
毛竹向阔叶林扩展过程细根可塑性变化
2021-11-02 02:38刘广路范少辉刘希珍农珺清申景昕
中南林业科技大学学报 2021年10期
黄 彪,刘广路,范少辉,刘希珍,冯 云,农珺清,申景昕
(国际竹藤中心 竹藤科学与技术重点实验室,北京 100102)
毛竹Phyllostachysedulis林资源是我国亚热带地区一种重要的森林资源,是仅次于马尾松和杉木的第三个人工用材树种。毛竹具有地下茎,是一种大型单轴散生竹种,生长特征独特。毛竹笋对光具有极度忍耐性使其能够在相邻的郁闭林地中生长[1],其生长所需的碳水化合物可以由鞭根系统母竹和根系的传输供给[2]。在竹阔混交林中,毛竹具有比阔叶林更大的比根长增幅和生长速率和周转率[3],具有很强的向周边系统扩展能力[4-5]。森林资源清查数据表明,近20年来毛竹林年平均面积增长率超过3%,与周边群落之间存在着广泛的交错带。竹产业迅速发展带来的竹资源需求增加,进一步促进了毛竹向周边群落的扩展进程。毛竹的扩展在增加竹资源供给的同时,也表现出降低群落内物种生物多样性[4,6-7]、阻碍阔叶树种更新[8]等负面影响。开展毛竹扩展策略的研究,可以为合理调控毛竹林扩展,保护生态安全提供科学依据。
植物功能性状是植物在长期适应环境过程中,通过内部不同功能之间协同进化,形成的能够响应外部环境变化的形态和功能[9-10],具有简单易测的特点,在植物与环境关系的研究中被广泛应用。细根作为植物吸收水分和养分的器官,其性状特征对植物的生长和分布具有重要的指示作用[10]。研究表明,当植物受养分限制或种间竞争时,通过增加比根长、比表面积或分支模式等提高对养分的获取能力或竞争力[11-13],同时根系组织中一些元素含量也与植物根系的生理生态功能密切相关[14]。毛竹向阔叶林扩展时,林分和土壤环境条件等均发生变化,开展毛竹扩展过程中细根功能性状的变化规律研究能够更好的揭示毛竹扩展策略。研究表明,毛竹向阔叶林扩展过程时,个体胸径和叶面积指数增加[15],叶功能性状发生适应性改变[16],但有关毛竹扩展过程中细根性状可塑性的研究较少,主要集中在毛竹纯林与竹阔混交林的对比研究上[3]。此外,年龄对植物细根的生长有重要的影响,有的树种细根生物量随着年龄的增长呈增加的趋势,如樟子松Pinus sylvestrisL[17]、欧洲赤松Pinus sylvestrics[18]、杉木Cunninghamia lanceolata[19];有的树种细根生物量随着年龄的增长呈降低的趋势,如欧洲山毛榉Fagus sylvatica[18],但有关不同年龄毛竹在扩展过程中细根性状的研究未见报道,不同年龄细根性状可能随毛竹扩展表现出不同的变化规律,使毛竹在生长竞争中处于有利地位。根序能较好的预测细根结构与功能的关系[20],大多数研究都把直径<2 mm 根作为同一个单元进行研究,而相关的研究表明这部分的根在形态和功能上都有大量的变化[21-22]。当前毛竹细根的研究主要针对小于2 mm 细根的研究,对小于1 mm 细根的研究报告较少,但有研究表明直径小于1 mm 的根系在根系中占据主要地位[23],研究毛竹扩展过程中0~1 mm 细根性状特征可以更深入的揭示毛竹扩展的细根策略。本研究以毛竹-阔叶林扩展界面为研究对象,研究了不同年龄毛竹在扩展过程中细根生物量、比根长、根长密度、细根主要养分含量及比值的变化规律,从而验证以下假设:1)随着毛竹扩展,毛竹细根性状发生可塑性变化;2)随着毛竹扩展,不同年龄毛竹的可塑性反应是否一致;3)0~1 mm 细根与1~2 mm 细根形态可塑性反应是否存在差异。
