
Co-nanoencapsulated meloxicam and curcumin improves cognitive impairment induced by amyloid-beta through modulation of cyclooxygenase-2 in mice
2021-11-02 02:59:14MariaEduardaZianiGutierrezAnneSulyPintoSavallEdinadaLuzAbreuKellyAyumiNakamaRenataBemDosSantosMarinaCostaMonteiroGuedesDaianaSilvavilaCristianeLucheseSandraElisaHaasCarolineBrandQuinesSimonePinton
中国神经再生研究(英文版) 2021年4期
Maria Eduarda Ziani Gutierrez, Anne Suély Pinto Savall, Edina da Luz Abreu,Kelly Ayumi Nakama, Renata Bem Dos Santos, Marina Costa Monteiro Guedes,Daiana Silva Ávila, Cristiane Luchese, Sandra Elisa Haas, ,Caroline Brandão Quines, , Simone Pinton,
Abstract Alzheimer’s disease is a progressive brain disorder and complex mechanisms are involved in the physiopathology of Alzheimer’s disease.However, there is data suggesting that inflammation plays a role in its development and progression. Indeed, some non-steroidal antiinflammatory drugs, such as meloxicam, which act by inhibiting cyclooxygenase-2 have been used as neuroprotective agents in different neurodegenerative disease models. The purpose of this study was to investigate the effects of co-nanoencapsulated curcumin and meloxicam in lipid core nanocapsules (LCN) on cognitive impairment induced by amyloid-beta peptide injection in mice. LCN were prepared by the nanoprecipitation method. Male Swiss mice received a single intracerebroventricular injection of amyloid-beta peptide aggregates (fragment 25–35, 3 nmol/3 µL) or vehicle and were subsequently treated with curcumin-loaded LCN (10 mg/kg) or meloxicam-loaded LCN (5 mg/kg)or meloxicam + curcumin-co-loaded LCN (5 and 10 mg/kg, respectively). Treatments were given on alternate days for 12 days (i.e., six doses,once every 48 hours, by intragastric gavage). Our data showed that amyloid-beta peptide infusion caused long-term memory deficits in the inhibitory avoidance and object recognition tests in mice. In the inhibitory avoidance test, both meloxicam and curcumin formulations (oil or co-loaded LCN) improved amyloid-beta-induced memory impairment in mice. However, only meloxicam and curcumin-co-loaded LCN attenuated non-aversive memory impairment in the object recognition test. Moreover, the beneficial effects of meloxicam and curcuminco-loaded LCN could be explained by the anti-inflammatory properties of these drugs through cortical cyclooxygenase-2 downregulation.Our study suggests that the neuroprotective potential of meloxicam and curcumin co-nanoencapsulation is associated with cortical cyclooxygenase-2 modulation. This study was approved by the Committee on Care and Use of Experimental Animal Resources, the Federal University of Pampa, Brazil (approval No. 02-2015) on April 16, 2015.
Key Words: Alzheimer’s disease; curcumin; cyclooxygenase-2; lipid core nanocapsules; meloxicam; memory; rats; inflammation
Introduction
Alzheimer’s disease (AD) is a progressive brain disorder and has become the third major cause of disability and death for the elderly (Du et al., 2018). Genetic factors are considered to be preponderant for its pathogenesis with mutations occurring in the amyloid precursor protein and presenilin genes, both of which are associated with the metabolism of amyloid-beta peptide (Levy-Lahad et al., 1995).
The pathological process of this disease consists of the slow deterioration of cognition and memory loss, accompanied by mood alterations (Frere and Slutsky, 2018). Complex mechanisms are involved in the physiopathology of AD,however, there is data suggesting that inflammation plays a role in its development and progression. This is because indirect damage to neurons produces inflammatory mediators,which in turn could contribute to the pathological processes of AD (Tuppo and Arias, 2005; Cuello, 2017).
Some non-steroidal anti-inflammatory drugs (NSAIDs), such as meloxicam, which act by inhibiting cyclooxygenase-2 (COX-2) have been used as neuroprotective agents in different neurodegenerative disease models (Ah et al., 2010).Treatment with NSAIDs could block incipient inflammationdriven AD pathogenesis at early stages (Harrington et al.,2011). Notably, novel nanotechnological formulations with meloxicam have optimized its pharmacological effect in an AD model and minimized its toxicity in mice (Ianiski et al., 2016;Villalba et al., 2016).
In addition to inflammation, oxidative stress has been associated with aging and the neurodegenerative process(Du et al., 2018). Thus, acknowledging that antioxidants can reduce the deleterious effects of oxidizing species,such as reactive oxygen species, molecules with antioxidant and anti-inflammatory properties appear promising in the face of neurodegenerative process in AD (Du et al., 2018;Frere and Slutsky, 2018). In this sense, curcumin is a natural phenolic compound extracted from the root or rhizome of Curcuma longa (Sharma, 1976), whose anti-inflammatory and antioxidant properties have been reported (Morales et al., 2017; Sundaram et al., 2017). Indeed, curcumin is a wellknown reactive oxygen species scavenger (Guerzoni et al.,2017; Rakotoarisoa et al., 2019).
