
雪峰虫草寄主巨疖蝙蛾的生殖行为节律
2021-10-19 13:37李幸,陈珊,周琼
环境昆虫学报 2021年5期
李 幸,陈 珊,周 琼
(湖南师范大学生命科学学院,长沙 410081)
昆虫作为动物界的最大类群,生殖行为对其种群繁衍起着至关重要的作用,生殖行为主要包括异性间的求偶、交配以及产卵等。地球因严格的公转和自转产生了准确的季节交换和昼夜更替,一些非生物因素(例如温、湿度光照等)随之表现出周期性变化,自然生物的行为与生理形成了周期性节律特征,形成生物钟(Pittendrigh, 1993; Takahishi, 1995; Saundersetal., 2002)。豆野螟Marucavitrata(Luetal., 2008)、二化螟Chilosuppressalis(Kanno, 1981)、蜀柏毒蛾ParocneriaorientaChao(张坤胜等, 2012)、玫瑰色卷蛾Choristoneurarosaceana(Delisle, 1995)和榆木蠹蛾Holcocerusvicarius(杨美红等, 2010)等蛾类昆虫的生殖行为均表现出明显的节律性。研究昆虫的生殖行为及其节律,有利于掌握该虫的生物学特性及种群活动规律,为益虫的利用和害虫的防治提供重要理论依据。
巨疖蝙蛾EndoclitadavidiPoujade隶属于鳞翅目Lepidoptera蝙蝠蛾科Hepialoidea,是一种蛀干性昆虫(朱弘复等, 2004),其幼虫被雪峰虫草菌Ophiocordycepsxuefengensis寄生感病后形成名贵中药材雪峰虫草(李纲等, 2017; 陈珊等, 2017)。通过分子生物学和真菌分类学的鉴定,证实雪峰虫草和冬虫夏草的亲缘关系很近(Wenetal., 2013)。目前有关虫草寄主昆虫蝙蝠蛾的生殖行为鲜有报道(胡德具, 2015; 黄雪峰等, 2019)。本研究在前期工作的基础上,系统研究了巨疖蝙蛾的生殖行为及其节律,以期为进一步揭示蝙蝠蛾的生殖行为机制,以及规模化人工饲养巨疖蝙蛾和培育雪峰虫草提供依据。
1 材料与方法
1.1 供试虫源和饲养条件
1.1.1供试虫源:
分别于2015年6月-9月和2016年7月-9月自湖南省邵阳市洞口县(27°5′N/110°30′E,海拔300~600 m)采集巨疖蝙蛾幼虫,带回实验室进行室内饲养。
1.1.2饲养条件:
幼虫用高8.5 cm、直径6 cm的组培瓶里,用人工饲料(周琼等, 2017)单头饲养至化蛹。人工气候箱(RXZ-280D-LED)温度19~22℃,相对湿度80%±2%、无光照。羽化后按性别和日龄(从羽化到第一个光周期结束为一日龄,以此类推)分装,选取展翅正常的健康雌、雄蛾供试。