1 研究区概况与研究方法
1.1 研究区概况
实验地位于福建永安竹林生态定位观测研究站天宝岩国家级自然保护区观测区,117°28′03″~117°35′28″E,25°50′51″~26°01′20″N。属戴云山余脉,中低山地貌,海拔高580~1 604.8 m,气候属中亚热带东南季风气候型。观测区年平均气温15℃,绝对最低温度-11℃,绝对最高温度40℃;年平均相对湿度80%以上,无霜期290 d左右。海拔800 m 以下区域主要为竹林,覆盖率达96.8%,主要竹种为毛竹,土壤为红壤或红黄壤。其中大部分毛竹林是经过约30 a 自然扩展形成,其间混杂少量马尾松Pinus massoniana、杉木、闽楠Phoebe bournei等树种,存在着大面积的竹林-阔叶林交错带,形成的竹阔混交林主要是毛竹林经过近10 a 向阔叶林扩展形成,阔叶树种主要是以闽楠为优势种的次生林,林龄40~50 a,主要伴生种有南酸枣Choerospodias axillaris、木荷Schima superba等。
1.2 研究方法
在研究区内选择典型的毛竹向阔叶林扩展的过渡地带,沿毛竹向阔叶林扩展方向设置3 个生长条件较好,大小为10 m×50 m 的调查样带,并对每一条样带平均划分为5 个样方,每个小样方的尺寸大小为10 m×10 m(表1)。从毛竹林到阔叶次生林方向分别编号,依次为1、2、3、4、5 号样方,并调查每个小样方内植株数量、胸径、树高,记录年龄。根据不同年龄毛竹的平均胸径,确定标准竹。根据每样方内测定毛竹的平均胸径,在每个样方内选择1、2、4、6年生的标准竹。因为并非每个样方内4 种年龄的毛竹均有生长,共选取到生长良好的1年生标准竹20 株,2年生标准竹18株,4年生标准竹19株,6年生标准竹18株。计算每个样方内毛竹胸高断面积与林分总胸高断面积的比值(毛竹比例),靠近阔叶林为扩展前沿,毛竹比例低,随着毛竹的扩展,毛竹比例逐渐增加,扩展后期为毛竹纯林。

表1 样带基本情况†Table 1 The basic condition of research belt transect
1.2.1 细根样品的选取
采用标准法采集细根样品,根据以上选取标准竹为中心,以20、40、60 cm 为半径划弧,在每条弧线上分别选取样品采集点。沿着毛竹向阔叶林扩展方向上依次在样方内用内径为65 mm 的根钻采集原状土,深度为0~30 cm,每株标准竹获取3 份样品,共获得根系样品225 份。把获取的土壤原状土用孔径为2.0 mm 的土壤筛初步分离土壤、碎石等杂质,然后将分离出的根系置于0.85 mm筛网在流水中冲洗,将细根上残留的杂物冲洗干净。将冲洗干净的根系平铺在蒸发皿内,利用游标卡尺、镊子、放大镜等工具仔细挑选出毛竹细根。根据根系构型特征、颜色、弹性、气味等区分毛竹活根和死根,小心分离出毛竹细根系。本研究将直径小于2 mm 的根定义为细根,并分为两个径级:I级(0~1 mm)和II 级(1~2 mm)。
1.2.2 细根参数计算