Chronic curcumin use reduces the loss of cognitive function and inflammation caused by AD (Aggarwal and Harikumar,2009; Sundaram et al., 2017). Similar to meloxicam,incorporation of curcumin into nanocapsules seems to be an alternative to improve its beneficial effects, since there is an improvement in its bioavailability, thereby increasing its anti-inflammatory effects (Hoppe et al., 2013; Jaques et al., 2013;Rakotoarisoa and Angelova, 2018).
Nanocapsule consists of a thin membrane surrounding a core (liquid, solid) with its size ranging from 10 nm to 1000 nm. Nanocapsules are submicroscopic colloidal drug carrier systems composed of an oily or an aqueous core surrounded by a thin polymer membrane. One specific type, lipid core nanocapsules (LCN), have an organogel in this core (Giacomelli et al., 2019; Nakama et al., 2020). The therapeutic efficacy of LCNs has been shown to improve the targeting and delivery of drugs to the brain, and LCNs have an important ability to cross the blood-brain barrier (Carreno et al., 2016;Sonvico et al., 2017). Based on these observations, the purpose of the present study was to analyze the effects of co-nanoencapsulated curcumin and meloxicam in LCNs on cognitive impairment and neuroinflammation in a mouse model of AD induced by amyloid-beta peptide.
Materials and Methods
Materials
Amyloid-beta peptides (fragment 25–35) were obtained from Sigma-Aldrich (St. Louis, MO, USA) and dissolved in sterile filtered water and aggregated by incubation at 37°C for 4 days before use. Poly(ε-caprolactone) (molecular weight 80.000 g/mol), curcumin, meloxicam and sorbitan monostearate (Span 60®) were obtained from Sigma-Aldrich.High performance liquid chromatography (HPLC) solvents were obtained from JT-Baker (Aparecida de Goiânia, GO,Brazil). All other chemicals were of analytical grade and obtained from standard commercial suppliers (Porto Alegre,RS, Brazil).
Lipid core nanocapsule preparation and characterization
LCNs were prepared by a nanoprecipitation method(Nakama et al., 2020). In brief, the LCN loading meloxicam(M-LCN, 0.3 mg/mL), curcumin (C-LCN, 0.6 mg/mL)and meloxicam + curcumin (0.6 + 0.3 mg/mL, MC-LCN)were prepared by interfacial deposition of poly(epsiloncaprolactone). The analytical method for the development,co-nanoencapsules and physicochemical characterization of nanocapsules of lipid nucleus of curcumin, meloxican and curcumin plus meloxican were described by our research group (Nakama et al., 2020). The organic phase(poly(ε-caprolactone), capric caprylic triglycerides, sorbitan monostearate drugs and acetone were solubilized at 40■C(Levy-Lahad et al., 1995). Then, this phase was poured into an aqueous solution with polysorbate 80. Curcumin-loaded LCN (0.6 mg/mL), meloxicam-loaded LCN (0.3 mg/mL)and meloxicam + curcumin-co-loaded LCN (0.3 + 0.6 mg/mL,respectively) were named LCN C, LCN M and LCN MC,respectively. Unloaded LCNs (LCN U) were prepared without any drug. All formulations were characterized according to particle size (laser diffraction, Mastersizer 2000 analyzer,Malvern Instruments, São Paulo, Brazil), zeta potential electrophoretic migration (Zetasizer® Nano ZS3600, Malvern Instruments, Westborough, MA, USA) and pH (Hanna Instruments, São Paulo, Brazil). Encapsulation efficiency and drug content were calculated following a previous study(Nakama et al., 2020). Briefly, curcumin and meloxicam were simultaneously determined by a high-performance liquid chromatography-diode array detector (Shimadzu LC System, Kyoto, Japan). For drug content, all formulations were dissolved in methanol and then quantified. For encapsulation efficiency, an aliquot of nanoformulation was placed inside an ultrafree-MC centrifugal filter (Merck-Millipore, São Paulo,Brazil) and centrifuged. The drugs in the ultrafiltrate were determined by a high-performance liquid chromatographydiode array detector.
Animals
Experiments were conducted using 67 male Swiss mice(40–50 g, 3-month-old). Animals were maintained at 22–25°C with free access to water and food under a 12-hour light/dark cycle with lights turned on at 7:00 a.m. Animals were obtained from the Federal University of Santa Maria, Brazil and used according to the guidelines of the Committee on Care and Use of Experimental Animal Resources, the Federal University of Pampa, Brazil (approval No. 02-2015) on April 16, 2015. All procedures involving animals concurred with the Animal Research Reporting ofIn VivoExperiments (ARRIVE)guidelines. All experiments, maintenance and manipulation of the animals were done according to the guidelines of CONCEA(National Council for Control of Animal Experimentation) law#11.794, of 08.10.2008 and normative resolution # 30, of 02.02.2016.
