
性比自调节:捕食性天敌异色瓢虫爆发成因*
2021-10-21 08:10王雪付雪赛序岐王佳佳于慧龄叶乐夫
中山大学学报(自然科学版)(中英文) 2021年5期
王雪,付雪,,赛序岐,王佳佳,于慧龄,叶乐夫
1. 东北农业大学农学院,黑龙江哈尔滨 150030
2. 黑龙江大学现代农业与生态环境学院,黑龙江哈尔滨 150080
异色瓢虫Harmonia axyridis是多种害虫的优良捕食性天敌,在生物防治上发挥着重大作用。然而,由于其极强的生存能力和竞争能力,异色瓢虫在作为天敌引入后, 可能会发展为入侵物种[1-2]。在入侵地,异色瓢虫的爆发[3]极大地威胁了当地其他天敌种群[4-5]。研究表明,异色瓢虫爆发机制的外部因素为其猎物的丰富度,如众多种类的蚜虫[6],内部因素可能为其超强的适应力及两性繁殖的特点。而在种群繁殖中,成虫性比的变化与最佳性比的关系将影响到昆虫种群的数量发展。
自然界中,昆虫种群受温度、扩散能力及种群竞争强度等多种因素影响,性比总是处于波动状态。温度升高时,雌雄个体间死亡率不同[7-8],导致种群性比偏倚[9]。在雄性个体扩散能力弱于雌性的许多分类学类目中,雌性个体比例高[10],而且根据当地雄性竞争程度,许多种群会调整后代性比[11]。当雌性个体受到营养条件差等环境压力时,其后代雄性比例也会增加[12]。昆虫种群的性比变化对成虫及子代都将产生连锁影响,包括交配行为、产卵量及个体寿命。
研究表明,随着雄性个体比例增加,未交配雌性个体比例将下降[13],当性比轻微偏向雄性时(雄雌比接近2∶1),挑衅、交配与防御行为频次增加,但当雄虫比例继续增加时,这些行为的频次下降,当雄性比例非常高时,雄虫的选择性降低,交配率下降[14]。就产卵量而言,不同昆虫种群的最佳性比不同,当白魔按蚊Anopheles alhirnanus与烟蚜夜蛾(别称绿棉铃虫)Heliothis virescens种群中雌性个体比例较高时,产卵量最大[15-16]。而在斜纹夜蛾Spodoptera litura与种群密度较高的葱蝇Delia antiqua种群中,升高雌性比例,总产卵量减少,产卵率下降[17-18]。在对美洲棉铃虫Heliothis zea的研究中也得出了相似结论,即种群中雄性个体比例增加时,产卵量增加[19]。然而,虽然日产卵量增加,雌性成虫寿命缩短,结果总产卵量不受影响[20]。相应地,种群性比对寿命也有影响,因为交配会使得营养更多流向生殖系统(如卵子发生和成熟),使得雌性寿命显著缩短[21-24]。
我们之前的研究表明,亚致死剂量杀虫剂将使异色瓢虫发育历期延长,蛹重下降[25],进而可能影响到种群的性比,当昆虫种群性比变化时,将对种群当代及子代的许多生物学指标造成影响。探究这种影响不仅能对异色瓢虫田间释放防控害虫及人工饲养工作起到指导作用,而且可能解释异色瓢虫在各类农林生态系统中爆发成因。本试验在实验室条件下,设置了异色瓢虫5 个性比组合,旨在探究性比对当代及子代的影响。尤其是,异色瓢虫这种捕食性昆虫能否在受到扰动而性比失衡后迅速做出响应,将性比调整至一个合适的水平以保证子代的繁殖力?
