
目标物直径对笼养川金丝猴和北豚尾猴抓握行为的影响
2021-10-25 04:26胡淑静刘昕晨赵大鹏
天津师范大学学报(自然科学版) 2021年5期
胡淑静,张 生,王 媛,刘昕晨,赵大鹏
(1.天津师范大学生命科学学院,天津300387;2.天津动物园,天津300381;3.北京野生动物园,北京100081)
抓握行为在四足动物的日常行为中普遍存在[1].与其他哺乳动物相比,灵长类动物具有可抓握的手部结构,能够更好地抓握食物、支撑物等,并可以有效完成更为复杂的抓握行为,从而更好地适应生存环境[1-2].灵长类的手部抓握方式在一定程度上可以代表物种的进化地位,比较研究不同的灵长类抓握行为特征对于理解人类手部进化具有重要意义[3].关于灵长类的早期研究中,Napier[4]依据抓握目的将人类手部抓握方式划分为2种类型:力量型抓握(power grip)和精确型抓握(precision grip).力量型抓握是将物体固定于手掌与除大拇指外的4根手指之间,手掌与四指朝同一方向弯曲,大拇指朝相反方向将物体固定;精确型抓握是将物体固定于一根手指和大拇指的指尖.二者差别主要体现在:①大拇指发挥的作用不同,在力量型抓握中大拇指主要起辅助作用,在精确型抓握中大拇指主要起固定手中物体的作用;②抓握对象不同,力量型多用于抓握体积较大的物体,精确型常用于抓握体积较小的物体.
一些非人灵长类虽然无法做到像人类那样典型的拇指与其他手指指腹间完全相对,但对于小型物体依然可以应用近似的精确型抓握方式[5].如黑猩猩(Pan troglodytes)在使用锤子砸开坚果时使用一只手拿锤子,另一只手会用5种方式固定坚果,其中就包含2种精确型抓握[5],而且它们在成年后会逐渐掌握精确型抓握的能力,虽然在8岁前这种能力并不常见[6].Hopkins等[7]、Jones-Engel等[8]也发现黑猩猩会使用精确型抓握方式.Marzke等[9]对39只黑猩猩进行测试,发现它们在对10种食物进行抓握时共使用了13种抓握方式,其中一种为新型抓握方式,拇指在抓握过程中仅发挥辅助作用.
迄今关于非人灵长类物种抓握行为的研究主要关注新大陆猴,如卷尾猴(Cebus apella)[10-11]、棕头蜘蛛猴(Ateles fusciceps rufiventris)[12]、倭狐猴(Microcebus murinus)[13]、白毛卷尾猴(Sapajus spp.)[14]和类人猿(如黑猩猩)[6-15],旧大陆猴物种的手部抓握行为研究相对较少,研究物种主要有普通猕猴(Macaca mulatta)[16]和食蟹猕猴(Macaca fascicularis)[17].关于目标物直径对于灵长类抓握行为类型影响的研究则更为有限[18].本研究创新性地选择主要营树栖生活的川金丝猴(Rhinopithecus roxellana)与主要营地栖生活的北豚尾猴(Macaca leonina)这2种生活方式不同的旧大陆猴濒危物种作为研究对象,应用不同直径的人工实验测试物,阐明这2个物种抓握方式的种类与特点,分析目标物直径对两物种抓握行为种类表达的影响,关注物种间的表达差异,为灵长类手部抓握行为学研究及人类手部进化研究提供实验依据.
1 研究对象与方法
1.1 研究对象
川金丝猴12只,饲养于北京大兴野生动物园动物繁殖中心,含8只成年和4只未成年个体,分别为6只雄性和6只雌性.个体分开饲养在6个不同笼舍内,数量分别为1、3、4、1、2、1只.每个笼舍结构相同,均包括1个室内和1个室外活动区域,区域之间以狭窄通道(4.5 m×3.2 m×5.0 m)相连接.室外活动区域为鸟笼状笼舍(底部直径为3.0 m,高度为5.0 m),个体昼间主要在室外活动区域,每日喂食2次.
北豚尾猴6只,饲养于天津动物园猿猴馆,含2只成年和4只未成年个体,分别为4只雄性和2只雌性.所有个体饲养在同一笼舍内,笼舍包括1个室内(4.8 m×3.2 m×7.0 m)和1个室外活动区域(7.1 m×5.6 m×3.8 m),区域之间以人工控制开关的小窗相连接,每日喂食1次.