1.2 研究方法
分别于2015年和2016年的9月至10月在实验室条件下(温度25℃±1℃,相对湿度60%±5%)进行。将羽化当天的成虫按雌、雄蛾各1头或1雌2雄合笼(30 cm×30 cm×30 cm的尼龙网笼,40目)饲养观察,合笼前雌蛾全部为处女蛾。共观察23笼,其中10笼为1雌1雄、13笼为1雌2雄,观察记录其求偶行为。
1.2.1巨疖蝙蛾的求偶和交尾行为的日节律研究方法
野外调查和预实验观察发现,巨疖蝙蛾成虫仅晚上活动,而白天不活动。因此,夜间采用红外摄像机(莱彩HD-A210)记录其行为。通过预实验,确定拍摄时间在每日的19∶00-次日凌晨03∶00。记录每对雌、雄蛾求偶、交尾始末的时间及次数,雌雄蛾腹部末端相接记为交尾。
1.2.2巨疖蝙蛾求偶率和交尾率随日龄的变化规律
与1.2.1相同,记录笼中求偶雄蛾数量,并计算各日龄求偶雄蛾的百分率。雄蛾求偶行为以雄蛾的后足气味刷舒展开为标准。某日龄雄蛾求偶率(%)=(当日龄求偶雄蛾数/当日龄雄蛾总数)×100。
1.2.3巨疖蝙蛾的产卵行为
与1.2.1同时进行。每个养虫笼下的白色瓷盘铺有潮湿滤纸,以收集雌蛾产下的卵。记录卵量,直至雌蛾死亡。
1.2.4产卵的日节律性
同上,每天分4个时间段(8∶00-12∶00、12∶00-18∶00、18∶00-23∶00、23∶00-次日8∶00)收集虫卵。
1.3 数据分析
采用SPSS 19.0统计软件,对所获得的数据进行单因素方差分析和邓肯氏新复极差多重比较,分析各组数据间的差异显著性水平。
2 结果与分析
2.1 巨疖蝙蛾求偶和交尾行为节律
巨疖蝙蛾的求偶行为仅发生在暗期,雄蛾振翅、并展开气味刷,一旦雌、雄蛾腹部末端相接触,即可进行交尾。交尾结束后,雄蛾爬离,雌蛾开始产卵。雌蛾不交尾也可产卵。
2.1.1巨疖蝙蛾的求偶和交尾行为的日节律
晚上室内一关灯进入黑暗环境,巨疖蝙蛾即很快由静息状态进入求偶前期的活动状态,时间间隔≤5 min。求偶和交尾行为主要发生在进入暗期的40 min内(图1),可以看出,求偶行为发生的频次最高是在进入暗期的5~10 min内(包括求偶过程中雌、雄蛾的多次掉落)。交尾行为主要发生在进入暗期的10~25 min。
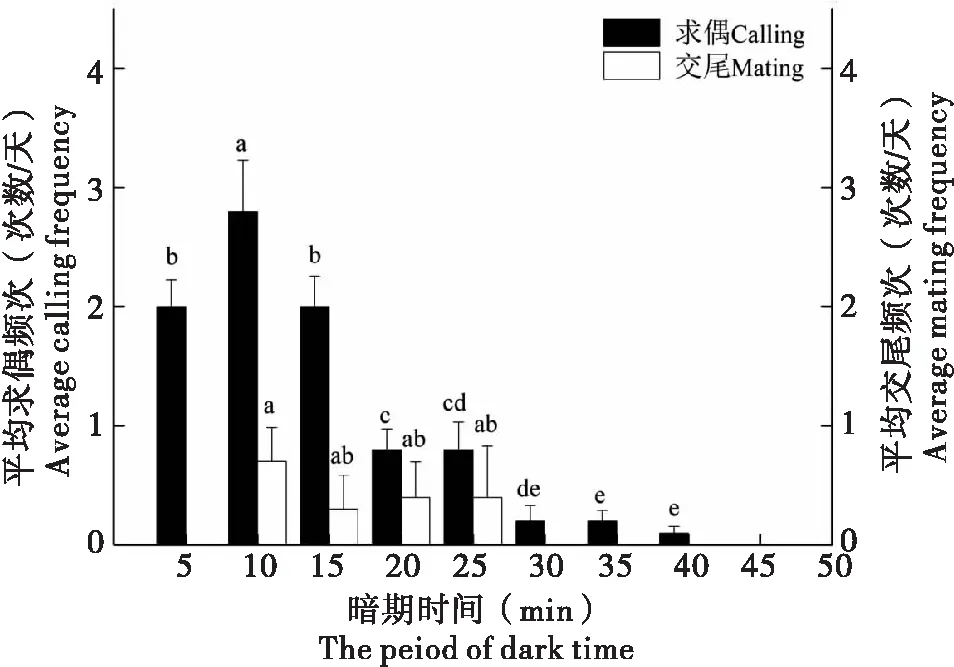
图1 暗期不同时段巨疖蝙蛾的求偶和交尾行为频次
