
间伐强度对光皮桦天然次生林天然更新及植物多样性的影响1)
2021-11-03 12:12:30汪娅琴王德炉吴丽丽
东北林业大学学报 2021年9期
汪娅琴 王德炉 吴丽丽
(贵州大学,贵阳,550025) (毕节市七星关区林业局)
林下植被作为森林生态系统一个重要组分。林下植被由林下灌木、藤本、乔木幼苗及草本组成,其组成与结构直接或间接影响着森林生态系统多样性、群落功能特征和演替过程、林地生产力维持和养分供给等[1-3]。群落物种多样性的高低是衡量森林质量变化的指标和确定科学抚育措施的依据[4-5]。马履一等[6]对油松(Pinustabulaeformis)人工林抚育间伐后,发现林下物种多样性短期内显著提高,6 a后反而有所下降。抚育间伐对林下物种多样性的影响尚未定论,林下植物多样性随间伐强度、频度、演替发展阶段及环境因子变化而发生相应的分异[7-8]。间伐对林下天然更新有重要影响,间伐后林下更新幼苗的种类和空间分布格局变化对植被恢复、种群的生存与繁衍、森林群落结构和演替方向具有一定的指示作用[9-10]。幼苗更新成功与否直接影响或制约着群落物种组成和生态系统平衡与稳定。米爽等[11]发现针阔混交林幼苗更新数量在轻度采伐和重度采伐下达到最大,中度间伐下最小。抚育间伐作为森林可持续经营的重要营林措施,科学的抚育措施能够优化林分结构[12]、调节林内光温水热资源再分配[13]、改善土壤交换性能[14]、扩大林下生态位空间[15-16]、增加林下小生境异质性、提高林木质量和林地生产力[17-18],以及对充分发挥森林生态和经济效益具有重要作用[8,19]。
光皮桦(BetulaluminiferaH. Winkl.)是我国南方特有速生用材树种和造林先锋树种,有良好的维持地力、涵养水源功能,对喀斯特地貌的植被恢复有着重要的意义[20-21]。对光皮桦的研究主要涉及在生物学特性、育苗技术[22]、生物循环和积累[23-24]、遗传育种[25]和生态效益[20]等方面。关于光皮桦次生林抚育间伐的系统研究主要集中在林分生长和经营技术等方面[26],而抚育间伐对光皮桦林下植被多样性和可持续性方面研究较少。贵州作为光皮桦天然次生林的主要分布地区,对长江、珠江上游生态屏障起着至关重要的作用。本文以贵州黔东南光皮桦天然次生林为研究对象,对抚育间伐3 a后的林下植物多样性和天然更新进行调查,系统分析不同抚育间伐强度对光皮桦林下植物多样性和更新发育的影响,以期为光皮桦次生林合理经营提供参考。
1 研究区概况
试验林分位于贵州省黔东南苗族侗族自治州麻江县西部的谷硐镇翁牛村,海拔1 100~1 370 m,平均海拔920 m。气候类型为亚热带季风温湿气候,年降水量1 200~1 500 mm,且主要集中在5—7月份。年日照时间1 200 h,年均气温15 ℃,相对湿度约80%。试验地位于东南坡中坡位,地处阳坡,平均坡度25°。地带性土壤为暗棕壤土。研究区森林类型属天然次生林,处于中幼龄林发展阶段,以光皮桦为主,乔木树种主要有云贵鹅耳枥(Carpinuspubescens)、白栎(Quercusvariabilis)、杉木(Cunninghamialanceolata)、三角槭(Acerbuergerianum)等。灌木主要有油茶(Camelliaoleifera)、香叶树(Linderacommunis)等。草本主要有芒萁(Dicranopterisdichotoma)、野燕麦(Avenafatua)等。
2 研究方法
2.1 样地设置