1.2 数据收集与分析
数据收集起止时间:川金丝猴为2018年6月19日—2018年8月23日;北豚尾猴为2018年1月23日—2018年6月11日和2018年11月27日—2019年7月4日.在收集数据之前进行有效个体识别.在实验正式开始之前,对每个物种的每个个体至少开展3 d的预观察实验,从而有效减少研究人员对研究个体行为表达的潜在影响作用.
选取2个物种食谱中均包含的自制窝头,将其制成直径分别为1、3和5 cm的人工实验测试物,分别称为小号、中号和大号测试物.考虑到北豚尾猴研究个体样本量较少,另选择玉米粒和葵花籽作为北豚尾猴的天然食物测试物.随机测量100粒玉米粒,长径平均值为(11.84±1.49)mm,短径平均值为(8.91±1.48)mm;随机测量100粒葵花籽,长径平均值为(24.07±1.44)mm,短径平均值为(9.68±0.83)mm.
每次收集数据前,先将少量人工实验测试物摆放于笼舍内外的地面上靠近围栏约15 cm处,既方便拍摄、记录相关数据又不影响个体正常抓取测试物.通过数码摄像机(Panasonic HC-W570M)跟踪拍摄所有个体的每次有效抓握行为,每次实验中提供的测试物种类随机.每日的数据收集时间集中于11:00—14:00.个体用手将测试物抓离地面即记为1次抓握数据;若测试过程中拍摄到某一个体单手在地面连续捡拾多个测试物后才离开地面,个体抓取第1个测试物时所采用的手部姿势记为有效抓握数据.对于每一个测试物,收集有效抓握行为数据量均为100个/每个实验个体.川金丝猴有效抓握数据共计3 600次,北豚尾猴有效抓握数据共计3 000次.
对所有个体的抓握行为视频数据逐帧回放分析,逐一记录个体进行每次抓握所采用的抓握方式并分类汇总.关于川金丝猴和北豚尾猴每一种抓握方式定义中涉及的手部结构主要参照普通猕猴手掌各部位命名方式[16],将川金丝猴和北豚尾猴的抓握方式进行归类比对.应用Friedman检验和Wilcoxon检验分析不同测试物任务间所表现出的抓握方式种类数差异是否具有统计学意义.应用SPSS 26.0完成所有数据统计分析,统计学意义定义为P≤0.05.
2 结果与分析
本研究中,川金丝猴和北豚尾猴在实验期间共表现出15种抓握方式,具体特点如表1所示.其中:川金丝猴共表现出13种抓握方式,北豚尾猴共表现出12种抓握方式.川金丝猴和北豚尾猴两物种共表现出10种相同的抓握方式,其中:4种为力量型,6种为精确型.

表1 研究物种的手部抓握方式Tab.1 Grasping patterns of study species
2.1 川金丝猴抓握方式
川金丝猴在大号测试物任务中共表现出9种抓握方式(Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅵ、Ⅷ、Ⅸ、Ⅻ),其中使用频率最高的是第Ⅳ种抓握方式,整体所占比例为29.00%;在中号测试物任务中共表现出10种抓握方式(Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅵ、Ⅷ、Ⅸ、Ⅹ、Ⅺ、Ⅻ),其中使用频率最高的是第Ⅱ种抓握方式,整体所占比例为58.58%;在小号测试物任务中共表现出5种抓握方式(Ⅶ、Ⅷ、Ⅹ、Ⅺ、す),其中使用频率最高的是第Ⅹ种抓握方式,整体所占比例为84.25%.
川金丝猴在不同测试物任务间表现出的抓握方式种类数存在显著差异(Friedman:χ2=13.535,df=2,P=0.001),如图1所示.