Experimental protocol
Animals were separated into eight groups (n= 8–9/group) and received:
(I) sterile filtered water (3 nmol/3 µL, intracerebroventricular(i.c.v.)) and unloaded LCN (LCN U) (17 mL/kg, gavage) (control group;n= 9);
(II) amyloid-beta peptide (3 nmol/3 µL, i.c.v.) and LCN U (17 mL/kg, gavage) (Aβ group;n= 9);
(III) amyloid-beta peptide (3 nmol/3 µL, i.c.v.) and curcuminloaded LCN (10 mg/kg, 17 mL/kg, gavage) (Aβ + LCN C group;n= 8);
(IV) amyloid-beta peptide (3 nmol/3 µL, i.c.v.) and meloxicamloaded LCN (5 mg/kg, 17 ml/kg, gavage) (Aβ + LCN M group,n= 8);
(V) amyloid-beta peptide (3 nmol/3 µL, i.c.v.) and meloxicam+ curcumin-co-loaded LCN (5 and 10 mg/kg, respectively, 17 mL/kg, gavage) (Aβ + LCN MC group,n= 9);
(VI) amyloid-beta peptide (3 nmol/3 µL, i.c.v.) and curcumin (10 mg/kg, in canola oil, 17 mL/kg, gavage) (Aβ + C group,n= 8);
(VII) amyloid-beta peptide (3 nmol/3 µL, i.c.v.) and meloxicam(5 mg/kg, in canola oil, 17 mL/kg, gavage) (Aβ + M group,n= 8);
(VIII) amyloid-beta peptide (3 nmol/3 µL, i.c.v.) and meloxicam+ curcumin (5 and 10 mg/kg, respectively, in canola oil,17 mL/kg, gavage) (Aβ + MC group,n= 8).
Animals from groups II to VIII were intracerebroventricularly injected with 3 nmol (3 µL) of aggregated amyloid-beta peptide, fragment 25–35. Group I received sterile filtered water (3 µL, i.c.v.) utilizing a microsyringe with a 28-gauge stainless-steel needle 3.0 mm long (Hamilton); local administration of amyloid-beta peptide or vehicle was performed by a “freehand” protocol with the bregma fissure as a reference point, as described by Haley and McCormick(1957). The treatments were initiated 24 hours after infusion of amyloid-beta peptide or vehicle, and mice were treated by intragastric administration (gavage) (i.g., 17 mL/kg body weight) on alternate days, once every 48 hours, as shown in Figure 1 and as previously described (Neervannan, 2006;Ianiski et al., 2016;). All the drug treatments (LCN C, LCN M or LCN MC) were carried out on days 2, 4, 6, 8, 10 and 12.Mice received a total of six doses of LCN C, LCN M or LCN MC. From the sixth day of experimental protocol, the mice performed behavioral tests. Twenty-four hours after the last drug administration, on the 13thday, animals were euthanized for dissection of prefrontal cortex and hippocampus used for quantitative analysis of COX-2 expression.
Open field test
Spontaneous locomotor activity was measured in the open field test (Walsh & Cummins, 1976). The floor of the open field apparatus, 50 cm × 50 cm × 50 cm, was divided into equal nine squares. Each animal was placed individually in the center of the area, and the number of segments crossed(four-paw criterion) and instances of rearing-up behavior were recorded in a 4-minute session.
Object recognition test
Long-term memory was measured with the one-trial object recognition task (ORT) method (Ennaceur, 2010), where the animals were exposed to an object in an open box made of wood (50 cm × 50 cm × 50 cm). Initially, mice went through habituation to the training box. Each mouse could freely explore the apparatus for 20 minutes per day during 2 consecutive days. After that, on the training day (on day 9 of experimental protocol), two equal objects (A1 and A2)were placed in the apparatus and animals were allowed to explore them freely for 5 minutes. Objects were made of plastic, had many colors and were attached to the floor so the animals could not displace them. There were no extra lights or sounds that could distract the animal. Then, 24 hours later, to measure long-term memory, one of the objects was randomly exchanged for a novel object (Ah et al., 2010) and mice were reintroduced into the apparatus for an additional 5 minute period. Exploration was defined as sniffing or touching the object with the nose and/or forepaws. The results were expressed as % of exploratory preference for each animal using the ratiotB/(tA+tB) × 100%, wheretAis the time spent for exploring the familiar object A andtBis the time spent for exploring the novel object B.
Inhibitory avoidance test
Aversive long-term memory was assessed with the inhibitory avoidance test (Pinton, Bruning, Sartori Oliveira, Prigol, &Nogueira, 2013). Briefly, the apparatus consisted of a single box with the presence of a safe platform and the mice were trained to associate the descent of this safe platform with an aversive stimulus (an electric shock of 0.5 mA was delivered for 2 seconds). Twenty-four hours after training, each mouse was placed again on the safe platform, and the time took to step down from the platform (transfer latency) was measured(cut-off 300 seconds). Latency (in seconds) was defined the time taken to fall from the platform in the acquisition and retention phase.