将分好径级的细根分成两部分,其中将各级别1/3 左右生物量的新鲜根系利用WinRHIZO TRON软件进行扫描并计算根长后,将细根放在85℃的烘箱中烘干并称质量,用于计算比根长。剩余2/3左右生物量样品烘干称质量。将两部分烘干根系样品相加,利用公式计算得到每一层土芯样品的生物量。计算完以上指标之后,将烘干的细根进行粉碎,装入自封袋中,用作细根养分的测定。根据不同级别细根根长、生物量和取样体积,计算细根生物量(记为WFRB,g·m-3)、比根长(记为LSRL,m·g-3)和根长密度(记为DRLD,m·m-3)[14-15]。

式(1)~(3)中:m为细根质量(g),指单位面积上所有活细根的干质量,l为细根长(m),π为圆周率,d为土钻内径(cm),h为土钻高度(cm)。
1.2.3 细根养分含量的测定
由于采用根钻采集的细根数量较少,因此将同一株标准竹采集的细根混合成一个细根样品,并将混合后的细根样品利用粉碎机粉碎,供测定养分使用。细根碳含量(C)采用高温外热重铬酸钾氧化-容量法,细根N 含量采用凯式定氮法,细根磷含量为钒钼黄比色法。
1.3 数据处理
运用SPSS16.0 软件对细根生物量、细根比根长、根长密度、细根养分特征等进行单因素方差分析(One-way ANOVA)、多重比较(LSD)、相关性分析(Pearson)等分析处理。采用Excel 2010 软件进行数据整理、制图表。
2 结果分析
2.1 毛竹扩展进入阔叶林细根变化特征
2.1.1 毛竹所占比例对细根性状特征的影响
随着毛竹比例的增加,毛竹细根生物量和根长密度呈增加的趋势,比根长呈降低的趋势(图1)。细根生物量和根长密度随毛竹比例增加的上升幅度达到极显著水平(P<0.01,图1a、c),细根生物量在毛竹比例为100%阶段为1 220.54±65.08 g·m-3,毛竹比例为2%时生物量为292.59±73.86 g·m-3,为毛竹比例为100%时的24%;根长密度在毛竹比例为100% 阶段为6 205.01±250.80 m·m-3,毛竹比例为2% 时根长密度为3 249.37±1 174.14 m·m-3,为毛竹比例为100%时的52%。毛竹比例为2%时细根生物量和根长密度的标准误差都高于毛竹比例为100%时的标准误差,反映了细根生物量和根长密度在扩展前沿较小,变化幅度较大。比根长随毛竹比例增加而降低,降幅达到极显著水平(P<0.01,图1b)。比根长在毛竹比例为100%阶段为4.70±0.13 m·g-1,毛竹比例为2%时比根长为5.58±0.59 m·g-1,是毛竹比例为100%时的1.19 倍;同时毛竹比例为2%时比根长的标准误大于毛竹比例为100%时的标准误,反映了比根长在扩展前沿的变化幅度更大。
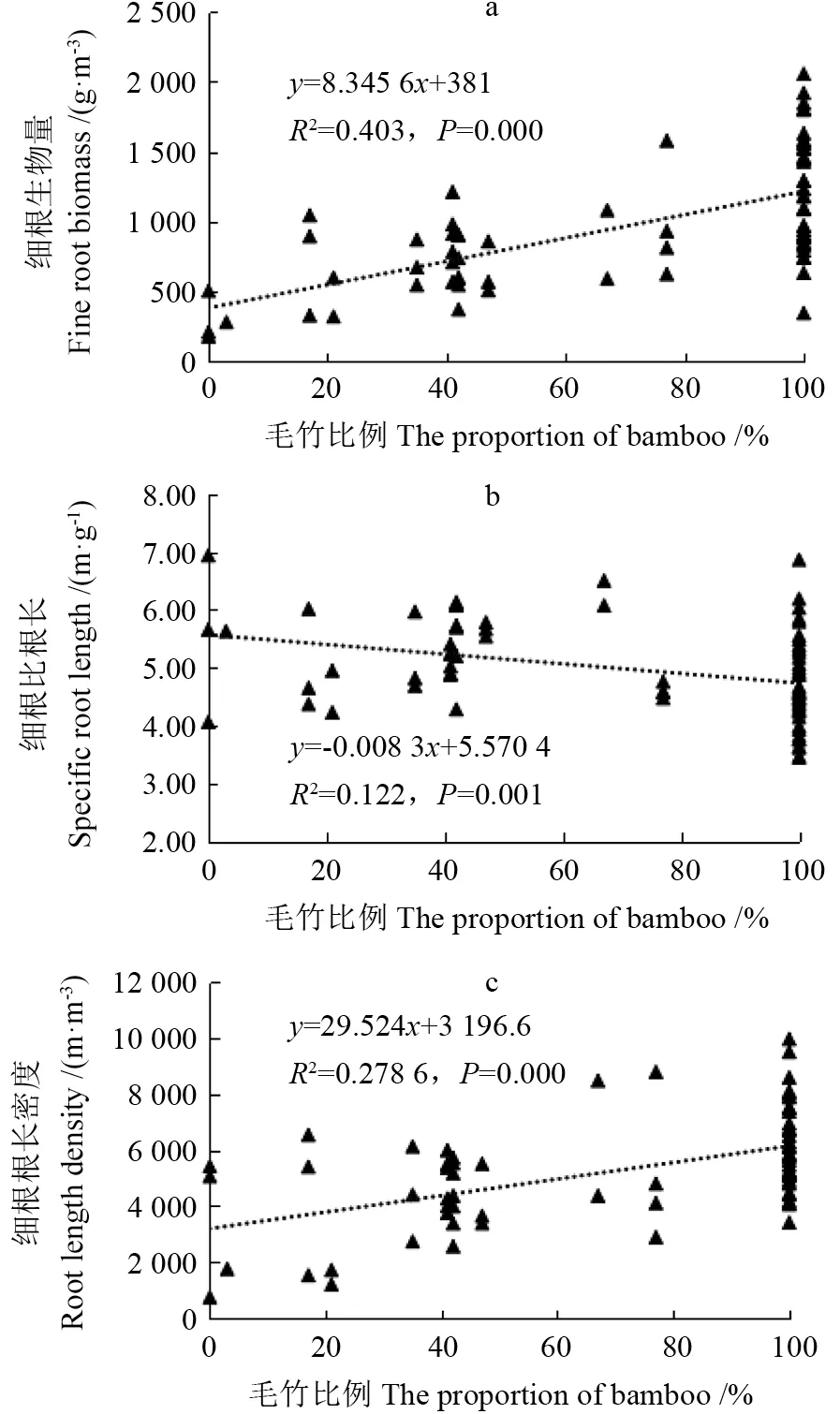
图1 毛竹向阔叶林扩展细根生物量(a)、比根长(b)和根长密度(c)变化Fig.1 The change in fine root biomass (a),specific root length (b),and root length density (c) of moso bamboo
2.1.2 毛竹年龄对细根性状特征的影响