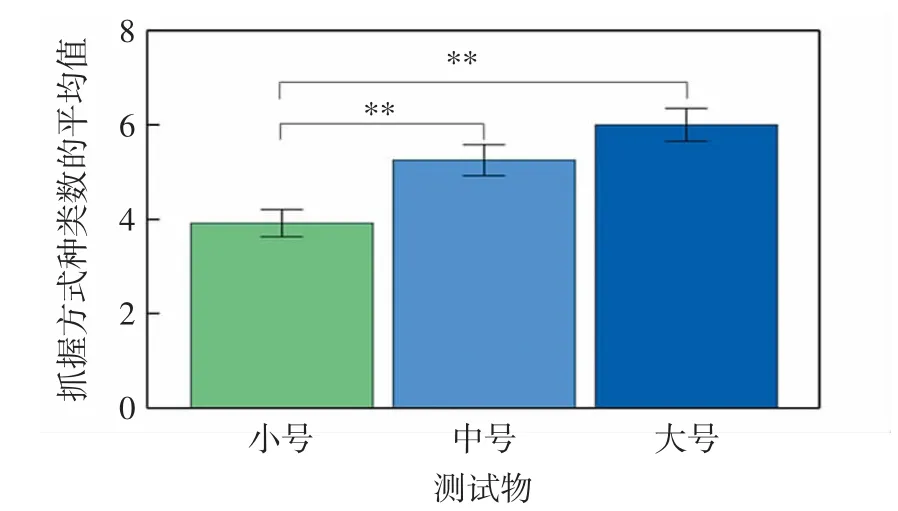
图1 不同测试物任务中川金丝猴个体所采用抓握方式的种类数Fig.1 Number of grasping patterns across tasks in Sichuan snub-nosed monkeys
由图1可以看出,川金丝猴在大号测试物任务中表现出的抓握方式种类数与其在中号测试物任务中的种类数之间无显著差异(Wilcoxon:Z=-1.305,P=0.192),但显著多于其在小号测试物任务中的种类数(Wilcoxon:Z=-2.825,P=0.005),在中号测试物任务中的种类数显著多于其在小号测试物任务中的种类数(Wilcoxon:Z=-2.724,P=0.006).
川金丝猴在3项测试物任务中仅表现出1种相同的抓握方式(Ⅷ).在大号与中号测试物任务中共表现出8种相同的抓握方式(Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅵ、Ⅷ、Ⅸ、Ⅻ),其中6种方式所占百分比呈现显著差异(Ⅰ、Ⅲ、Ⅳ、Ⅵ、Ⅷ、Ⅻ).川金丝猴在大号测试物任务与小号测试物任务中共表现出1种相同的抓握方式(Ⅷ),其所占百分比无显著差异,如表2所示.川金丝猴在中号与小号测试物任务中共表现出3种相同的抓握方式(Ⅷ、Ⅹ、Ⅺ),其所占百分比均存在显著差异.
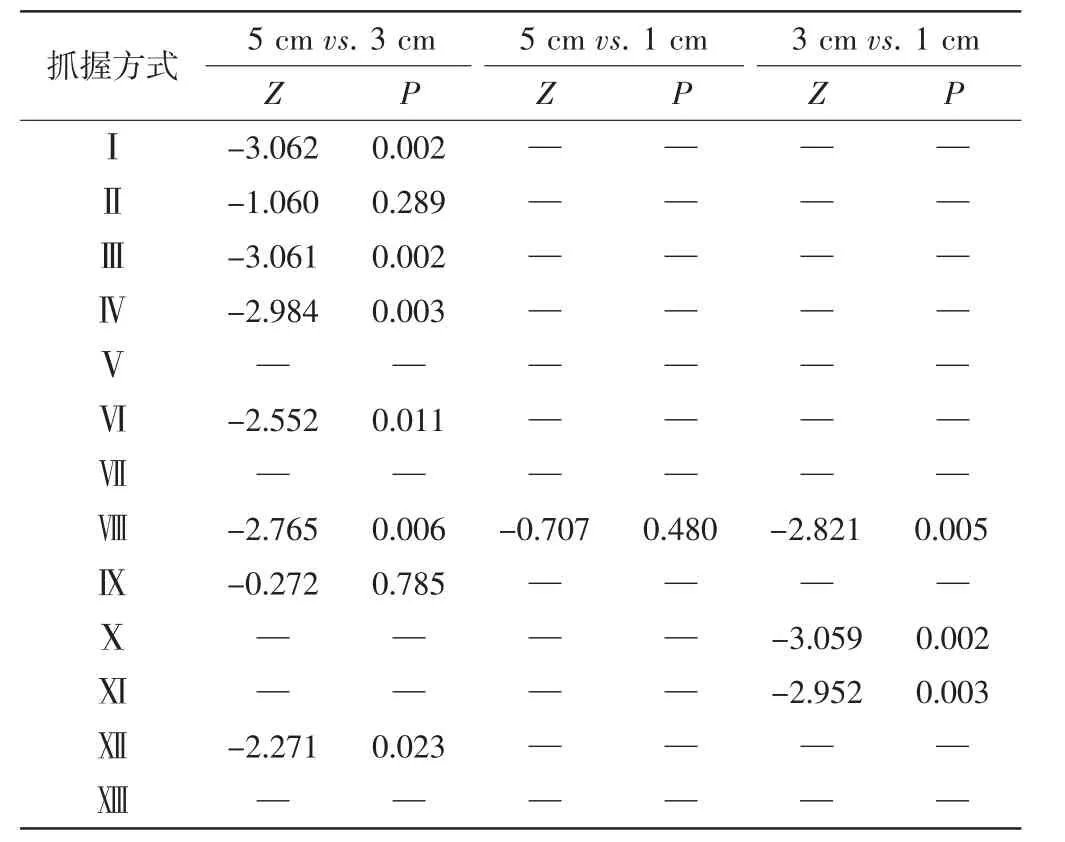
表2 两两测试物任务间川金丝猴共有抓握方式所占百分比差异Tab.2 Percentage difference of the common grasping patterns between two tasks in Sichuan snub-nosed monkeys
2.2 北豚尾猴抓握方式
