟需要DHA、EPA与ARA的适量添加,对于初始体重75 g以上的雌蟹,推荐的HUFA和磷脂添加量在2%左右。对于初始体重更小的雌蟹,可增加DHA、EPA和磷脂的添加量至6%以上。而饲料中饱和脂肪酸和固醇的添加量应予以控制,避免中华绒螯蟹的早熟和发育不良,影响亲蟹的营养品质及繁殖性能。
与中华绒螯蟹相似,三疣梭子蟹对DHA、EPA、ARA的需求要也不高,仅需满足其最低需求量[36,43,52],并且HUFA含量过高时对梭子蟹有一定的负面影响[52]。ARA的添加量只需在0.06%~0.24%的范围内,过度添加易造成梭子蟹卵巢水分含量过高,阻碍卵膜卵柄发育[36]。
3.2 不同脂质摄入比例对性腺发育的影响
不同脂质与脂肪酸在亲蟹体内的比例平衡十分重要。中华绒螯蟹饲料中添加卵磷脂后饱和脂肪酸含量下降,MUFA、PUFA、HUFA含量的上升有利于卵巢发育[54]。DHA/EPA、n-3/n-6、磷脂/HUFA的配比适当,对促进雌蟹性腺成熟,提高卵子受精、孵化率起着重要作用(表5)。中华绒鳌蟹生长所需DHA/EPA比例为2左右[55]。过高或者过低的DHA/EPA比例都会抑制营养物质的积累,甚至造成肝胰腺白化症。饲料中n-3和n-6 PUFA的动态平衡对性成熟十分重要,单纯追求高营养价值的n-3 PUFA的含量而忽略了n-6 PUFA的含量可能会适得其反。中华绒螯蟹卵巢发育饲料中n-3/n-6比例至少为2时有较好的性腺发育效果[56],磷脂/HUFA的比例处于1.6/2.5~2.3/2.0时也有助于维持中华绒螯蟹体内n-3和n-6 PUFA的平衡[45],促进卵巢发育。

表4 雌蟹繁殖期多不饱和脂肪酸和磷脂添加量的饲养试验结果
与中华绒螯蟹相似,配合饲料中较高的DHA/EPA比例也更有利于三疣梭子蟹肝胰腺营养物质的积累和卵黄物质合成,适宜的比例至少为2[43,52,57]。但HUFA含量过高,油酸、亚油酸和亚麻酸含量过低,则不利于色素在三疣梭子蟹肝胰腺中的沉积,或者是提供能量的饱和脂肪酸和MUFA过少,不能保证梭子蟹肝胰腺正常营养物质的合成及代谢,存在较大氧化压力,甚至会破坏肝胰腺的正常功能[36,43,52]。再者,ARA可能会抑
制n-3 HUFA的合成,并且ARA也可与EPA和DHA竞争与前列腺素的转化酶系统相结合,影响梭子蟹的脂质利用[36]。
由此可知,繁殖期应注意亲蟹饲料DHA/EPA和n-3/n-6比例为2最佳,磷脂/HUFA比例为0.6~1.2,以及MUFA、PUFA、HUFA等的适量补充,利于蟹类性腺发育,避免肝胰腺白化症等问题的发生。

表5 雌蟹繁殖期饲料脂质最适比例的饲养试验结果
3.3 脂质来源对性腺发育的影响
混合植物油替代鱼油可以平衡饲料中的脂肪酸组成,也能够影响亲蟹的性腺发育和繁殖性能。鱼油被植物油替代能够降低脂肪吸收和脂肪酸生物合成,增加β-氧化,并且增加Δ6脂肪酸脱氢酶的表达水平[58]。随着植物油含量的增加,饲料中的亚油酸和亚麻酸含量显著增加,这可能在一定程度上可提高中华绒螯蟹的生殖力[59]。添加鱼油与豆油比例为2∶1和1∶1时,更有利于中华绒螯蟹卵巢发育,比例为1∶2时次之[60]。使用50%~75%植物油替代饲料中鱼油喂养雌性中华绒螯蟹可以适当提高其产卵量[11,60]。因而,在33%~75%替代水平内,对雌蟹卵巢发育都是有益的。对于雄蟹,研究认为混合植物油(豆油∶菜籽油=1∶1)替代不同水平鱼油对中华绒螯蟹雄蟹性腺发育无显著影响[61],其适宜的替代水平为25%~50%。对三疣梭子蟹来说,50%的鱼油替代水平可明显提高卵巢和肌肉中蛋白质含量,而100%的鱼油替代水平会显著降低梭子蟹肝胰腺和卵巢中HUFA的含量[62]。