视频的观察分析发现,雌蛾可以多次交尾,但大多数(约70.00%)每晚只交尾一次(图2-A),而且巨疖蝙蛾的交尾时间短暂,一般1 s到71 s不等,平均交尾时长为10.11 s。其中,交尾时长1~5 s的占73.34%(图2-B)。一晚有多次交尾时,交尾间隔时间在1~36 s不等,平均约13.64 s,仅观察到一例是雌雄蛾均掉落重新爬上笼顶再求偶交尾,间隔时长达420 s。
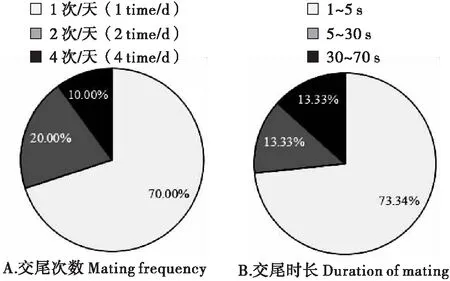
图2 巨疖蝙蛾不同交尾次数和时长所占比例
2.1.2巨疖蝙蛾求偶率和交尾率随日龄的变化规律
求偶行为随日龄的变化规律见图3。可以看出,1日龄雄性巨疖蝙蛾一进入暗期即表现求偶行为,且日求偶率在2~3日龄达到高峰,2日龄求偶率可达87.33%,随后求偶率随日龄的增加逐渐降低,7日龄后的雄蛾极少见求偶行为,在所研究的36头雄蛾中,仅观察到1头在8日龄仍有求偶行为,但并未与雌蛾交尾。
交尾行为随日龄的变化规律:在养虫笼内,巨疖蝙蛾的成虫羽化当日(1日龄)即可交尾,但交尾率较低,仅为25.82%,2日龄雌虫的交尾率达到最大值,为61.09%,此后交尾率会逐渐降低(图3),6日龄之后的雌虫未观察到交尾行为,交尾平均值为39.41%。
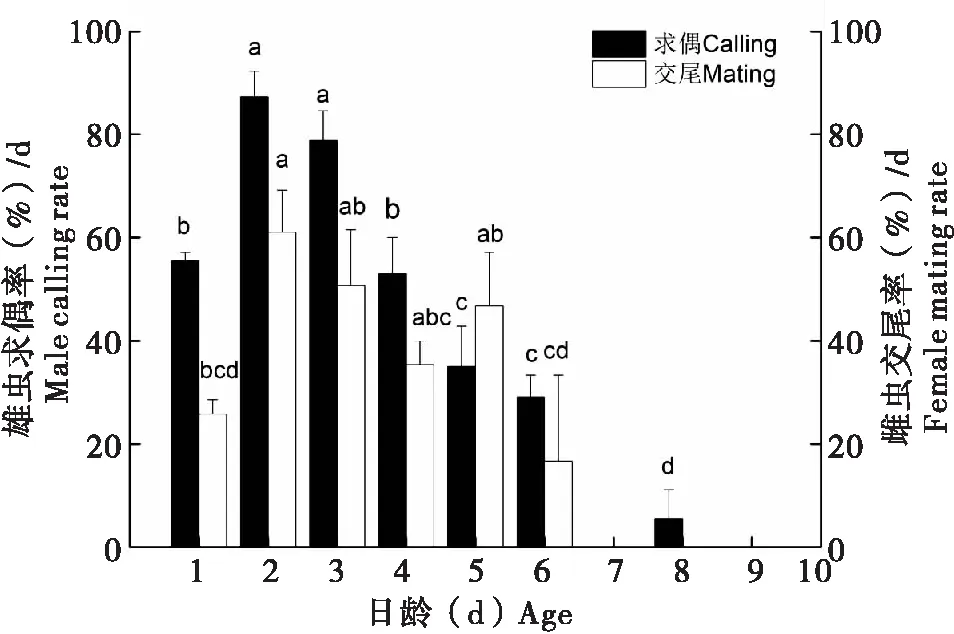
图3 不同日龄巨疖蝙蛾的求偶率和交尾率
2.2 巨疖蝙蛾的产卵行为及其节律
2.2.1巨疖蝙蛾的产卵行为
巨疖蝙蛾雌虫刚羽化时即可排卵。据室内观察,成虫产卵分数次进行,产卵呈喷射状,卵粒散落,无黏着性。产卵时,雌虫攀附在笼内,双翅静止或轻微的振动,腹部末端产卵孔有节奏的收缩和舒张,将卵一粒粒连续喷射出来,产出的卵渐渐聚集成小丘状(图4)。雌蛾在一处产卵较多时,就会攀爬或飞到另一处继续产卵。

图4 正在产卵的雌性巨疖蝙蛾
巨疖蝙蛾的繁殖力强,平均产卵量约为4 250粒/雌,最少约2 500粒/雌,最高可达到约10 000粒/雌。对其中16头雌蛾日产卵量的统计结果见图5,可以看出产卵量在羽化第二天(2日龄)时达到高峰979粒,随后逐日递减。巨疖蝙蛾的产卵行为通常持续到雌虫死亡,大多雌蛾死后腹内仍有少量遗腹卵。
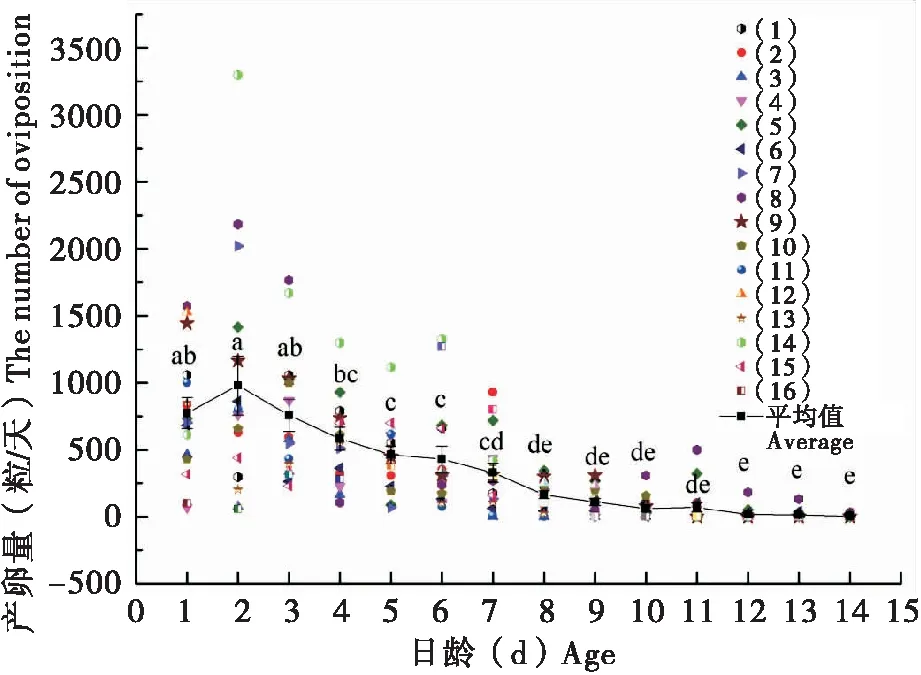
图5 巨疖蝙蛾日产卵量随日龄的变化情况
2.2.2产卵的日节律性