北豚尾猴在大号测试物任务中共表现出9种抓握方式(Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅵ、Ⅻ、い、こ),其中使用频率最高的是第Ⅰ种,整体所占比例为60.33%;在中号测试物任务中共表现出8种抓握方式(Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅵ、Ⅶ、Ⅻ、こ),其中使用频率最高的是第Ⅰ种,整体所占比例为33.00%;在小号测试物任务中共表现出4种抓握方式(Ⅶ、Ⅹ、Ⅺ、こ),其中使用频率最高的是第Ⅺ种,整体所占比例为71.67%;在玉米粒测试任务中共表现出4种抓握方式(Ⅶ、Ⅹ、Ⅺ、こ),其中使用频率最高的是第Ⅺ种,整体所占比例为90.17%;在葵花籽测试任务中共表现出3种抓握方式(Ⅶ、Ⅹ、Ⅺ),其中使用频率最高的是第Ⅺ种,整体所占比例为76.83%.
北豚尾猴在不同测试物任务中所表现出的抓握方式种类数存在显著差异(Friedman:χ2=12.857,df=4,P=0.012),如图2所示.北豚尾猴在玉米粒测试任务中表现出的抓握方式种类数显著低于其在大号测试物任务(Wilcoxon:Z=-2.032,P=0.042)和中号测试物任务(Wilcoxon:Z=-2.214,P=0.027)中表现出的抓握方式种类数.其他两两测试任务间表现出的抓握方式种类数均无显著差异(Wilcoxon:allP>0.05).
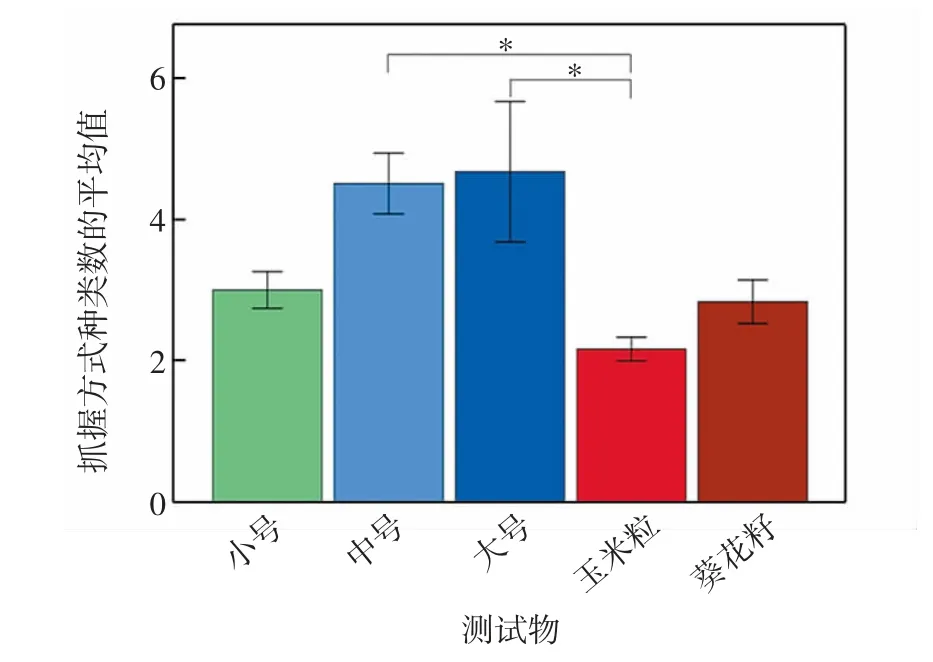
图2 不同测试任务中北豚尾猴个体所采用抓握方式的种类数Fig.2 Number of grasp patterns across tasks in northern pig-tailed macaques
北豚尾猴在5项测试物任务中未表现出任一相同的抓握方式.北豚尾猴在大号与中号测试物任务中共表现出7种相同的抓握方式(Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅵ、Ⅻ、こ),其中1种抓握方式(Ⅻ)所占百分比呈现显著差异,如表3所示.

