
奶牛育种中关注的新性状
2021-11-04 00:45张海亮娄文琦陈紫薇米思远王雅春
畜牧兽医学报 2021年10期
张海亮,常 瑶,娄文琦,王 凯,陈紫薇,米思远,温 万,王雅春*
(1.中国农业大学动物科技学院 农业农村部动物遗传育种与繁殖(家畜)重点实验室 畜禽育种国家工程实验室,北京 100193; 2. 宁夏回族自治区畜牧工作站,银川 750004)
自上世纪20年代起,奶业发达国家的奶牛育种目标逐渐开始形成。最初,各国的奶牛育种家通过测奶计划收集产奶量和乳脂量数据对奶牛进行选种,同时考虑奶牛的体型外貌。此后,很长一段时间内,奶牛选育目标仅包括产奶和体型外貌性状;期间,体型外貌性状的权重不断增加,并于1937年总体上超过20%。产奶和体型性状的高强度选择对奶牛的健康和繁殖性能间接造成了负面影响,育种家们也逐渐意识到了这个问题,逐步开始重视并研究这些功能性状。上世纪90年代后,许多国家将繁殖、产犊和健康等性状加入选择指数中,在强调高产的同时,不断重视奶牛的健康和长寿性,产量和体型性状的权重逐步降低,重要功能性状的权重不断提高,平衡育种的理念在奶牛育种中逐渐形成[1]。随着时间的推移,各国奶牛的选育目标在性状组成和权重占比等方面不断变化。目前,奶业发达国家的选择指数中常见的性状可以分为6类,包括产奶性状、体型外貌、长寿性、产犊性状、繁殖性状和健康性状。遗传选择对这些性状的提高显而易见,自从这些性状被纳入选择指数之中,已经在牛群中产生了巨大的遗传进展,尤其是当基因组选择技术开始应用以后[2]。
随着行业的不断发展,人们对奶牛的认识和理解更加深入,消费者对产品提出了更多要求;同时,人们对气候变化和环境保护的意识不断增强,奶牛养殖可持续发展的要求越来越高。因此,从遗传育种层面也对奶牛提出了新的要求,许多新性状进入各国研究人员的视野,并逐步开始在奶牛的遗传选育中应用。此外,随着牧场规模化水平的提高,各类传感器、摄像头等现代化技术在牧场中广泛应用,为奶牛遗传选育提供了规模更大、信息更丰富的个体记录,这些个体记录为新性状的开发奠定了数据基础。基因组选择技术的应用,为低遗传力、表型测定困难的性状提供了有效的选择工具,有助于新性状在育种实践中快速应用。
为了确保奶牛选育持续的进展,并使育种目标始终符合生产者及消费者的期望,新性状的研究和应用势在必行。随着产业的发展,奶牛选育中考虑的性状仍在持续变化,在世界范围内,许多新的性状正在被考虑并处在不同的发展阶段。本文收集整理了近10年来各国奶牛遗传选育中正在研究和应用的新性状,并将这些性状分为生产效率相关的新性状、应对环境挑战的新性状、健康福利相关的新性状、产品和加工相关的新性状、管理相关的新性状五大类,详细介绍了这些性状的选育背景、定义方法、遗传基础和选育应用情况等,以期能够为我国奶牛育种提供参考。
1 生产效率相关的新性状
1.1 饲料效率(feed efficiency)
奶牛养殖中,饲料成本是奶牛养殖总成本中最大的组成部分。提高奶牛的饲料效率,不仅能够提高牧场的养殖效益,改善产品的竞争力;世界范围内,提高饲料效率对解决人口增加带来的粮食安全问题、人畜争粮矛盾更是具有深远意义。通常,饲料效率指单位饲料投入带来的单位产品产出,包括产品的质量、经济价值、能量或蛋白等[3]。在世界范围内,遗传选择能够提高奶牛的饲料效率已经成为了共识。然而,由于表型测定困难、性状定义复杂等原因,饲料效率的选育应用经历了较长时间[4-5]。