在尽可能使地形因子和土壤条件保持一致的前提下,选取生长情况基本一致的光皮桦次生林作为研究对象,立地类型为东南坡向,斜坡阳坡,土壤为暗棕壤土,试验林分海拔为1 100~1 370 m,林分密度为2 800~3 025株/hm2。采用4种不同强度处理,即设立弱度间伐(10%)、中度间伐(20%)、强度间伐(30%)和对照(未间伐)标准地,样地大小均为20 m×20 m,每种间伐强度重复3次,共12块标准地。根据间密留匀、留优去劣的总体原则,伐去生长不良以及非目的树种,保留乔木幼苗幼树,使保留木相对均匀分布,并在样地四周布设围栏,避免人为破坏。2014年9月份对样地进行间伐,2017年10月份进行调查。试验林分概况见表1。

表1 试验林分基本概况
2.2 样地调查
灌木层和幼苗调查:在样地中心和四角各设4 m×4 m的灌木样方,调查灌木种类、个体数和高度、盖度等。将更新的光皮桦幼苗(树高≤1.3 m)统计在灌木植物中。同时,单独记录光皮桦幼苗的数量和高度,将幼苗根据其高度(h)分为5个等级[31]:第1等级(h<30 cm)、第2等级(30 cm≤h<50 cm)、第3等级(50 cm≤h<100 cm)、第4等级(100 cm≤h<150 cm)、第5等级(h>150 cm)。
草本层调查:在每个4 m×4 m的灌木样方的左下角设1个1 m×1 m的草本样方,调查记录草本植物的种类、株数(丛数)、平均高度和平均盖度等。
林窗调查:采用对角线法调查林窗数量、林窗的长轴与短轴的长度和林窗高度(即林窗边界木冠层平均高)。林窗的形状近似于椭圆形,因此多采用椭圆面积公式计算林窗的面积。林窗面积(A):A=πLW/4。式中:L为椭圆长轴长,W为短轴长。
2.3 生物多样性评价指标
丰富度指数(R):用相对物种丰富度表示:R=S。式中:S为物种总数量。
辛普森(Simpson)指数(D):D=1-∑(Ni/N)2。式中:Ni为第i个种的个体数,N为样方内所有物种总个体数。
香浓-威纳(Shannon-Weiner)指数(H′):H′=-∑(Pilog2Pi)。式中:Pi是第i个种在全体物种中的比例,以其个体数占总个体数的百分数表示。
2.4 数据处理
数据用Excel 2007整理后,利用SPSS 21.0进行统计和分析,用LSD法多重比较进行差异显著性检验。
3 结果与分析
3.1 不同间伐强度对光皮桦天然次生林林下灌草种类的影响
由表2可见,间伐3 a后调查样地内共发现维管植物46种,分属27科41属。灌木17科29属31种,优势种为白栎、鹅耳枥和香叶树等;草本10科12属15种,其中禾本科和马鞭草科植物均有4种,占草本物种数的25%,优势种为芒草、野燕麦等。对照样地调查到的灌木种类最少,共13种,随着间伐强度增加,灌木物种数逐渐增多,弱度、中度和和强度间伐样地的灌木层物种数分别比对照增加了23.07%、53.84%和69.23%。而草本层物种数表现出一定波动性,但各处理间差异不大。弱度、中度和和强度间伐样地草本层物种有9种,占草本总数60.0%,对照样地有8种。随着间伐强度的增大,林下植被由耐阴的(或中性)植物(如荆条、大青)向阳性(或喜光)植物(如构树、刺槐)变化趋势。未间伐样地草本层植物主要由凤尾蕨、芒萁等耐阴植物组成,间伐样地出现了对光照要求较高的野燕麦、野地瓜等阳生性植物,且长势较好。间伐后灌木层的植物组成增幅大于草本层,这是由于间伐改善林下灌木层的光照和温湿度条件,加速了植物更新演替,喜光植物出现促进灌木层种类增加;而灌木通过对草本层的光线进行一定截留,对草本层耐阴草类更新起抑制作用,使草本层物种数上下波动。

表2 不同间伐强度光皮桦林下的植物种类

续(表2)
3.2 间伐强度对林下灌、草层多样性指数的影响