表3 两两测试物任务间北豚尾猴共有抓握方式所占百分比差异Tab.3 Percentage difference of the common grasping patterns between two tasks in northern pig-tailed macaques
北豚尾猴在大号与小号测试物任务中共表现出1种相同的抓握方式(こ),其所占百分比无显著差异.北豚尾猴在中号测试物任务与小号测试物任务中共表现出2种相同的抓握方式(Ⅶ、こ),其所占百分比均无显著差异.北豚尾猴在大号测试物任务与玉米粒测试物任务中共表现出1种相同的抓握方式(こ),其所占百分比无显著差异.北豚尾猴在大号测试物任务与玉米粒测试物任务中无相同的抓握方式.北豚尾猴在中号测试物任务与玉米粒测试物任务中共表现出2种相同的抓握方式(Ⅶ、こ),其所占百分比均无显著差异.北豚尾猴在中号测试物任务与葵花籽测试物任务中共表现出1种相同的抓握方式(Ⅶ),其所占百分比无显著差异.北豚尾猴在小号测试物任务与玉米粒测试物任务中共表现出4种相同的抓握方式(Ⅶ、Ⅹ、Ⅺ、こ),其所占百分比均无显著差异.北豚尾猴在小号测试物任务与葵花籽测试物任务中共表现出3种相同的抓握方式(Ⅶ、Ⅹ、Ⅺ),其所占百分比均无显著差异.北豚尾猴在玉米粒测试物任务与葵花籽测试物任务中共表现出3种相同的抓握方式(Ⅶ、Ⅹ、Ⅺ),其中2种抓握方式(Ⅹ、Ⅺ)所占百分比呈现显著差异.
2.3 物种间比较
比较川金丝猴和北豚尾猴的抓握方式,结果如表4所示.
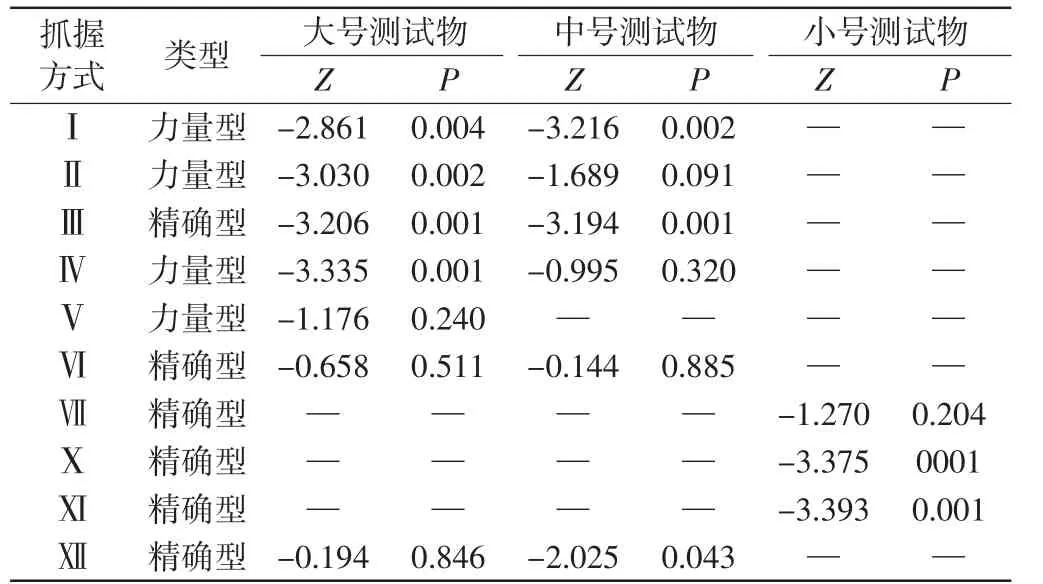
表4 两物种面对不同测试物所表现出相同抓握方式的百分比差异Tab.4 Interspecific differences on percentages of the common grasping pattern between two species
2个物种在面对大号测试物时表现出7种相同的抓握方式(Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅵ、Ⅻ)(力量型:57.14%;精确型:42.86%),其中4种方式(力量型:75.00%;精确型:25.00%)在抓握频次百分比方面存在显著差异;在面对中号测试物时表现出6种相同的抓握方式(Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅵ、Ⅻ)(力量型:50.00%;精确型:50.00%),其中3种方式(力量型:33.33%;精确型:66.67%)在抓握频次百分比方面存在显著差异;在面对小号测试物时表现出3种相同的抓握方式(Ⅶ、Ⅹ、Ⅺ)(精确型:100%),其中2种方式在抓握频次百分比方面存在显著差异;在面对大号和中号测试物时表现出6种相同的抓握方式,而在面对小号测试物时表现出完全不同于前者的3种相同的抓握方式.