由于直接对饲料效率进行度量的难度较大,研究人员提出了许多用于选择饲料效率的指示性状,例如剩余采食量(residual feed intake, RFI)、剩余干物质产量(residual solids production, RSP)和牛奶中红外光谱等[6-7]。其中,RFI是研究最成熟、应用最广泛的定义方法。RFI可以通过线性回归的方法进行估计,其代表了奶牛实际的干物质采食量(实际测定获得)与预期的干物质采食量(基于体重、产奶水平等)之间的差异,RFI越小代表奶牛的饲料效率越高;此外,RFI还可以考虑奶牛的生产性能水平、代谢体重、体况评分和泌乳阶段等因素,RFI被认为代表了决定效率的基础代谢过程的固有差异[8-9]。研究指出,饲料效率是一个中等遗传力性状,不同奶牛群体中RFI定义的饲料效率性状遗传力在0.01~0.40之间[8]。
在奶牛饲料效率的选育中,基因组选择已经成为了不可或缺的工具。此外,由于泌乳牛的采食量数据测定更加困难,整合青年母牛的饲料效率数据提升奶牛在全生命阶段的饲料效率也是一个值得探索的角度[10]。近年来,在美国(TPI指数)、澳大利亚(BPI指数)、荷兰(NVI指数)等国家的奶牛选择指数中,已经考虑了饲料效率性状,其权重在5%~8%之间。对饲料效率的研究和选育中,同时积累奶牛的采食量记录、体重记录、生产性能记录十分必要。
1.2 繁殖技术性状
为提高优秀母牛遗传物质的利用效率,超数排卵、体外受精和胚胎移植等繁殖技术在全世界广泛应用。随着数据的不断积累,育种家对繁殖技术相关的性状也进行了一些遗传学研究。例如,在德国荷斯坦牛中[11],研究人员对母牛冲卵数、可移植胚胎数、退化胚胎数、未受精卵子数和胚胎移植率等进行了遗传分析,结果发现,这些性状具有低到中等的遗传力(0.14~0.23);在加拿大荷斯坦牛[12]中,每次冲卵获得的胚胎总数和可成活胚胎数为中等遗传力性状(0.14~0.17)。总体上,繁殖技术相关的新性状为中等遗传力性状,具有选育提高的潜力。这些性状间一般存在较高的遗传相关,相关系数一般在0.5~0.9之间;与产奶性状之间存在较低的遗传相关,相关系数通常在-0.1~-0.3之间;与繁殖等功能性状一般也存在较低的遗传相关,遗传相关系数在0.1~0.3。由于繁殖技术的应用中涉及的动物角色更多,根据不同的性状,对这些繁殖技术新性状进行遗传分析的模型将更加复杂,通常需要考虑供体母牛和与配公牛的加性遗传效应,以及供体母牛、与配公牛和受体母牛的永久环境效应等。此外,部分研究也鉴定了一些与繁殖技术性状相关的基因组分子标记[13]。
1.3 公牛繁殖力性状(male fertility)
根据牛的繁殖过程和公母牛各自的生理特点,可将奶牛的繁殖性状分为3类。第一类为公牛直接表现的性状,如精液品质和精液产量等,这些性状可利用公牛自身的表型对种公牛进行评估,为第一类公牛繁殖力;第二类为仅受母牛影响的繁殖性状,如首次配种日龄和产后首次配种天数等,这类性状可利用母牛自身的表型对母牛及母牛父亲进行遗传水平的估计,这一类繁殖性状与公牛繁殖力无关;第三类为母牛及与配公牛共同影响的繁殖性状,如配妊次数和56 d不返情率等,这类性状可利用母牛自身表型对母牛及母牛父亲进行遗传水平的估计,同时还可以估计与配公牛的遗传影响,与配公牛对这类繁殖性状的影响为第二类公牛繁殖力[14]。公牛繁殖力受公牛与配能力的遗传水平和精液性状的遗传水平共同影响,公牛繁殖力与母牛繁殖力性状之间仅存在较低的遗传相关。尽管公牛繁殖力为低遗传力性状,对公牛繁殖力的选育仍可以进一步提高牛群的繁殖效率[15-16]。