1 材料与方法
1.1 田间种群调查
2018年9月和2020年9~10月,于东北农业大学向阳农场、东北农业大学校园及黑龙江大学共调查8个异色瓢虫种群,每次调查取样方法为五点取样法。
1.2 供试虫源
大豆蚜为室内大豆苗(东农46) 饲养多年且未使用任何农药的品系。异色瓢虫采于东北农业大学试验田向阳农场,在人工气候箱内孵化并以大豆蚜饲养。饲养条件为:温度(25±1)℃,RH 70%,光周期14 L∶10 D。
1.3 仪器
HPG-280HX 人工气候箱; MOTIC SMZ-140 Series 连续变倍体解剖镜。
1.4 试验方法
1.4.1 成虫性别配比从东北农业大学向阳农场采集回来的异色瓢虫,在实验室培养箱内以大豆蚜饲养,交配产卵后,取新孵化的幼虫继续饲养,直至成虫。在解剖镜下观察成虫,确定雌雄。按雌雄比1∶9,3∶7,5∶5,7∶3,9∶1 的比例,在直径为15 cm 的培养皿中放置10头成虫,每天提供新鲜的充足(预实验) 的大豆蚜并更换培养皿内滤纸,每个成虫性别配比组合设5个重复。
1.4.2 成虫与幼虫观测自成虫产卵后,将卵块取出,移至新的培养皿中培养箱内饲养直至孵化,随机选取健康的新孵幼虫15 头,置于有充足大豆蚜的大豆叶片上,放入干净的培养皿中单头饲养,每天提供充足的大豆蚜并更换滤纸,设5 个重复。每天于8:00,14:00,20:00观察并记录幼虫龄期。待饲养至成虫后,在解剖镜下观察,确定雌雄并记录。用于配对的成虫饲养至死亡。
1.5 数据处理与分析
以上数据结果均采用SPSS 23. 0 数据处理软件进行统计分析,用Excel 2016作图。采用单因素方差分析LSD 法分析不同性别配比的成虫产卵量及幼虫发育历期。对雌虫寿命、孵化率、子代存活率以及雌率进行非参数检验,并进行显著性检验(P<0. 05)。雌虫寿命采用线性回归分析拟合。

每雌日增殖率=(每雌产卵量×孵化率×子代存活率×子代雌率)/雌虫世代历期
每雌日增殖率,该参数可衡量昆虫种群单位时间内繁殖的下一代的雌虫数,它能用于比较种群下一代的增殖潜力。
2 结 果
2.1 自然种群雌性比例
3 个采样地调查的8 个自然种群中,自然种群中样本量为20~518 头,雌率为0. 45~0. 66;均值±标准误:0. 51±0. 02。除两个种群抽样量偏小,雌性比例为0. 45,0. 46,其他雌性比例几乎都超过了0. 50。调查结果表明,在不曾施用杀虫剂田块或校园中, 异色瓢虫雌性比例为(51±2)%(见图1)。