Tail immersion
A tail immersion test was carried out as previously described(Janssen, 1963) to determine the nociceptive reaction of mice and ensure the efficacy of the inhibitory avoidance test(Schimidt et al., 2014). The animals’ tails were placed in a water bath heated to 50°C, and the latency of their response was measured in seconds by watching the reflexive withdrawal of the tail after its immersion in water.
Western blot assay of COX-2
Western blot assay was carried out as previously described(Quines et al., 2018). Prefrontal cortices and hippocampi were manually homogenized in a glass–glass potter in ice-cold 0.32 M sucrose buffer (pH 7.4) containing 1 mM 4-(2 hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) and 0.1 mM ethylene glycol tetraacetic acid (EGTA) in the presence of protein inhibitor cocktail (Sigma-Aldrich Co., St. Louis, MI, USA). The samples (20 µg of protein and prestained molecular weight standards (Sigma-Aldrich Co.,)) were separated by 10%SDS-PAGE electrophoresis and transferred to nitrocellulose membrane (0.45 µm, Bio-rad) using Transfer-Blot® TurboTMTransfer System (1.0 A; 45 minutes) and equal protein loading was confirmed by Ponceau S staining. After blocking with 3%bovine serum albumin solution, the blots were incubated overnight at 4°C with rabbit anti-cyclooxygenase-2 (anti-COX-2)(1:1000; Cell Signaling, Danvers, MA, USA), and mouse anti-β-actin (1:5000; Cell Signaling) was marked as an additional control for the protein loading. After incubation with primary antibody, membranes were washed and incubated for 1 hour at room temperature with anti-mouse or anti-rabbit secondary antibodies conjugated with horseradish peroxidase (1:1000,Bio-Rad Laboratories, Hercules, CA, USA). The software ImageJ (Version 1.51p) (NIH, Bethesda, MD, USA) was used for quantification of optical density of the western blot bands.Each value was derived from the ratio between arbitrary units obtained by the COX-2 band and the respective β-actin band.The protein concentration was determined by the Coomassie blue method, using bovine serum albumin as a standard (1 mg/mL), as described previously by Bradford (1976).
Statistical analysis
We performed statistical analysis using one-way analysis of variance followed by the Tukeypost hoctest when necessary.For ORT, Student’st-test for one sample with theoretical mean= 50 was used. Experimental results are given as the mean± S.E.M. Probability values less than 0.05 (P< 0.05) were considered to be statistically significant. The statistical analysis was performed using the GraphPad Prisma (Version 6, São Paulo, Brazil).
Results
Characterization of lipid core nanocapsules
LCNs were successfully prepared using the nanoprecipitation method. The diameters ranged from 291 to 312 nm, with adequate polydispersity. Using the laser diffraction method,LCNs of 257 ± 5.6, 312 ± 5.5, 291 ± 1.53 and 293 ± 2 nm were observed for LCN C, LCN M, LCN MC and LCN U, respectively.The zeta potential was negative (22–36 mV), with a pH value between 6.01 and 6.98. The drug content ranged from 99.4 to 100.05 for all formulations (P> 0.05).
Meloxicam and curcumin-co-loaded LCN demonstrates greater effectiveness to recover amyloid-beta-induced memory impairment in mice
Initially, ORT was performed to verify the protective effect of meloxicam and/or curcumin formulations on memory in mice. All groups explored both objects equally (around 50%)during the training session (Figure 2A). In the long-term memory retention test, the control group showed a significant preference for exploring the novel object (P= 0.0262; Figure 2B). However, amyloid-beta-infused mice were not able to distinguish between the novel and familiar objects and spent a similar time in exploring these objects (P= 0.814; Figure 2B).Mice in the Aβ + LCN MC group explored the novel object for a longer duration than the familiar object (P= 0.021), similar to control mice (Figure 2B). These results provided evidence that amyloid-beta peptide injection induced long-term memory deficits in mice as measured with ORT, which were rescued by LCN MC treatment; thus, MC effectiveness was favored by conanoencapsulation.
During inhibitory avoidance training, there was no difference in the transfer latency time between groups (F(7,59)= 1.535;P> 0.05) (Figure 3A). In the probe test, the results revealed a significant difference in the transfer latency time between groups (F(7,59)= 4.955;P= 0.0002) (Figure 3B). Amyloidbeta peptide infusion caused an impairment in memory in mice, since amyloid-beta peptide significantly decreased the transfer latency time as compared to the control group (P=0.0014). Notably, meloxicam (P= 0.676), meloxicam LCN (P= 0.167), meloxicam + curcumin association (P= 0.971) and meloxicam + curcumin-co-loaded LCN (P= 0.969) protected partiality against this alteration, since these groups were similar to the control group. Importantly, only the meloxicam and curcumin association completely reversed amyloid-betainduced memory impairment, in both canola oil (P= 0.0384)and nanocapsule (P= 0.0282) formulations. Additionally, there was no significant difference in the latency time in the tail immersion test (data not shown) (F(7,59)= 1.514,P= 0.180).Thus, no animal took longer latency time than the control mice, allowing us to verify that none of the animals had altered nociceptive reaction systems and, therefore, we could perform the inhibitory avoidance test. Finally, drug treatment did not alter locomotor activity in mice (crossing and rearing numbers) (Table 1).