2 应对环境挑战的新性状
2.1 甲烷排放量(methane emission)
由于甲烷与温室效应强烈相关,所以减少反刍动物的甲烷排放量已经成为了目前动物科学领域的研究热点。通过比较不同个体、品种及生命阶段的甲烷排放量差异,发现通过遗传选择可以减少个体的甲烷排放量[17]。测定奶牛的甲烷排放,需要投入额外的设备和人力,且数据的准确性无法保证,收集甲烷排放量的大规模个体记录目前仍具有一定困难。研究指出[18],采食量、采食行为、瘤胃功能、代谢物、微生物、产奶量、乳成分和粪便排放等都可以尝试用于对甲烷排放量进行间接选择;例如,RFI与甲烷排放量之间存在一定相关,高饲料效率的奶牛,其甲烷排放量也更低[17]。
在荷兰荷斯坦牛[19]中,研究人员使用体重、采食量和产奶量数据预测了奶牛的甲烷排放量,并使用随机回归模型对预测的甲烷排放量进行了遗传分析,发现不同泌乳阶段下,日甲烷排放量的遗传力在0.29~0.42之间;不同泌乳阶段下日甲烷排放量的遗传相关系数的变异较大(0.31~0.98),相邻泌乳阶段之间的遗传相关更高;此外,每单位乳脂乳蛋白校正乳的甲烷排放量性状的遗传力更高,遗传力介于0.39~0.65。在意大利的瑞士褐牛群体[20]中,研究人员使用牛奶脂肪酸的中红外光谱数据预测甲烷排放量,发现该性状同样为中高遗传力性状(0.16~0.35)。在丹麦荷斯坦牛[21]中,通过自动挤奶机器人上增加的自动采样和分析单元直接测定了奶牛甲烷排放量的个体数据,并对甲烷排放量性状进行了遗传分析,其遗传力在0.16~0.21之间。在甲烷排放量性状的定义中,排放量的度量和计算基础也存在许多选择,常见的研究中通常以浓度、单位时间内的总排放量或单位产品(例如,每千克牛奶、乳蛋白、乳脂等)的排放量为基础量化奶牛的甲烷排放量差异。此外,由于甲烷排放量性状不能直接产生经济效益,将其纳入选择指数时,确定该性状的经济权重也存在一定困难。
2.2 热应激抗性(heat stress)
随着全世界范围内气候环境的变化,在不久的将来,选育更加耐热的奶牛将变得非常重要。Ravagnolo和Misztal[22]认为,对热应激耐受性状的选育是可行的,尤其在平均温湿度指数较高的地区非常有效;在研究热应激耐受性状的遗传规律时,积累气象资料十分必要[23]。在热应激耐受性状的研究中,如何定义热应激耐受性状,一直是该性状研究中的难点。在研究中,奶牛的生理指标(如直肠温度、呼吸频率、流涎评分等)、产奶量随环境温湿度的变化幅度等都被用作热应激耐受性状进行研究。例如,在美国荷斯坦牛中[24],热应激条件下直肠温度的遗传力为0.17,且直肠温度与产奶量、乳脂量、乳蛋白量、体细胞评分、女儿怀孕率和生产寿命等存在较低的遗传相关(-0.013~0.096)。澳大利亚将热应激耐受性状定义为热应激状态下产奶量随温湿度指数增加而下降的幅度;在其荷斯坦牛群中,以产奶量、乳脂量、乳蛋白量降幅而量化的热应激耐受性状的遗传力分别为0.22、0.20和0.23;在娟姗牛群中,该性状的遗传力更高,分别为0.33、0.26和0.27;此外,热应激耐受性状在不同胎次之间存在较高的遗传相关,两个品种中均高于0.9;对该性状进行基因组选择时,在荷斯坦牛中的准确性处于0.48~0.50之间,在娟姗牛中的准确性处于0.44~0.61之间。2017年,澳大利亚开始发布热应激耐受性状的育种值,并对该性状进行选育。
3 健康与福利相关的新性状
3.1 免疫反应(immune response)