随着测试物直径的减小,川金丝猴和北豚尾猴两物种所呈现的共有抓握方式的种类数相应减少,如表4所示.两物种共有精确型抓握方式种类数在共有抓握方式总种类数中所占比例相应增加,两物种抓握频次百分比方面存在显著差异的精确型共有抓握方式种类数在抓握频次百分比方面存在显著差异的共有抓握方式总种类数中所占比例相应增加.
在3项人工测试物任务中,川金丝猴和北豚尾猴所表现出的抓握类型种类数均不存在显著物种间差异(大号测试物:Z=-1.631,P=0.103;中号测试物:Z=-1.192,P=0.233;小号测试物:Z=-1.914,P=0.056),如图3所示.
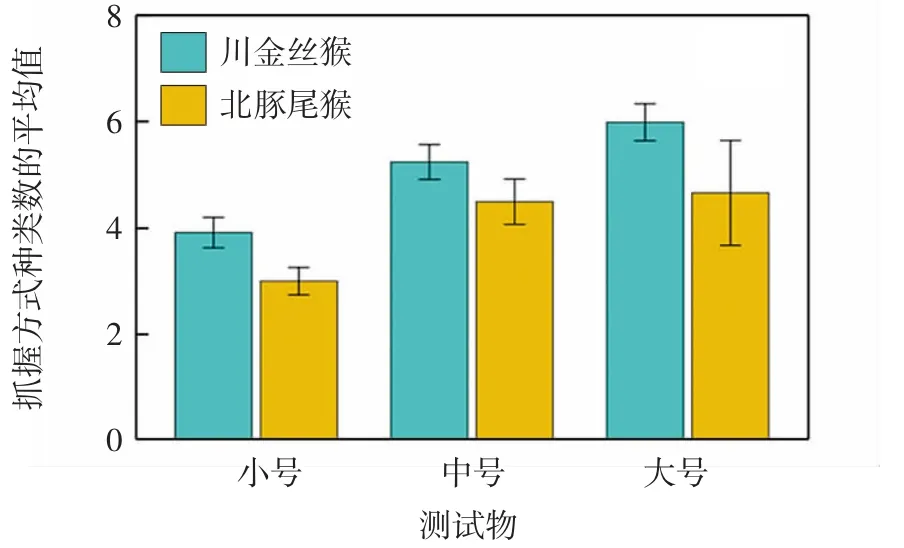
图3 不同测试球实验任务中抓握方式种类数的物种间差异Fig.3 Species differences in the number of grip patterns across tasks
3 讨论与结论
本研究测试了川金丝猴和北豚尾猴对不同直径测试物的抓握方式,两物种共表现出15种抓握方式,其中川金丝猴共表现出13种方式,北豚尾猴共表现出12种方式,两物种共表现出10种相同的方式,这在一定程度上说明虽然2个研究物种手部形态存在一定的差异,但在抓握方式上还是表现出了部分一致性.其中,在大号测试物任务中共有7种相同的抓握方式(Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅵ、Ⅻ),共有方式Ⅰ与普通猕猴(Macaca mulatta)自然情况下抓握较大物体时的某种抓握方式相同[16];在中号测试物任务中共有6种相同的抓握方式(Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅵ、Ⅻ),共有方式Ⅳ与普通猕猴自然情况下抓握中等大小物体时的某种方式相同[21];在小号测试物任务中共有3种相同的抓握方式(Ⅶ、Ⅹ、Ⅺ),共有方式Ⅹ和Ⅺ与人类(Homo sapiens)、倭黑猩猩(Pan paniscus)和普通猕猴抓握较小体积食物(如草叶、水果片)时的某些抓握方式相同[16].