由表3可知,间伐3 a后,光皮桦林下灌木层的物种丰富度随间伐强度的增加而增加,光皮桦次生林不同间伐强度林下灌木层的物种丰富度均高于对照,强度间伐样地灌木层的物种丰富度最高。强度间伐样地和中度间伐样地分别与对照样地的物种丰富度差异显著(P<0.05);林下灌木层的香浓-威纳指数(H′)也表现出间伐组显著高于对照,但不同间伐强度之间差异不显著(P>0.05);中度间伐样地和弱度间伐样地的皮卢均匀度指数都高于对照,强度间伐样地的皮卢均匀度指数低于对照,不同间伐强度的皮卢均匀度指数与对照之间差异不显著(P>0.05),但不同间伐强度之间的皮卢均匀度指数差异显著(P<0.05);不同间伐强度之间的辛普森指数差异均不显著(P>0.05)。
间伐3 a后,不同间伐强度,光皮桦林下的草本层物种丰富度指数均高于对照,且达到显著水平(P<0.05),但不同间伐强度之间差异不显著;草本层的香浓-威纳指数随间伐强度的增加而增大,不同间伐强度的香浓-威纳指数分别与CK差异显著(P>0.05),不同间伐强度之间的香浓-威纳指数差异显著(P>0.05);随着间伐强度的增强,草本层的辛普森指数和皮卢均匀度指数虽有所增加,但各样地之间差异均不显著(P>0.05),说明间伐并不能显著影响草本层的辛普森指数和皮卢均匀度指数,由于间伐强度大,林分光照充分,林下灌木养分充足,优势物种竞争能力强,生长发育更好,物种富集度高,导致分布均匀度有所下降。

表3 不同间伐强度林下植物多样性指数
3.3 间伐强度对林下更新幼苗的影响
由表4可知,对照样地的幼苗主要分布于0~150 cm,总密度为4 100株·hm-2。间伐3 a后,弱度、中度和和强度间伐样地的林下各层总密度分别比对照增加了350、400、850株·hm-2,各间伐强度林下更新幼苗密度基本上随高度级的增加呈现先增后减的变化,均呈现以50 cm≤h<100 cm高度级为主的分布。在<30 cm亚层中,对照样地的幼苗密度最大,达到了1 050株·hm-2;0≤h<50 cm亚层中,幼苗密度在各样地的分布较均匀。各间伐强度在50 cm≤h<100 cm高度级的林下幼苗密度从小到大依次为中度间伐样地、弱度间伐样地、强度间伐样地、对照样地。强度间伐样地中,处于100 cm≤h<150 cm和h≥150 cm高度级的幼苗株数最大,分别达到1 125、625株·hm-2,林下植被高度层次增加。

表4 不同间伐强度光皮桦林下幼苗的数量和苗高变化
3.4 间伐对林窗的影响及其相关性
由表5可知,随着间伐强度的增加林窗数量逐渐增加,各处理林窗总面积变化趋势与林窗数量基本一致,强度间伐样地林窗总面积比对照样地高出75.38%,抚育间伐对光皮桦林分的林窗数量和面积的增加有主导作用。
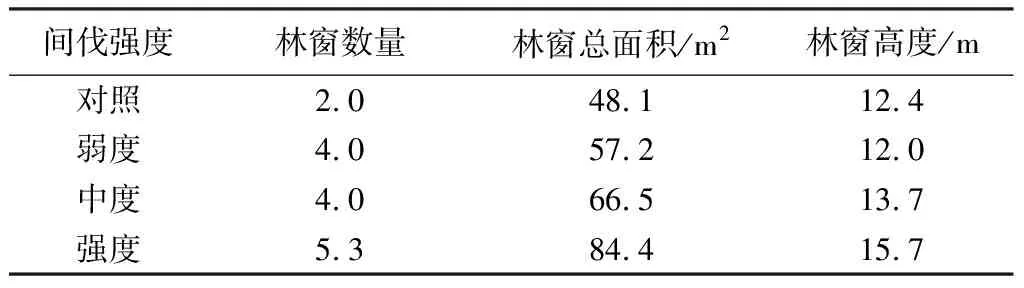
表5 不同间伐强度光皮桦林分的林窗数量和面积

表6 灌草层物种多样性指数与林分环境因子的相关性
4 结论与讨论
抚育间伐改变了林内植物组成区系,植物种、属和科数随间伐强度的增大而增加,植物多样性指数、林下幼苗数量及林窗面积均随间伐强度的增大而表现不同程度的增加,间伐强度在30%下拥有更丰富的植物多样性和较强的天然更新。间伐不仅能够促进光皮桦林下植物多样性,而且有助于幼苗生长和更新,强度间伐后短期内林下植物多样性较高、幼苗更新情况良好。
光皮桦天然次生林下灌草层物种数量随着间伐强度的增大而增加。由于光皮桦是强阳性树种,且主干通直,林冠层枝叶较多,对阳光截留较多,导致高密度或高郁闭度林分的林下环境阴冷潮湿而影响灌草层的生长。抚育间伐降低乔木层的郁闭度,使林隙增多,太阳直射透光率增强,保留木林下光热条件得以改善,林地空间异质性增加,提高了林下植物种类出现的不确定性[7],加速新物种入侵与定殖,耐荫树种与中性树种逐渐向阳生树种演替,促进林下植物种类增加[6]。本研究中,灌木层的物种数增幅大于草本层,与马双娇等[8]、王媚臻等[27]研究的结果一致。由于灌木层距离乔木层最近,能最先接收乔木层密度变化所产生的冠层结构变异引起植被分化,草本层对抚育间伐的反应比灌木层慢。