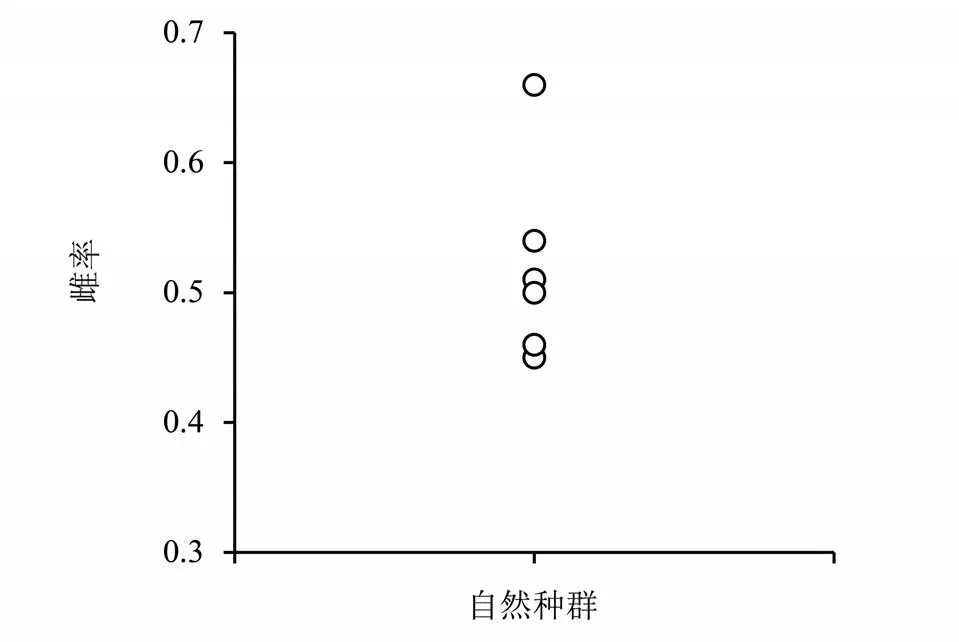
图1 自然种群雌虫比率Fig. 1 Female adult proportion of field populations
2.2 不同性比人工种群增殖力
我们建立了由10头异色瓢虫组成的人工种群。
1) 单雌产卵量,雌雄数量相等时最大,显著高于其他雌雄比例人工种群(F= 12. 864;df1= 3;df2= 16; 与5♀处理相比的P值:P3♀, P7♀<0. 001;P9♀= 0. 006 )。(F为方差分析中的F值,df1为处理自由度,df2为重复自由度,下同)。
2) 总体上, 雌性越多, 产卵量越大(F=29. 484;df1= 3;df2= 16;9♀与7♀处理相比P9♀-7♀=0. 003,9♀与5♀处理相比P9F-5F=0. 005,3♀与5♀,7♀,9♀处理相比P3♀-5♀,7♀,9♀<0. 001),虽然5♀与7♀人工种群的产卵量无显著差异。
3) 幼蛹历期,5♀人工种群子代发育历期显著短于其他种群(16 d),7♀人工种群的最长(17 d)(F=3. 828;df1=3;df2= 16;5♀与3♀处理相比P5♀-3♀= 0. 043;5♀与7♀处理相比P5♀-7♀= 0. 006)。
4) 雌虫寿命随雄性个体比例减少而增加,雌性数量由3 增加到9,寿命约延长1. 1 倍(54~115 d )(方差分析中,χ2=41. 192;处理自由度df=3;P<0. 001),但5♀与7♀人工种群寿命差异不显著。
5) 单雌增殖率,人工种群中雌雄数量相等时达到最大,平均每天可增加约2头雌性个体,约为其他处理增殖率的2倍(见表1)。
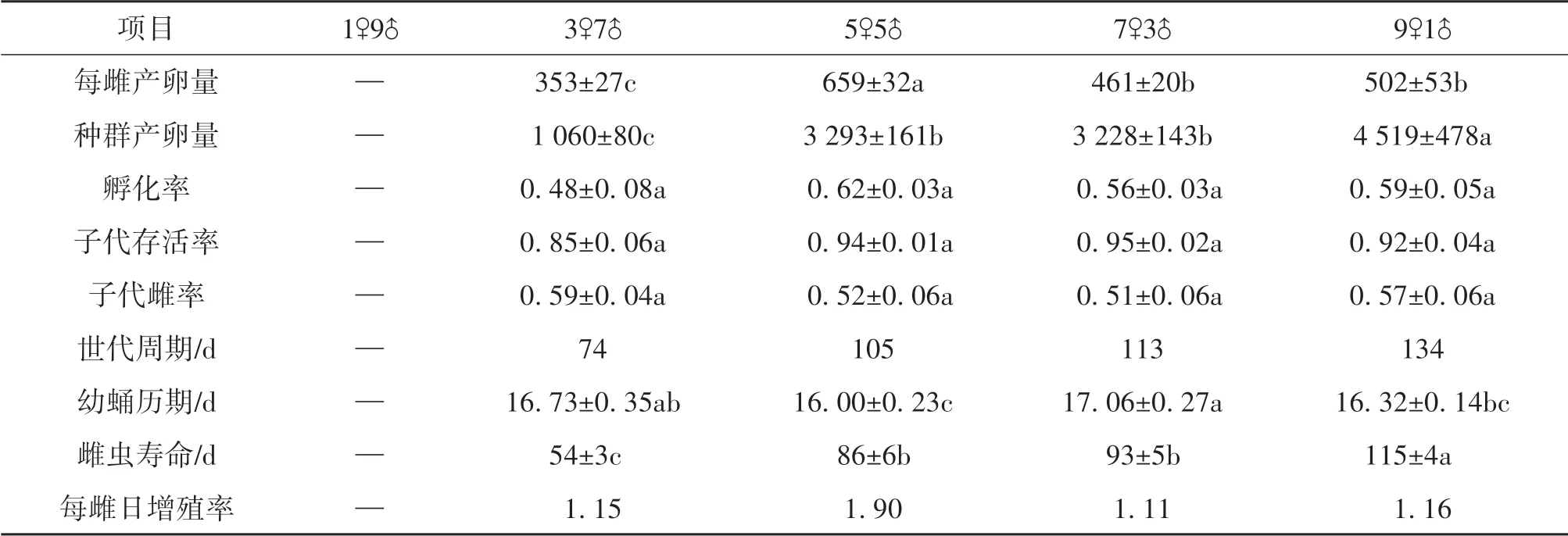
表1 不同性比人工种群生命表参数1)Table 1 Life table parameters for artificial populations with different sex ratio
2.3 不同性比人工种群平均日增加数量和雌虫寿命
1) 5♀5♂人工种群中,日均雌成虫增加数量约为2头,显著高于其他处理(约为2倍)(见图2)。