对奶牛养殖产业来说,奶牛对感染和疾病的免疫反应能力是一个具有经济意义的复杂性状。通过研究牛血清免疫球蛋白含量的遗传参数,发现遗传选择可以改变免疫球蛋白的含量,最终达到减少疾病的目的[25-26]。在加拿大荷斯坦牛中[27],研究人员分别对由细胞介导和抗体介导的免疫反应进行了遗传分析,这些性状的遗传力在0.16~0.41之间。通过血液中T细胞、B细胞、NK细胞和粒细胞的含量,发现这些细胞免疫相关的性状为中高遗传力性状(0.15~0.46);此外,通过研究这些性状与产奶、繁殖和健康等之间的遗传关系,发现对细胞免疫相关性状的遗传选择在提高动物健康、适应性和繁殖力中具有较大意义[28]。一些奶牛育种公司通过筛选具有高免疫反应的种公牛,以提高公牛女儿的疾病抗性,这类高免疫反应种公牛的遗传物质已经在一些国家的牛群中广泛应用。根据我国牛群中的研究,高免疫反应水平的公牛,其女儿对部分疾病的抗性高于普通公牛的女儿[29]。
3.2 健康性状(health traits)
3.2.1 肢蹄病 研究表明,跛足对奶牛的产奶量和长寿性均有影响,肢蹄健康对奶牛业非常重要[30-31]。在各国奶牛育种体系中,体型外貌性状中大多包含线性肢蹄评分性状,对包括肢蹄在内的线性体型性状开展遗传评估已有多年。然而,一些研究[32-33]发现,肢蹄健康性状和线性肢蹄评分之间的遗传相关较低,这在一定程度上说明了通过选择体型性状来提高肢蹄健康的效率较低,也指出了将肢蹄健康性状直接纳入选育目标的必要性。肢蹄健康性状一般根据奶牛的发病状态定义为二分类或多分类阈性状,可使用线性模型和广义线性模型进行遗传分析。奶牛肢蹄健康性状是一个低遗传力性状,其估计遗传力在0.01~0.03之间[33-35];对肢蹄健康性状进行选育时,需要积累大量的肢蹄疾病诊疗记录。此外,基于日活动量可以尝试对肢蹄健康性状进行预测,利用奶牛的行为变化开发肢蹄健康性状的自动化监测系统将会是奶牛场管理的有用工具,并为选育肢蹄健康性状提供新的思路[36]。在荷兰(NVI指数)和北欧(NTM指数)的奶牛选择指数中,均考虑了肢蹄健康性状,在选择指数中该性状占3%~7%的权重。
3.2.2 临床乳房炎 奶业发达国家很早就开始了对乳房健康相关性状的遗传选择,早期主要将牛奶体细胞数、乳房深度和前乳房附着等作为乳房健康的指示性状,对临床乳房炎抗性进行间接选择。然而,这些乳房健康的指示性状并不能完全解释临床乳房炎抗性的遗传方差[37]。因此,各国相继建立临床乳房炎的表型数据采集系统对临床乳房炎进行直接选择。一般根据奶牛泌乳期内是否发生临床乳房炎,定义为二分类阈性状,采用线性模型和广义线性模型对其进行遗传评估。线性模型估计的遗传力范围为0.01~0.03[38-39],广义线性模型遗传力估计结果(0.06~0.10)高于线性模型[37, 40-42]。目前,一些国家已将临床乳房炎抗性加入了选择指数中。例如,在美国NM$指数、加拿大LPI指数和法国ISU指数中,临床乳房炎抗性的权重分别为0.07%、6.6%和7.2%。
3.2.3 其他健康性状 影响奶牛生产效益和动物福利的健康问题众多,且具有不同发病机制的健康性状之间存在较低的遗传相关[39]。对奶牛疾病抗性进行选育提高时,除肢蹄病和乳房炎之外,其它健康性状的选择也很重要。表型数据是影响健康性状遗传评估的主要因素,国外很早就开始探索健康性状的直接选择,并建立了专门的数据收集体系,对健康性状进行了相关研究,有较多关于健康性状遗传参数的报道。相比生产性状(>30%),健康性状的遗传力(<5%)一般都较低[43]。目前,美国HTH(health trait subindex)指数中包括6个健康性状,分别是乳房炎、子宫炎、酮病、真胃变位、产乳热和胎衣不下,HTH在NM$中的权重为2.33%[38]。由于一些健康性状缺乏表型数据,研究中也常将某类健康问题整体考虑,根据特定时间是否至少发生一次该类健康事件,将某类健康问题的整体抗性定义为性状。例如,澳大利亚奶牛群体中,将子宫炎、胎衣不下、卵巢囊肿和子宫炎等整合为繁殖障碍类健康性状,其估计遗传力为0.01[39]。