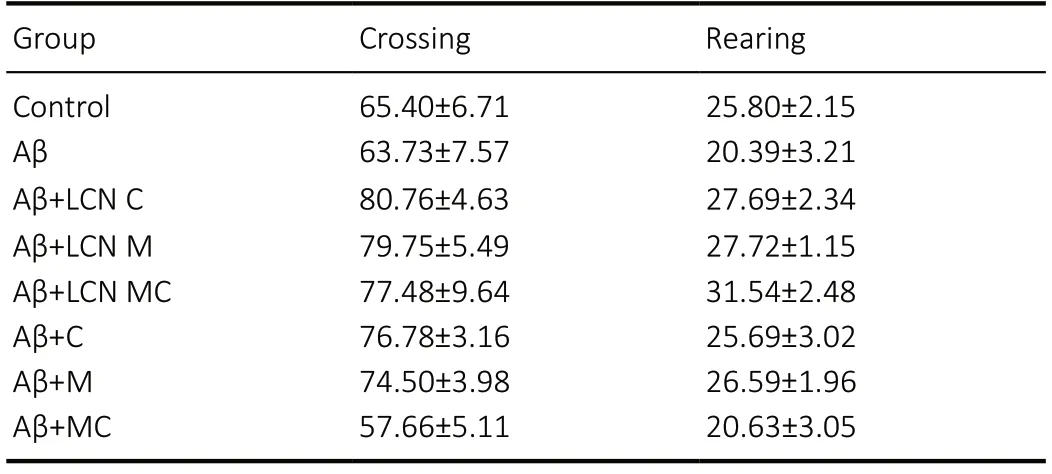
Table 1 |Effect of amyloid-beta peptide and different formulations of meloxicam and/or curcumin on locomotor activity in the Open Field Test in mice
Meloxicam and/or curcumin formulations attenuateamyloid-beta-induced memory impairment in mice via downregulation of COX-2 protein expression
The analysis of cortical COX-2 protein expression levels revealed a significant difference between the groups (F(7,32)= 5.546,P= 0.0003) (Figure 4A). Post hoc comparisons demonstrated that the amyloid-beta-treated group presented increased COX-2 protein expression levels in the prefrontal cortex when compared to the control group (P= 0.0039).Curcumin LCN treatment was not effective in restoring the protein expression levels of COX-2 (P= 0.0045, from control group). COX-2 protein expression levels in the Aβ + M, Aβ+ CM and Aβ + LCN M groups were similar to those in the control and Aβ groups. Importantly, oil formulations of curcumin as well as meloxicam and curcumin-co-loaded LCN were effective to reduce the amyloid-beta-increased COX-2 protein expression levels in the prefrontal cortex (P= 0.015 andP= 0.0487 when compared to the Aβ group, respectively).Furthermore, there was no significant difference in COX-2 protein expression level in the hippocampus of mice between groups (Figure 4B).
Discussion
The present study showed for the first time the beneficial effect of meloxicam + curcumin co-nanoencapsulation in a mouse model of AD. Collectively, among the formulations tested, meloxicam + curcumin co-nanoencapsulation showed the best effects on improving the memory deficits induced by amyloid-beta peptide in mice. LCN CM administration attenuated amyloid beta-induced memory impairment in mice as evaluated by inhibitory avoidance and object recognition tests. Furthermore, our findings also indicate that LCN CM exerted an anti-inflammatory action by downregulation of COX-2 expression in the prefrontal cortex of amyloid-betainfused mice.
AD is a multifactorial disease, associated with regional cerebral hypometabolism, extracellular amyloid-beta plaques,neuroinflammation and oxidative stress, beyond the loss of synaptic connections, neural death, and atrophy (Jeong, 2017;Velasques et al., 2018). AD is also associated with progressive dysfunction and a loss of neurons and synapses, particularly in the cortex and hippocampus. Deposition of amyloid-beta peptide in the brain is considered the primary event in AD and amyloid precursor protein, a trans-membrane receptor,is central to the pathogenesis of the disease (Jeong, 2017).Amyloid-beta fragment 25–35 has been reported as the functional neurotoxic domain, widely used bothin vitroandin vivoto induce memory deficits (Millucci et al., 2010; Peters et al., 2016). Consistently, in the present study, i.c.v. injection of amyloid-beta peptide fragment 25–35 aggregate caused memory impairments in mice.
For encapsulation efficiency, we were unable to quantify meloxicam or curcumin in the ultrafiltrate, which means that encapsulation was 100% for all drugs, according to previous studies (Velasques et al., 2018; Nakama, 2020). These results are consistent with the findings of Nakama et al. (2020) who determined that the distribution of curcumin (LCN C and LCN CM) and meloxicam (LCN M and LCN CM) presents type VI distribution profile.