川金丝猴和北豚尾猴在不同测试物任务间所表现出的抓握方式种类数存在显著差异,其中,川金丝猴在大号测试物任务中的种类数与其在中号测试球任务中的种类数之间无显著差异,但二者显著多于其在小号测试物任务中的种类数.北豚尾猴在玉米粒测试任务中表现出的抓握方式种类数显著低于其在大号测试物任务、中号测试物任务中的种类数,其他两两测试任务间均无显著差异.这在一定程度上说明测试物直径大小影响两物种抓握方式的多样化[16].随着测试物直径的减小,川金丝猴和北豚尾猴共有抓握方式的种类数减少,两物种共有精确型抓握方式种类数在共有抓握方式总种类数中所占比例增加,抓握频次百分比方面存在显著差异的精确型共有抓握方式种类数在抓握频次百分比方面存在显著差异的共有抓握方式总种类数中所占比例相应增加,这在一定程度上说明直径较小的实验测试物会进一步激发两物种精确型抓握方式的表达强度.
早期关于人类抓握行为的相关研究已显示出将测试物体积进行规范化的重要性[19-21].Pouydebat等[18]对人类和其他几种非人灵长类物种(黑猩猩、婆罗洲猩猩(Pongo pygmaeus)、日本猕猴(Macaca fuscata)、几内亚狒狒(Papio papio)、卷尾猴(Cebus apella))开展研究,发现测试物大小会影响动物个体抓握方式的使用,在抓握大小不同的食物时,不同的灵长类表现出对不同抓握方式的偏好性.本研究中,川金丝猴和北豚尾猴在大号测试物任务中使用频率最高的分别是第Ⅳ种和第Ⅰ种抓握方式,均为力量型抓握方式;在中号测试物任务中两物种使用频率最高的分别是第Ⅱ种和第Ⅰ种抓握方式,均为力量型抓握方式;在小号测试物任务中两物种使用频率最高的分别是第Ⅹ种和第Ⅺ种抓握方式,均为精确型抓握方式.由此可见,2个旧大陆猴物种在不同测试物任务中表现出了抓握方式的偏好性和抓握类型的一致性.
综上所述,本研究是国内关于灵长类物种抓握行为的研究,研究中对目标测试物的形状和直径进行了统一规范,从而为今后灵长类抓握方式的比较研究提供了标准化参考.在今后的相关研究中,建议考虑选用更多不同形状尺寸、不同颜色图案的人工测试物和天然测试物,开展野生与笼养环境下灵长类物种抓握行为的综合比较研究,分析年龄性别的组差异,为进一步探寻人类手部行为进化起源提供更多的基础性研究信息.
猜你喜欢
云南画报(2021年10期)2021-11-24
东方少年·布老虎画刊(2021年1期)2021-06-22
小哥白尼(野生动物)(2020年4期)2020-07-27
智富时代(2019年8期)2019-09-23
智富时代(2019年8期)2019-09-23
小天使·一年级语数英综合(2019年8期)2019-08-27
学生天地(2018年36期)2019-01-28
摄影之友(影像视觉)(2017年1期)2017-07-18
电子制作(2017年20期)2017-04-26
小学阅读指南·低年级版(2016年6期)2016-05-14