不同年龄毛竹细根性状随着毛竹比例增加的变化趋势不同(图2)。细根生物量虽然随着毛竹比例的增加呈升高的趋势,但是不同年龄细根生物量随着毛竹比例的增加的变化趋势不同。1、2年生毛竹细根生物量随着毛竹比例的增加呈“升—降—升”的波浪型变化,4年生毛竹细根生物量随着毛竹比例的增加呈增加的趋势,6年生毛竹细根生物量随着毛竹比例的增加呈先上升后降的趋势。毛竹比例为20%时,1、2年生毛竹细根生物量较高,4、6年生毛竹细根生物量较低;在毛竹比例为80%时,1、2年生毛竹细根生物量较低,4、6年生毛竹细根生物量较高(图2a)。比根长的变化趋势与细根生物量的变化趋势相反,不同年龄毛竹比根长随着毛竹扩展的变化趋势也不同。1年生毛竹比根长随着毛竹比例的增加呈降低趋势,2年生毛竹呈“降—升—降”的波浪型变化,4年生毛竹呈先升高后降低的抛物线型变化,6年生毛竹呈缓慢上升的趋势(图2b)。不同年龄细根根长密度随毛竹扩展的变化趋势也不尽相同,1年生毛竹细根根长密度随着毛竹比例的增加,变化趋势不明显;2年生毛竹根长密度随着毛竹比例的增加呈“升—降—升”的波浪型变化;4、6年生毛竹根长密度随着毛竹比例的增加呈先上升后降低的趋势,在毛竹比例为80%时达到最大值(图2c)。

图2 不同年龄毛竹细根生物量(a)、比根长(b)和根长密度(c)变化Fig.2 The change in fine root biomass (a),specific root length (b),and root length density (c) of moso bamboo with different ages
2.1.3 0~1 mm 细根和1~2 mm 细根性状的差异分析
0~1 mm 细根生物量、比根长、根长密度显著高于1~2 mm细根生物量、比根长、根长密度(图3)。其中,0~1 mm 细根生物量是1~2 mm 细根生物量的1.82 倍,0~1 mm 细根生物量平均为669.97±88.07 g·m-3,1~2 mm 细根生物量平均为669.97±45.73 g·m-3(图3a);0~1 mm 细根比根长平均为8.82±0.93 m·g-1,1~2 mm 细根比根长平均为2.49±0.31 m·g-1,0~1 mm 细根比根长是1~2 mm 比根长的3.54 倍(图3b);0~1 mm细根根长密度平均为5 021.82±582.92 m·m-3,1~2 mm 细根根长密度平均为749.56±84.84 m·m-3,0~1 mm 细根根长密度是1~2 mm 比根长的6.70 倍(图3c)。不同扩展阶段0~1 mm 和1~2 mm 细根的变化幅度不同。0~1 mm 细根生物量、根长密度扩展前期比后期低32.14%和14.55%,比根长高23.03%;1~2 mm 细根生物量、根长密度扩展前期比后期低44.04%和29.15%,比根长高8.88%。
2.2 毛竹扩展进入阔叶林细根养分含量变化特征
随着毛竹比例的增加,细根碳、氮含量及C∶N的变化差异未达到显著水平,细根碳、氮含量的平均值分别为484.42、3.55 g·kg-1,C∶N 的平均值为140.08;磷含量和N∶P 的差异达到极显著水平(P=0.002,表2)。其中,磷含量随着毛竹比例的增加,呈先增加后降低的趋势,毛竹比例为40%~70%时,磷含量较高,平均为0.23 g·kg-1,比扩展前沿高64.29%,比扩展后期高43.75%(图4a);N∶P 呈先降低后升高的趋势,平均为16.66,比扩展前沿(25.84)低35.54%,比扩展后期(21.70)低23.21%(表2)。
2.3 细根形态特征与养分含量相关性分析