抚育间伐提高了光皮桦天然次生林林下植被多样性,抚育间伐的对灌木层和草本层的影响不同,草本层植物多样性指数随间伐强度增大而升高,而灌木层物种呈先增后减趋势。灌木层和草本层物种多样性大致在中、强度间伐呈现较高峰值,说明这两种间伐强度林内光照资源较好,阳生植物逐渐生长并开始显现优势,与Alem et al.[28]研究的结果一致。由于强度间伐形成的生境类似于演替早期阳性环境[29],间伐后到达地面的光线增加,使气温升高、空气湿度降低以及植物光合作用增强,营养物质的积累使土壤肥力增强,微生物物种数量增加,林分内光温水汽肥条件的改善,使可容纳物种的阈值有所提高,为新物种的定居提供了机会[7,20,30],恰好解释了林下物种丰富度指数、Shannon-Weiner指数随间伐强度增大而显著提高的结果。但与李淑辉等[31]研究的结果不同,由于间伐对灌木层干扰较大,原有生态系统的稳定性受到改变甚至破坏,短期内难以恢复,同时也由于研究区域、演替发展阶段与抚育间伐强度的不同所导致。间伐后,林下植被的Pielou均匀度指数无显著变化,且草本层均匀度指数在强度间伐时有所下降,这与众多研究学者结果一致[27,29]。其原因是间伐后乔木层对林下灌草层的抑制减弱,林窗分布不均致使林内环境变化不均匀,植物生存条件发生改变,导致林内植物物种分布存在一定差异。但也有研究表明,抚育间伐后,林下植物的Pielou均匀度指数有所下降或显著增加[8,32],原因是研究地理位置、植被类型、土壤肥力、气候条件等存在一定差异。
不同间伐强度,林下更新幼苗总密度、林窗数量和林窗面积均大于对照组,且随着间伐强度增大而逐渐升高,与已有研究结果一致[17,33-34]。由于对乔木层的间伐增加了林窗数量,改善了林下郁闭的小气候生境,提高林下植物获取资源的有效性[35],生态空间的释放有利于灌木草本植物的传播繁殖,同时灌草层竞争压力减弱,生态位互补效应增强[20],为更新苗创造了适宜的生长环境,加速了林下植被的更新演替。但与米爽等[11]研究结果不一致,这是由于间伐方式以及针阔混交林形成的生境不同,林下更新幼苗物种对外界干扰的敏感程度不同。间伐样地更新幼苗在较大高度级的幼苗密度所占比重随着间伐强度增大而增大,表明抚育间伐提高了光皮桦天然次生林林下幼苗的更新能力,与秦燕燕等[36]、罗桂生等[37]研究结果一致。
但在实际经营实践中,应对间伐后的林分补植优良针叶或其他树种,丰富林分植物的多样性,同时形成针阔混交的较稳定结构。由于本研究伐后间隔时间较短,只能反映了间伐对林分林下植物多样性和天然更新的短期影响,不能完全反映其生长状况。且林下植物多样性和更新状况随着林分逐渐郁闭以及采伐方式、频度不同而综合受影响,应对不同采伐处理的林分尽可能详细地对其他环境因子进行长期监测调查,以期得到更准确的间伐对林下植被影响的机理。抚育间伐对林下植被多样性的影响是一个长期的动态过程,受到时间和空间尺度的影响,系统研究还需设置固定样地进行长期观测,全面了解林分变化规律。
猜你喜欢
中华建设(2024年3期)2024-03-16 08:04:52
中华建设(2023年10期)2023-10-07 05:23:04
西南林业大学学报(2022年5期)2022-10-17 09:56:38
少儿美术(2020年8期)2020-11-06 05:21:38
农业机械学报(2020年1期)2020-03-13 00:27:38
中国医药指南(2018年16期)2018-07-13 02:28:32
西部林业科学(2018年4期)2018-02-14 22:43:50
西南林业大学学报(2014年6期)2014-07-24 19:02:11
小说月刊(2014年1期)2014-04-23 08:59:59
中南林业科技大学学报(2013年12期)2013-12-29 03:00:39