3.3 后备牛存活力
上世纪70年代起,奶牛育种就开始关注成母牛淘汰造成的经济影响,并定义了各种长寿性状以选育成母牛抵抗淘汰的能力[44];然而,后备牛的淘汰也占据了不小比例,奶牛育种缺乏对后备牛淘汰应有的重视[45]。近年来,各国开始对后备牛淘汰从个体层面进行量化,定义出一系列的后备牛存活力性状,并对后备牛存活力性状进行遗传分析,考虑通过遗传的手段改善后备牛存活能力。
后备牛存活力性状通常定义为某一阶段内(或某一时间观察点)个体的存活状态(0或1),该性状通常为二分类阈性状。不同国家牛群的研究中,通常将犊牛出生后第30、60、180、365、450 天和头胎产犊作为分界点,对后备牛各阶段的存活力进行分阶段定义[46]。在不同群体中,后备牛存活力性状是一类低遗传力性状,使用线性模型估计的遗传力多在0.01~0.10之间[47-49]。目前,北欧已将后备牛存活力性状纳入选择指数(NTM指数)之中,其权重为4%;德国也开始对后备牛存活力性状进行常规遗传评估。在美国荷斯坦牛[50]和娟姗牛[51]中,后备牛存活力性状基因组选择的可靠性分别可达47.3%和50.5%。
4 产品和加工相关的新性状
4.1 傅里叶中红外光谱(Fourier-transform mid-infrared, FT-MIR)
傅里叶中红外(FT-MIR)光谱在乳品行业发挥重要作用。牛奶FT-MIR光谱蕴含牛奶的全部信息,利用光谱中波数信息可以对直接(牛奶组分)和间接(疾病、机体能量水平等)表型进行预测,借助常规DHI检测就能快速获取奶牛的疾病、能量水平和繁殖状态等信息,不增加额外的数据收集成本,这无疑对奶牛育种和整个行业的发展具有极大推动作用[52]。据前人研究,FT-MIR光谱预测的脂肪酸遗传力在0.07~0.51之间[53],乳蛋白率遗传力为0.36[54],乳铁蛋白遗传力在0.20~0.22之间[55],预测的矿物元素如钙、磷遗传力在0.10~0.26之间[56],预测的乳脂滴体积及表面积遗传力分别为0.30和0.24[57],这些中高遗传力的新性状可以在未来的奶牛育种中加以考虑。此外,FT-MIR光谱自身也是一条具有中高遗传力的曲线,借助FT-MIR光谱可深入挖掘牛奶中的很多未知信息。
4.2 乳成分
由于牛奶的成分对其营养价值和加工性能均存在影响,各种牛奶组成成分的遗传变异备受关注。研究表明,牛奶脂肪酸含量受遗传控制。随着不同脂肪酸碳链长度的增加,不同脂肪酸含量的遗传力趋于降低[58]。此外,饱和脂肪酸含量的估计遗传力高于不饱和脂肪酸[59]。因此,通过遗传选择可以改变牛奶中的脂肪酸含量,以及不同脂肪酸的比例。一些国家已经开始在奶牛育种规划中考虑脂肪酸含量性状,如比利时等。
牛奶中的矿物质,如Se、Ca、K、Zn、Mg和P等参与人体的重要生理过程。在荷兰荷斯坦牛中[60],研究发现不同奶牛和不同牛群之间牛奶矿物质浓度具有较大差异,这些牛奶矿物质浓度的遗传力较高(0.20~0.62),可以通过营养或遗传的手段改变。