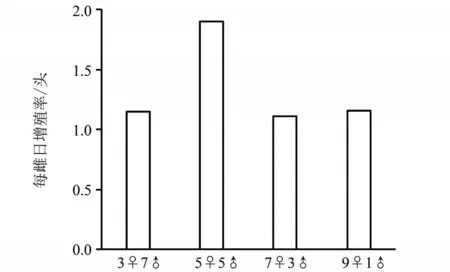
图2 人工种群每雌日均增加雌虫数Fig. 2 Daily increase of female adult individual on average
2) 随雌虫比例增加,雌虫寿命显著延长。其中,9♀种群中,雌虫寿命最长,3 雌的最短。5♀与7♀种群间无显著差异(见图3)。
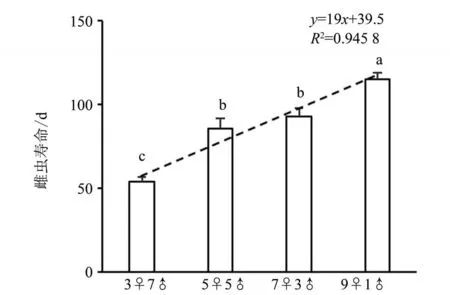
图3 不同性比人工种群中雌虫寿命Fig. 3 Life span of female adult in artificial populations with different sex ratio
3) 无论亲代雌性占比高低,通过一代繁殖获得的子代, 雌性占比都回归到了0. 5 附近(见图4)。
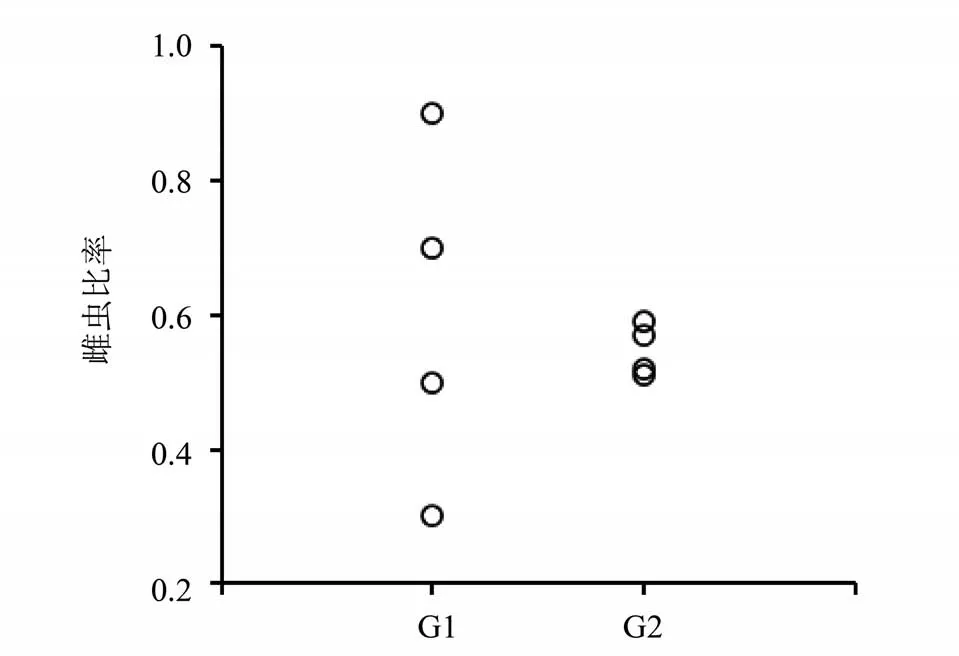
图4 当代与子代成虫雌虫比率Fig. 4 Female adult proportion of parent and offspring generations
3 讨 论
自然界中,异色瓢虫种群性比变化,将引起子代种群结构的变化,而性比的变化可能是食物和光温条件变化导致的。本试验中,虽然实验种群性比设置变化幅度较大,子代性比却维持在0. 5左右, 这可能反映出实验种群遭到的环境压力相对较弱。当雌雄比为1时,异色瓢虫人工种群将处于理想的生活状态。此时,产卵量最大,孵化率和子代存活率较高,雌虫寿命较长。在雌性比低的实验种群中,种群规模一样大,但生活状态变差,如显著缩短的寿命,以及随之降低的每雌增殖率。然而,当雌性比例升高时,虽然雌虫寿命延长,但产卵量明显下降。
成虫性比作为影响种群繁衍的一个重要因素,对产卵量、寿命及子代性比均有显著影响。成虫性比不同,导致了种群内交配等竞争压力,而交配会消耗成虫营养,使得雌虫寿命缩短[21-24]。此外,在对白魔按蚊、烟蚜夜蛾等昆虫的研究中,还发现了合适的成虫性比可提高单雌产卵量[13,15-17],这与本试验结果一致。除了对当代成虫的影响外,成虫的性比对子代也产生了一定影响。报道指出,许多种群会根据当地雄性竞争程度调整后代性比[26],曾有研究认为,雄性个体更偏好雌性后代,以更好地传递它们的基因[27-29]。