巨疖蝙蛾单次产卵上百粒,一天内可多次产卵,表现出明显的日节律性。每天分4个时间段连续对其产卵量进行统计,发现巨疖蝙蛾产卵时间主要发生在夜间23∶00到翌日上午8∶00,卵量与其他时间段相比差异显著(P<0.05)(图6-A),上午时间和午后偶见产卵;其中19头雌虫共计连续统计到151次产卵,其中有134次发生在22∶00-次日8∶00时,占总产卵频次的88.74%(图6-B)。
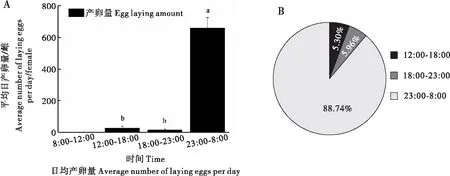
图6 巨疖蝙蛾的产卵行为日节律
3 结论与讨论
昆虫求偶和交配行为的发生主要由内在生理因素决定,但不同种类昆虫长期在不同的环境下,求偶和交配的时间、高峰存在差异,这也是同域物种产生生殖隔离的一个重要机制(Jennions and Petrie, 1997; Gavrilets, 2000)。昆虫的求偶是一种自发行为,表现为两性之间的相互吸引,是昆虫完成交配行为的必要环节,对昆虫繁衍不息起着举足轻重的作用。鳞翅目昆虫的求偶交配都有一定的时辰节律,仅出现在某一特定时段(张坤胜等, 2012),如松异舟蛾Thaumetopoeapityocampa求偶高峰期为暗期的6.4 h(Zhang and Paiva, 1998)、甜菜夜蛾SpodopteraexiguaHübner求偶高峰期为暗期的6.5 h(董双林和杜家纬, 2002)、印度谷螟Plodiainterpunctella高峰期为暗期2 h(Nordlund and Eugene, 1974)、豆野螟求偶高峰期为暗期的5 h(陆鹏飞等, 2007)。本研究发现,自然网室环境下,巨疖蝙蛾通常傍晚开始求偶、交配活动,18∶30-19∶00为活动高峰期;同时,室内研究发现,22∶00-23∶00熄灯后5 min内即可以观察到巨疖蝙蛾开始求偶活动,而有灯光则一直不动,说明黑暗条件可能是触发该虫求偶、交尾行为的重要诱发因素。昆虫性行为的日节律受内源性的昼夜节律调节(Giebultowicz, 2000),随着对昆虫昼夜节律机制的深入研究发现,Cry基因可构建新型的昆虫生物钟机制(Yuanetal., 2007),昼夜节律时钟基因可在神经系统外的其他组织中表达,意味着昼夜节律时钟不是传统上认为的由一个中枢主时钟实现的(Giebultowicz, 1999),昼夜节律是生物体对环境周期的适应,所经历的环境条件对其行为周期性起着至关重要作用(Castrovill and Carde, 1979)。