毛竹细根生物量、比根长和根长密度与细根碳、氮、磷含量及C∶N 和N∶P 比的相关系数不同(表3)。比根长与氮、磷含量正相关,与碳含量、C∶N和N∶P 负相关,相关性未达到显著水平(P>0.05);根长密度和细根生物量与氮、磷含量负相关,与碳含量、C∶N 和N∶P 正相关。其中,细根生物量与叶片磷含量和N∶P 的相关性达到显著或者极显著水平,与其它指标的相关性未达到显著水平。

表3 细根形态特征与养分含量相关性分析†Table 3 Correlation analysis between fine root morphology and nutrient content
3 结论与讨论
3.1 结 论
近年来,胡文杰等[26]主要研究比较了毛竹不同土壤层次间物理性质及养分含量的差异,潘璐等[27]讨论了毛竹林扩张对周边森林群落菌根系统的影响,毛翼虎等[28]分析了毛竹林细根生长的趋富特化及对养分限制的响应,研究表明毛竹扩展进入相邻林分,通过影响周围森林群落结构、土壤质量、幼苗更新及幼树生长,从而影响整个森林生态系统[29]。基于毛竹扩展过程中的细根可塑性变化的研究发现,随着毛竹扩展进入阔叶林,毛竹的生长环境发生变化,细根生物量、根长密度上升,比根长下降,N 含量先上升后下降,N∶P先下降后上升,毛竹细根性状随着环境的变化发生了适应性的变化,表现出明显的可塑性;不同年龄毛竹细根性状随着毛竹扩展的变化存在差异,表现为不同年龄细根生物量、根长密度和比根长随着毛竹的扩展变化趋势不同,具有较强的互补效应;不同扩展阶段0~1 mm 细根与1~2 mm细根特征存在差异。
3.2 讨 论
3.2.1 毛竹扩展过程中的细根可塑性变化
克隆植物适应异质性环境时生物量分配发生改变是植物协调生长的重要生物学特性[30-31],根系生物量的大小在一定程度上反映细根的广布性[3],本研究中扩展前沿较小的细根生物量反映了在扩展前沿细根广布性较差,随着毛竹比例不断增加,毛竹细根广布性增加。当植物受养分限制或种间竞争时,通过增加比根长、比表面积或分支模式等提高对养分的获取能力或竞争力[11-13],植物的根长密度越大,养分及水分吸收的表面积就越大;比根长高则根系增殖速度快,活力也高,细根对土壤资源的利用效率也高[32-33]。本研究中扩展前沿毛竹细根根长密度较小,比根长较大,反映了毛竹在阔叶树种激烈竞争的环境下,通过增加细根比根长,提高吸收效率来更好的利用土壤资源。植物根系的生理生态功能除与其形态有关外,还与根系组织中一些元素含量密切相关[14]。“生长速率假说”认为快速增长的组织磷含量更高,N∶P 更低[34-36]。本研究中,毛竹不同扩展阶段细根P 含量和N∶P 与细根生物量之间存在显著的相关性,细根P 含量越高,N∶P 越小,生物量越低,比根长越大,反映了在扩展前沿毛竹通过增加养分利用效率来弥补细根生物量的不足。研究表明,N∶P<14 可以认为是受N 限制,N∶P>16 是受P 限制[37],本研究中扩展前期和后期的N∶P 均大于16,可能受到P 限制,在扩展中间阶段平均值为16.66 处于临界值,受P 限制较弱。
3.2.2 不同年龄毛竹细根性状随着毛竹扩展的变化差异