Furthermore, our findings demonstrated that only treatment with meloxicam and curcumin-co-loaded LCN rescued the memory deficit induced by amyloid-beta peptide in the ORT and inhibitory avoidance tests. Interestingly, meloxicam,meloxicam-loaded LCN, and the association of meloxicam+ curcumin and meloxicam + curcumin-co-loaded LCN treatment partially protected against the memory impairment in the inhibitory avoidance test. Taken together, these results suggest that the co-nanoencapsulation potentiates the effects of curcumin and meloxicam alone.Co-encapsulation of curcumin with another drug is a strategy that has already shown promise in other models. Coencapsulation of curcumin and fish oil showed an antioxidant action in human SH-SY5Y cells (Rakotoarisoa et al., 2019) by enhancing the activity of catalase, an antioxidant enzyme(Rakotoarisoa et al., 2019). Indeed, the dual co-encapsulation of curcumin and docosahexaenoic acid (DHA) promoted neuronal survival through enhanced BDNF secretion and increased phosphorylation of CREB, improving neuronal function (Guerzoni et al., 2017).
Curcumin administration has been shown to improve cognitive performance, enhance neurogenesis and restrain inflammation and oxidative stress, which normalize mitochondrial respiration (Kodali et al., 2018; Sharma et al.,2018) and promote neuronal survival. Although this effect was not observed in the present study, curcumin-loaded LCN treatment improved neurocognition in an AD model (Jacques et al., 2013) by regulating the BDNF and Akt/GSK-3β signaling pathways (Hoppe et al., 2013). Indeed, Rakotoarisoa and Angelova (2018) described the promising effects of curcumin encapsulation in several neurodegenerative models. Similar to curcumin, it has been proposed that meloxicam-loaded LCNs reverse the memory impairment induced by amyloid-beta peptide via activation of Na+,K+-ATPase and oxidative stress reduction (Ianiski et al., 2012, 2016).
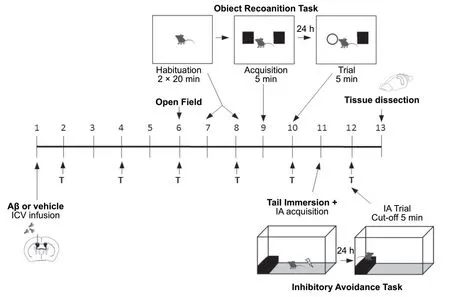
Figure 1 |Schematic representation of the experimental design.
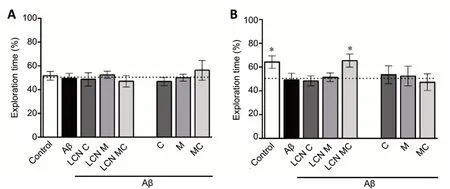
Figure 2 |Effect of different formulations of meloxicam and/or curcumin on amyloid-beta-induced memory impairment in the Object Recognition Test in mice.
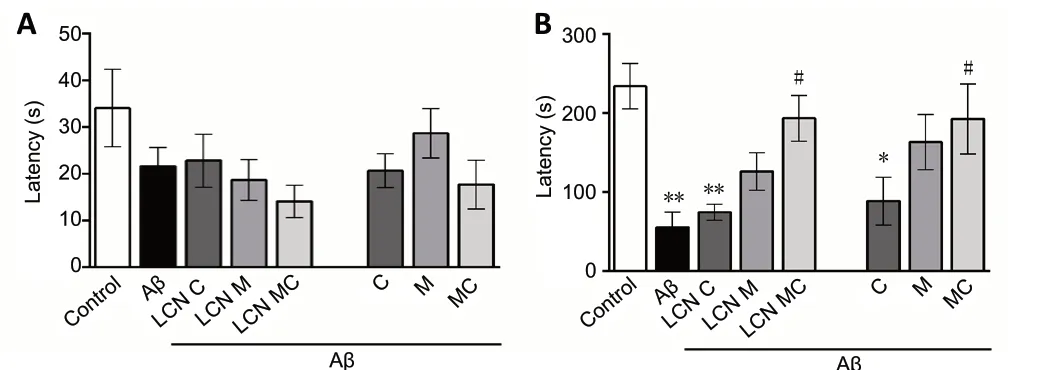
Figure 3 |Effect of different formulations of meloxicam and/or curcumin on amyloid-beta-induced memory impairment in the Inhibitory Avoidance Test in mice.