4.3 牛奶凝固性能
由于奶酪行业的快速发展,牛奶凝固性性状逐渐被关注。研究发现,不同品种奶牛所产的牛奶,其牛奶凝固性能不同,牛奶凝固性能受遗传因素的影响;例如,荷斯坦牛和北欧奶牛品种的牛奶凝固性能较差,而瑞士褐牛和西门塔尔牛等品种的牛奶凝固性能更好。在芬兰爱尔夏牛中[61],凝乳时间性状的遗传力最早被估计,该性状为中等遗传力性状(0.23~0.27)。此后,奶酪加工过程中重要的牛奶物理特性,如凝固时间、加入凝乳酶30 min后的凝固状态、凝乳时间等性状被广泛研究[62]。然而,牛奶凝固性能的表型收集具有时限性,性状测定费用昂贵,这些因素限制了牛奶凝固性能的选育应用。随着牛奶检测设备的发展,一些研究通过FT-MIR光谱预测牛奶的凝固性能。研究发现[63],FT-MIR光谱的预测值和牛奶凝固性能的实测值之间的遗传相关可达0.7~0.9,且FT-MIR光谱预测的牛奶凝固性能具有较高的遗传力(0.22~0.34),FT-MIR光谱预测值可以作为选育牛奶凝固性能的指示性状。
5 管理相关的新性状
5.1 性情(temperament)
在奶牛众多的行为中,性情最受研究人员青睐,也是为数不多纳入奶牛选择指数的行为性状之一。性情指奶牛在面对紧张性刺激或不利外界环境时所表现出一贯的行为方式和生理上的差异[64],在各国奶牛群体中,研究人员通过奶牛挤奶时的行为表现对奶牛性情进行评分;通过行为学实验,也可以评价奶牛性情。研究表明,奶牛性情受年龄[65]、饲养模式[64, 66]、品种[67]和性别[65]等因素影响。国际上,对奶牛性情的遗传学研究开展较早,奶牛性情为中低遗传力性状(0~0.67)[68-71],长期选育使牛群中温顺奶牛的比例持续增加。例如,加拿大牛群的遗传评估显示,拥有产奶记录的奶牛中,超过90%奶牛的性情可以被评价为中等、温顺和非常温顺[72]。自2009年以来,加拿大、德国、荷兰、瑞士、挪威、瑞典和丹麦等国家对性情性状进行育种值估计。其中,北欧已将性情性状纳入了综合育种指数(NTM指数),在娟姗牛、红荷斯坦牛、荷斯坦牛和北欧红牛4个品种中,性情的选择权重分别为0.02、0.03、0.04和0.10[73]。此外,相关研究已经进一步证明了行为性状的基因倾向性,同时证实了不同基因组区域影响特定行为性状的假设[74-77]。在各奶牛群体中,在2、3、5、18、23和29号染色体上均发现了一些与性情性状相关的QTL[75,77-78]。
5.2 挤奶速度(milking speed)
挤奶速度是一个重要的管理性状,该性状代表了单位牛奶产出所需要的挤奶时间。挤奶机器人可以对奶牛的挤奶速度进行量化,但由于挤奶机器人并没有在各国牛群中广泛应用,这类数据的收集十分有限。目前,加拿大、美国和法国等通过主观评分,收集挤奶速度性状的表型记录,对该性状进行评估选育。其中,美国采用8分制[79],加拿大[80-81]和法国[82]采用5分制。研究发现,挤奶速度是一个中高遗传力性状(0.10~0.37);在美国的荷斯坦牛和瑞士褐牛、加拿大的荷斯坦牛、法国的荷斯坦牛群体,以及丹麦的红牛、荷斯坦牛和娟姗牛等群体中[79-82],挤奶速度被广泛研究。在美国、加拿大、北欧和法国等地,已经开始对挤奶速度进行评估,甚至纳入了选择指数。例如,在北欧荷斯坦牛的NTM指数中,该性状的权重为3.3%。此外,一些研究通过全基因组关联分析定位了一些与挤奶速度相关的遗传标记[82-83]。
6 总结与展望