Yangetal.(2020)研究认为夜行性昆虫在黄昏或夜间活动,视觉受光线限制,远距离上利用嗅觉分辨寄主植物,在短距离上比起颜色主要依靠亮度进行觅食和产卵(Yangetal., 2020)。不同蛾类交配高峰的时间有差异,笋秀夜蛾Apameaapameoides交配高峰发生在暗间4.5~6 h(叶碧欢等, 2014);榆木蠹蛾交尾高峰出现在23∶00左右,光期未见交尾(杨美红等, 2010);豆野螟交尾高峰期在暗期5 h(Luetal., 2008);毛健夜蛾Brithyscrini具有两个交配高峰,分别在晚上19∶00-20∶00和凌晨00∶00-01∶00(涂小云等, 2012),而巨疖蝙蛾的交配行为高峰发生进入暗期的10~25 min内。蛾类昆虫的求偶交配只在暗期进行,与一天中其他时间活跃的生物区分开,能有效避免天敌的危害,增加种群的存活率(薛羿等, 2017)。本研究观察到巨疖蝙蛾的交尾时长平均约10.11 s,这与胡德具(2015)报道的疖蝙蛾PhassusnodusChu交配时间短暂相一致,但与贡嘎蝠蛾Hepialusgonggaensis交尾时长平均42 min(黄天福等, 1992)、蒲氏钩蝠蛾Thitarodespui交尾时长平均150 min(刘昕等, 2017)相距较远。周祖琳(1989)的研究发现麦蛾Sitotrogacerealella交配时长一般150 min,需要达到30 min以上时精包才能进入雌蛾体内;并认为,雌蛾体内的精包数是麦蛾类是否成功授精的标志,也是交配几次的判断依据。本研究主要观察分析了巨疖蝙蛾的生殖行为节律,但在这数十秒的交尾时间里巨疖蝙蛾是否成功授精尚不清楚,相关工作尚有待进一步的研究明确。另外,巨疖蝙蛾求偶过程中,常出现雄虫或雌雄均掉落的现象,以及求偶成功率低(39.41%),很可能是限制巨疖蝙蛾种群繁衍的重要因素之一。
蛾类的求偶和交尾行为受日龄的影响(Delisle, 1995),与其性成熟程度有关。巨疖蝙蛾在羽化当天就有求偶和交尾行为,2日龄的求偶率和交尾率最高。榆木蠹蛾(杨美红等, 2010)、蜀柏毒蛾(张坤胜等, 2012)、杂食卷叶蛾Platynotastultana(Webster and Carde, 1982),羽化当天即可交尾,雌蛾在2日龄时性信息素含量最高,均表现出2日龄雌蛾交尾率最高。交尾成功率的高低依赖于性信息素的释放及对信息素的感受,两者能否协调一致(陆鹏飞等, 2007)。