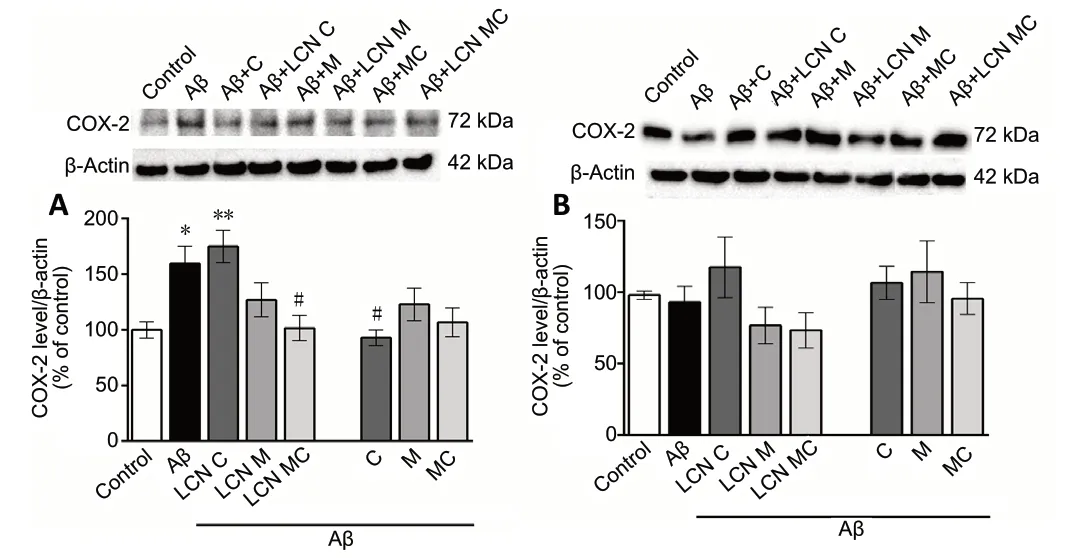
Figure 4 |Effect of different formulations of meloxicam and/or curcumin on COX-2 protein expression levels in prefrontal cortex and hippocampus of amyloid-beta-infused mice.
Curcumin and meloxicam have been used as anti-inflammatory drugs, with important biopharmaceutical limitations. In addition to enabling the administration of these compounds,nanoencapsulation into LCNs may improve their bioavailability and cerebral penetration. Hence, we have nanoencapsulated these two drugs, both isolated and in association. The physical-chemical characterization was adequate and in accordance with other studies involving nanoencapsulated curcumin (Hoppe et al., 2013; Jacques et al., 2013; Velasques et al., 2018), meloxicam (Ianiski et al., 2012, 2016; Villalba et al., 2016) and both (Tuppo and Arias, 2005).
Thus, considering the potential neuroprotective effects of both drugs and the need to increase their bioavailability and targeting to the central nervous system, their incorporation into nanocapsules is a suitable approach. Curcumin presents a below 30% bioavailability and nanoencapsulation increases this parameter (Jacques et al., 2013; Villalba et al., 2016).Recently, rhodamine-labeled LCNs were found inside the vessels of the brain parenchyma (Rodrigues et al., 2016)after oral and intravenous administration. With the same nanosystem used in this work, Carreño et al. (2016) showed that quetiapine, an antipsychotic, has an increased cerebral adsorption in comparison to the non-encapsulated drug.These works highlight the importance of LCNs as a drug shuttle through the blood-brain barrier.
Although different mechanisms appear to be involved in the pathogenesis of AD, it has been demonstrated that neuroinflammation accounts for its progression. In aging and neurodegenerative disease, microglia become steadily more reactive and increase the release of pro-inflammatory cytokines that could induce neuronal dysfunction (Heneka et al., 2015; Sen et al., 2017). Activation of glial cells induced by accumulation of amyloid-beta peptide further damages neurons by producing inflammatory mediators, such as nitric oxide, interleukin-1, and interleukin-6, and transcription factor nuclear factor-kappaB (Hensley, 2010; Heneka et al., 2015;Sen et al., 2017).
As mentioned previously, inflammatory process has a key role in neuronal dysfunction. Once under the physiological condition, the expression of COX-2 enzyme in neurons is related to the possible involvement of synaptic activity(Minghetti, 2004; Heneka et al., 2015; Mhillaj et al., 2018;Velasques et al., 2018). COX-2 is selectively expressed by glutamatergic neurons localized in the cerebral cortex and many other regions of the adult brain, suggesting that this enzyme might be directly involved in the regulation of postsynaptic signaling and plasticity mechanisms underlying learning and memory processes (Minghetti, 2004; Mhillay et al., 2018).
Although COX-2 expression varies with AD stage(Minghetti, 2004), pharmacological evidence suggests that neuroinflammation is associated with an increased expression of COX-2 in both AD patients and animal models (Minghetti,2004; Nazem et al., 2015; Mhillay et al., 2018). Considerable evidence suggests that increased COX-2 activity is due to both direct and indirect activation of microglia by amyloid-beta peptide (Minghetti, 2004; Nazem et al., 2015; Mhillaj et al.,2018)
In agreement with this, our findings indicated that the memory deficit induced by amyloid-beta peptide was accompanied by an increase in the cortical COX-2 levels.Indeed, the expression of COX-2, an enzyme that mediates the inflammatory response in the brain, increased in neurons during the early phase of AD (Nazem et al., 2015), suggesting that activation of COX-2 could be involved in the development of neuroinflammation and progression of AD by the high production of prostaglandins E2 and reactive oxygen species(Hoozemans and O’Banion, 2005; Nazem et al., 2015).