就磷脂源而言,磷虾油比大豆卵磷脂和蛋黄卵磷脂更有效促进中华绒螯蟹肝胰腺和卵巢的脂质沉积,主要是极性脂质和n-3 PUFA的沉积[54]。此外,磷虾油对中华绒螯蟹卵黄发生的促进作用比其他来源的磷虾油更为明显,其抗氧化效果也最好。
因此,建议选择33%~75%植物油替代鱼油,能够提高亲蟹的性腺发育水平,而磷虾油的抗氧化效果使之成为最有利于不饱和脂肪酸在性腺积累的磷脂源。
4 小结与展望
对蟹类性腺发育各时期脂质组成的研究阐明了脂质在性腺发育期的转移规律,这可能和脂质在性腺发育的氧化供能与内分泌调控方面的生理功能相关。根据脂质摄入量、不同脂质的摄入比例和来源等试验数据,分析脂质摄入失衡的后果,认为满足不同蟹类性腺对脂质的需求,都应适量补充DHA、EPA、ARA、固醇、磷脂等,并且注意配比合理。来源的选择方面,部分植物油替代鱼油是有利于性腺发育的。同时,配合补充维生素C、维生素E、硒、β-胡萝卜素、虾青素等天然抗氧化剂,并且采用抗氧化能力强的磷虾油作为磷脂源,能有效保护性腺脂质的积累。
不过,目前对于蟹类脂质生理作用的理解不够完善,饲料营养的配比还未到最优,因此应从如下方面进一步开展脂质营养调控的研究:1)继续阐明蟹类的性腺发育过程中脂质代谢相关的功能基因,并且开展营养基因组学研究,分析在各种脂质缺乏、适宜和过剩条件下基因表达情况,以期从分子角度为脂质调控性腺发育提供参考依据;2)探索脂质生殖调控作用下可能存在的反馈机制,即固醇类激素是否促进了性腺脂肪的堆积,而积累到一定程度的性腺脂质是否又反过来调控激素的分泌等;3)在传统的干预试验中引入营养遗传学的思路以丰富营养调控蟹类性腺发育的研究,比如不同遗传背景、家系的亲蟹是否对脂质营养有不同需求,应通过特异性的饲料配方有针对地进行饲喂,此外,营养学与遗传学结合也有助于发现蟹类性腺发育中与脂质代谢相关的特殊遗传变异;4)目前的研究仍存在偏重雌蟹的问题,以后应同时关注雄蟹性腺发育的营养生理与需求。
综上所述,蟹类繁殖营养学的研究为高利用率亲蟹营养强化饲料的开发奠定基础,而脂质营养在性腺发育期的平衡摄入能够提高亲蟹的营养价值,优化其繁殖性能,推进经济蟹类苗种产业的高效、健康、稳步发展。
猜你喜欢
文萃报·周五版(2022年24期)2022-06-21
中成药(2019年12期)2020-01-04
中成药(2018年9期)2018-10-09
中成药(2018年7期)2018-08-04
中成药(2018年1期)2018-02-02
中成药(2017年12期)2018-01-19
中成药(2017年5期)2017-06-13
中成药(2017年4期)2017-05-17
食品工业科技(2014年15期)2014-03-11
中成药(2014年9期)2014-02-28