智利小植绥螨Phytoseiulus persimilis交配过程中精子数量也可影响后代性比,交配时间为15~30 min 时,后代中雄性比例更高,而交配时间增长,雌性后代比例增加[30]。此外,雌性成虫与年龄更大的雄性交配,也将导致类似结果[31]。据研究,水蚤类新孵化的幼虫性别在早期发育中确定下来,内分泌干扰物使得大型蚤初孵幼虫中雄性比例增加[32-37]。不同性比的蠋蝽Arma chinensis(Fallou) 成虫交配繁殖一代后(3♀3♂,2♀4♂,4♀2♂),子代成虫性比间无显著差异,雌性占比约为0. 5[38]。这与本试验结果一致,可能原因是世代间性比没有直接的联系。
异色瓢虫为什么能够入侵各种生态系统并且种群异常繁荣壮大,必定有不同于其他天敌昆虫的特点。我们推测这种特点与其高繁殖力有关,首先,异色瓢虫个体产卵量比较大,是因为其食物范围广,补充营养食物充足(花蜜和各种蚜虫都算补充营养的来源);而孵化率又比较高,可能是因为受精率比较高,雄虫生育力比较强(雄性也会补充营养)。其次,雄虫依靠嗅觉易于及时找到配偶[39-40],交配时雄性精子的长时间传递[41]以及雄虫的配后保护行为[42]都提高了受精成功率。最后,雄性成虫期可以有多个配偶,也增加了繁殖力。
4 结 论
异色瓢虫的最佳性比是1∶1,当异色瓢虫种群受到扰动时,包括对特异性别明显不利或有利的环境变化因素(逆境因子),这个物种在性比上会做出一定的响应,而且总能在逆境因子解除后通过一代繁殖,恢复到最佳性比,这种响应特点不仅在室内种群中得到验证,也在自然种群的调查结果中得到广泛支持。当异色瓢虫种群为最佳性比时,繁殖力最强;而自然种群中性比总是接近最佳值,这可能是异色瓢虫爆发的重要原因之一。
猜你喜欢
中国生育健康杂志(2022年4期)2022-11-25
植物保护(2021年4期)2021-11-12
红蜻蜓(2020年9期)2020-09-26
小学生作文(低年级适用)(2018年4期)2018-11-29
安徽农学通报(2017年9期)2017-05-19
中国医药导报(2017年6期)2017-04-06
农业与技术(2016年22期)2017-03-07
小朋友·快乐手工(2015年10期)2015-11-02
百科知识(2015年7期)2015-04-08
小朋友·快乐手工(2015年2期)2015-03-13