It is well known that neuroinflammation plays a key role in the development of neuronal dysfunction, therefore, NSAIDs such as meloxicam are being used for their neuroprotective action in different neurodegeneration models (Haile et al.,2016). Thus, in AD, meloxicam could promote the inhibition of COX-2 enzyme, reducing the inflammation, neuronal loss and behavioral changes caused by amyloid-beta peptide(Azizi et al., 2015; Faraji and Wipf, 2009). In fact, meloxicam treatment has beneficial effects in AD patients and animal models (Ianiski et al., 2012, 2016). Meloxicam can reduce astrocyte proliferation by decreasing the expression of GFAP in the hippocampus and cortex in a rat model of AD (Mao et al., 2010). There is evidence that COX-2 involvement in synaptic plasticity has been demonstrated using COX inhibitors inin vivoandin vitromodels (Minghetti, 2004; Mhilay et al.,2018). Indeed, some findings indicate that COX-2 is a required biochemical component for the consolidation of memory(Teather et al., 2002). Thus, COX-2 inhibition, by reducing neuroinflammation, might prevent morphological as well as the phenotypical alterations of microglia and probably increases amyloid-beta peptide clearance (Gulyaeva et al.,2017). In our study, treatment with meloxicam and curcumin co-encapsulated (LCN MC) fully protected against the increase in cortical COX-2 levels induced by amyloid-beta peptide,suggesting that the anti-inflammatory action is involved in the neuroprotective effect of meloxicam and curcumin.
Moreover, studies have found that curcumin can inhibit GSK- 3β mediated presenilin-1 activity and reduce the generation of amyloid-beta peptide, thus inhibiting the inflammatory response (Muglikar et al., 2013; Shi et al.,2015). In this context, curcumin can also decrease the levels of phosphorylated tau (p-tau) by regulating BCL2-Associated Athanogene 2 (BAG2) activity, and should play a role in the treatment of AD (Zarghi and Arfaei, 2011; Muglikar et al., 2013). However, despite all the beneficial effect in the treatment with LCN CM, the mechanisms involved in the pharmacological effects of these two drugs together are not fully understood. Therefore, further studies are needed.
Conclusion
The present study showed the effectiveness of meloxicam+ curcumin co-encapsulation in lipid core nanocapsules in improving the memory and reducing COX-2 expression in amyloid-beta-infused mice. Thus, our findings suggest that the neuroprotective potential of meloxicam and curcumin co-nanoencapsulation is associated with cortical COX-2 modulation. Although additional studies are necessary to determine the other mechanisms involved in the effect of this nanoformulation, meloxicam and curcumin conanoencapsulation may have potential for rescue or delaying the progression of cognitive impairment associated with AD.
Acknowledgments:We thank Cristina Wayne Nogueira for critically reading the manuscript and collaboration.
Author contributions:SP designed the study. MEZG was a major contributor in writing the manuscript, and analyzed and interpreted data.ASPS, ELA and MCMG contributed to animal experiments. KAN and RBDS synthesized and characterized LCNs. CBQ, DSÁ, CL and SEH reviewed the manuscript. SP analyzed and interpreted data. All authors contributed to the planning of the experimental protocol, read and approved the final manuscript.
Conflicts of interest:The authors declare that there are no conflicts of interest.Financial support:This study was supported by the Rio Grande do Sul Science Foundation (FAPERGS), grants # 16/2551-0000207-0 and 16/0526-5 (PRONUPEQ) (to SP), National Counsel of Technological and Scientific Development (CNPq) (Universal grants # 423435/2016-7 and 460122/2014-2) and for student scholarships (to CL, SEH and DSÁ).
Institutional review board statement:The experiments were approved by the Committee on Care and Use of Experimental Animal Resources, at the Federal University of Pampa, Brazil (Protocol 02-2015) on April 16,2015. All experiments, maintenance and manipulation of the animals were done according to the guidelines of CONCEA (National Council for Control of Animal Experimentation) law #11.794, of 08.10.2008 and normative resolution # 30, of 02.02.2016. All efforts were made to minimize suffering of the animals and to reduce the number of animals used in the experiments.
Copyright license agreement:The Copyright License Agreement has been signed by all authors before publication.
Data sharing statement:Datasets analyzed during the current study are available from the corresponding author on reasonable request.
Plagiarism check:Checked twice by iThenticate.
Peer review:Externally peer reviewed.
Open access statement:This is an open access journal, and articles are distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 License, which allows others to remix,tweak, and build upon the work non-commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.

- 中国神经再生研究(英文版)的其它文章
- Patterning inconsistencies restrict the true potential of dopaminergic neurons derived from human induced pluripotent stem cells
- Efficacy of epothilones in central nervous system trauma treatment: what has age got to do with it?
- Association between plasma immunoproteasome and 90-day prognosis after first-ever ischemic stroke
- Melatonin ameliorates microvessel abnormalities in the cerebral cortex and hippocampus in a rat model of Alzheimer’s disease
- Regulated upon activation, normal T cell expressed and secreted (RANTES) levels in the peripheral blood of patients with Alzheimer’s disease
- Comparison of cerebral activation between motor execution and motor imagery of self-feeding activity