不同日龄的巨疖蝙蛾产卵量明显不同,羽化当天即可产卵,2日龄产卵量最高。在一天中的不同时段,以夜间11点至次日8点的产卵量最多,表现为多次产卵、以及单次产卵量多等特点,这与疖蝙蛾(胡德具, 2015)的产卵行为极相似。巨疖蝙蛾一般2~3年一代,其中,幼虫期约一年半(未发表数据),长时间的取食生活,使幼虫体内积蓄充足的营养物质,保证了成虫羽化前生殖器官的发育成熟,羽化时即可进行生殖行为,繁殖后代。雌虫产卵量大,平均约为4 250粒/雌,最高可达到约10 000粒/雌,相比于其它成虫期需要饲喂补充营养的蛾类,如榆木蠹蛾雌蛾最高产卵量720粒/雌(杨美红等, 2010),蜀柏毒蛾雌蛾最高产卵量402粒/雌(张坤胜等, 2012),亚洲型舞毒蛾Lymantriadisparasiatica雌蛾最高产卵量357粒/雌(薛羿等, 2017),巨疖蝙蛾繁殖力强大。巨疖蝙蛾羽化第二天产卵量最大,可能是一种自我保护和适应策略,因为其整个成虫期不摄食,在羽化早期体内蓄积能量充足,能尽量多产卵以充分繁殖后代。最后死亡的雌蛾体内通常还有遗腹卵,说明其生命后期可能已无足够的能量供其产出所有的卵。同时,该虫仅在暗期繁殖行为(包括求偶、交尾、产卵)时活动,完成后又进入悬挂的静息状态,这样可以减少能量的消耗,以保证有足够的能量完成其强大繁殖能力的产卵行为。巨疖蝙蛾的卵散产,与雌蛾内生殖系统缺少粘液腺器官(未发表数据)密切相关,这与玉龙蝠蛾HepialusyulongensisLiang(赵万源等, 1989)相似。巨疖蝙蛾的繁殖能力强,整个成虫期少活动、不取食,卵散产和经常更换产卵位置等,既可以减少成虫被天敌发现,又可以有效防止所产的卵被天敌一次性捕食或寄生,提高后代的存活率,这些因素很可能是蝙蝠蛾科作为鳞翅目昆虫最原始的类群,能适应环境生存繁衍至今的重要原因。
巨疖蝙蛾生殖行为发生在暗期,黑暗条件可能是其进入生殖行为的诱因,羽化当天晚上即可进行求偶、交尾和产卵,交尾时间短暂。在第二天其求偶率、交尾率、产卵量达到最高,之后随着日龄的增加而逐渐降低。巨疖蝙蛾产卵量大的特点可为人工规模化饲养巨疖蝙蛾及培育雪峰虫草提供有利条件。
致谢:承蒙2014级硕士生李纲参与部分研究,在此表示衷心的感谢!
猜你喜欢
资源导刊(信息化测绘)(2022年4期)2022-11-24
作物学报(2022年4期)2022-02-10
安徽林业科技(2021年2期)2021-06-15
空间科学学报(2021年1期)2021-05-22
环境昆虫学报(2021年2期)2021-05-14
四川蚕业(2021年1期)2021-02-12
昆虫学报(2020年6期)2020-08-06
昆虫学报(2020年1期)2020-03-03
小溪流(画刊)(2017年6期)2017-07-10
中国学校体育(2014年4期)2014-12-06