毛竹林是由地上不同年龄的毛竹单株和地下相互交错的鞭根系统连接在一起的一个整体,鞭根系统担负着养分吸收传输和新竹生长,是一种天然异龄林。本研究中,不同年龄细根生物量、根长密度和比根长随着毛竹的扩展变化趋势不同,具有较强的互补效应。在竞争性的环境中,树木通常通过增加比根长来增加细根的竞争能力[38],欧洲水青冈[39]和赤松细根比根长在竞争性的环境中显著增加[40]。随着毛竹扩展不同年龄毛竹细根发生显著变化,这种变化促进各年龄毛竹发挥自身的作用,在竞争生长中处于有利地位。最先扩展进入阔叶林的毛竹为1年生单株,其具有较其它年龄毛竹更大的比根长,使其在能够在竞争性的环境中更高效的利用土壤资源。研究发现,植物多样性增加会导致细根生物量、根长密度增加,细根具有“超产”现象[25,31],本研究中在生物多样性较高的扩展前沿毛竹的细根生物量较低,而扩展后期毛竹细根生物量较高,可能与毛竹细根生物量远高于常绿阔叶林细根生物量有关[3]。
3.2.3 不同扩展阶段0~1 mm 细根与1~2 mm细根特征差异
根系形态和分布状况决定了植被从土壤吸收水分和养分的能力,也反映了林木对立地条件的利用状况[41],一般随着细根直径的减小,根系N含量增高,比根长增加,土壤资源的获得效率更高,但同时细根周转也加快,导致更高的碳消耗[42]。低级根在根系数量上的绝对优势,保证了它们能够充分的吸收水分和养分并且传输到其它部分[43]。本研究中,0~1 mm 细根生物量、根长密度和比根长分别是1~2 mm 细根的1.82、6.70 和3.54 倍,毛竹分配更多的资源到0~1 mm 细根,反映了0~1 mm 细根比1~2 mm 细根具有更强的资源利用能力,在毛竹生长中具有重要的作用。研究也表明,扁刺栲<1 mm 的细根代谢活跃且具有吸收功能,N 含量最高,C∶N 最小,根生物量、根长、表面积和比根长均较高[23]。与1~2 mm 细根相比,0~1 mm 细根生物量、根长密度扩展前期比后期相比变化幅度更低,比根长的变化幅度更高。在有限的养分条件下,树木或者增加细根的生物量(或根长),或者改变根形态来增强其养分吸收能力[44],直径越小的根系其生理代谢活动较为活跃,根尖细胞分裂旺盛[45],0~1 mm 细根的吸收能力高于1~2 mm 细根,在毛竹根系中具有重要的作用。
本试验初始想法是为了揭示随着毛竹扩展毛竹细根性状的变化规律,主要研究了在毛竹扩展过程中不同年龄毛竹0~1 mm 和1~2 mm 细根生物量、根长密度、比根长、细根碳氮磷含量及比值的变化规律,探讨了毛竹细根性状随着毛竹扩展的适应性改变,比较了不同年龄、不同级别细根随着毛竹扩展的变化差异。在下一步工作中我们将进一步开展多因素实验研究,在实验设计中将毛竹比例、年龄双因素及交互作用的影响纳入实验研究,以期获得更多的规律信息,进一步阐明毛竹扩展进入阔叶林的细根策略,为实现毛竹林扩展调控提供更多支撑。
猜你喜欢
草业学报(2022年9期)2022-09-16
热带亚热带植物学报(2022年4期)2022-08-18
东北林业大学学报(2022年6期)2022-07-25
小小说月刊·下半月(2021年8期)2021-09-05
绿色科技(2021年5期)2021-04-09
意林·少年版(2020年13期)2020-08-02
东坡赤壁诗词(2020年2期)2020-06-04
阅读与作文(小学高年级版)(2019年3期)2019-04-20
动漫界·幼教365(大班)(2018年3期)2018-05-14
湖南大学学报·自然科学版(2016年12期)2017-05-12