回顾我国奶牛的选育目标,20世纪80年代初,我国借用美国的奶牛总性能指数(total performance index, TPI)计算公式将产奶量、乳脂率及体型外貌3类性状的育种值合并为TPI 指数,并按测定公牛的指数进行种公牛排队选留,由此实现了公牛遗传评定。2007年,中国奶业协会发布了中国奶牛性能指数(China Performance Index, CPI),指数中包含产奶量、乳脂率、乳蛋白率、体细胞评分、体型总分、泌乳系统和肢蹄系统等性状;2012年,随着基因组选择技术在我国奶牛育种中的应用,中国奶业协会发布GCPI(Genomic China Performance Index)指数;2020年,中国奶业协会对CPI指数进行了修订,将产奶量、乳脂率和乳蛋白率3个性状修订为乳脂量和乳蛋白量性状。目前,我国奶牛选育目标中仅包含产量性状、体型性状和体细胞评分,对繁殖性状、长寿性状和健康性状的遗传评估正在研究和小范围内应用;不久的将来,这些重要的功能性状也会纳入CPI指数中。
如图1所示,任何一个新性状的选育应用均会逐步经历不同阶段,最终在育种实践中被成熟应用。通常,为了解决行业或牛群的某个新需求,研究人员将尝试定义一个新的性状,并试验性地收集一些该性状的表型数据,通过研究逐步建立适合该性状的评估模型和分析方法,获得该性状的遗传参数及其与其他性状的遗传关系等;通过经济重要性分析,确定性状在选择指数中应有的权重;通过收集更大规模的数据,逐步对该性状开展大规模评估和选育;随着该性状选育的持续进行,可以观察其选择效果,并进一步对该性状的评估、选育等进行调整。可见,任何一个性状的选育都不是一蹴而就的,数据积累和科学研究的过程需要一定时间,同时育种工作又要求具有一定的前瞻性。因此,总结奶业发达国家的性状选育经验,并在我国牛群内进行相关的研究和数据积累十分必要。
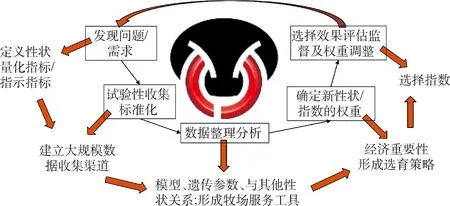
图1 奶牛育种中新性状的研究应用过程Fig.1 The technical processes of research and breeding application for novel traits in dairy breeding
我国奶业正随着社会的进步和行业的发展而快速进步,养殖与环境矛盾、动物福利问题同样给我国奶牛养殖业带来了压力,乳制品消费行为的变化对牛奶各项性能提出了新的要求,自动记录设备和各类传感器在牧场中的大规模应用给奶牛遗传选育提供了新的可能。这些挑战和机遇都提醒我们,一些对整个产业发展有重要意义的新性状纳入遗传评估是奶牛育种行业发展的必然趋势。不久的将来,我国奶牛选育指数所包含的性状将更加丰富,奶牛选育目标将更加健全。在育种实践和研究中,我们应该注重对各类数据的积累和新性状的开发,并在基因组选择参考群的建立中留出空间,以应对未来随着行业发展奶牛种业所面临的新挑战,最终建立健全我国自主创新的奶牛育种体系。
猜你喜欢
贵州畜牧兽医(2022年6期)2022-12-29
养猪(2022年4期)2022-08-17
文苑(2018年20期)2018-11-15
学生天地(2018年30期)2018-10-17
少年漫画(艺术创想)(2018年5期)2018-09-11
湖北畜牧兽医(2015年11期)2016-01-11
湖北畜牧兽医(2015年11期)2016-01-11
华南农业大学学报(2015年5期)2015-12-04
畜牧兽医学报(2015年3期)2015-07-05
山东农业科学(2014年1期)2015-03-09
