
直灌咸水与施氮磷对滨海重盐碱地柽柳生长及养分分布的影响*
2021-10-19 08:22邱发根杨莉琳朱向梅陈淑伶
中国生态农业学报(中英文) 2021年10期
邱发根,杨莉琳**,刘 敏,朱向梅,陈淑伶
全球约有9.5亿hm2盐碱地,并以每年133万hm2左右的速度持续增加。我国约有1亿hm2盐碱地[1],主要分布在东北、西北、华北、内蒙及东部滨海地区。随着全球气候的持续变暖这些盐碱地面积及盐碱化程度大概率还会加剧。地处华北低平原的环渤海区盐碱荒地广泛分布,该区地下淡水资源严重匮乏,而咸水资源丰富。受季风气候影响,春季普遍大风、干旱,极易导致高矿化度的浅层地下咸水向上运移造成土壤返盐,加重了土壤的盐碱化程度,而春季是植物生长发育的关键时期,绝大多数植物都会遭受春旱、重度盐碱与土壤贫瘠等多重危害。
柽柳(Tamarix chinensisL.)属于泌盐植物,具有根系丰富、发芽能力强、抗旱、耐盐碱、耐瘠薄、抗逆性强、防风固沙、保水保土以及生态适应性强等特点,是改良盐碱地及滨海景观改造的重要树种[1]。柽柳的嫩枝叶是中药材; 柽柳叶富含蛋白质、脂肪和硫,是优良饲料; 柽柳茎作为优质薪炭材,是农村绿色能源的理想树种,且具有越平茬生长越好的生长特性[2],是优质的再生生物资源。针对重盐碱地区淡水缺乏而咸水资源极为丰富的特点[3],许多学者开展了利用咸水结冰灌溉为植物春季萌发提供适宜土壤条件的研究[4-5],及可灌溉咸水的矿化度[6-9]、柽柳耐受阈值[10]、灌溉量与灌溉时间等单纯的咸水灌溉技术等[11]方面的研究探索。而针对重盐碱地栽植柽柳面临的土壤缺氮(N)、贫磷(P)、春旱和因返盐而加重盐害等关键问题的综合解决途径尚少见报道。
为此,本研究开展了春季灌溉地下咸水与不同施肥的定位试验,研究滨海重盐碱地区地下咸水资源直接灌溉利用能否缓解柽柳春旱与盐害,咸水灌溉对柽柳生长、养分吸收、盐分平衡的影响以及施用氮磷肥与咸水的交互作用。旨在为滨海重盐碱地种植柽柳克服土壤贫瘠、春旱和盐碱等逆境障碍,探索咸水就地利用,实现障碍性资源高效循环、发展绿色能源植物,实现生态恢复与重建提供科学依据。
1 材料与方法
1.1 试验区概况
试验区设在河北省海兴县小山乡,地处河北省东南部渤海低平原区,地势低洼平坦,属暖温带半湿润半干旱季风气候区,年均气温12.1 ℃,1月份均温-4.5 ℃,最低气温-19.9 ℃,霜期和终霜期多分别出现在10月下旬和4月中旬;年平均降水量为582.3 mm,6-8月份降水量约占全年降水量的74%,冬春季降水极少。土壤为滨海砂壤质盐碱土,0~100 cm土层含盐量周年不低于5 g.kg-1。地下水位埋深1.5~2.0 m,盐分为 8.02~9.34 g.L-1。
1.2 试验设计
定位试验始于2008年,为期4年。供试柽柳品种为人工栽植的3年生‘中华柽柳’,试验开始当年平茬,平茬高度为保留地上基茎3 cm以内,以后每隔1年平茬1次。试验为水、肥2因素定位试验,其中,水处理设不灌水与灌咸水处理(W); 肥料设不施肥(CK)、单施氮肥(N)和施氮磷肥(NP)处理,组成不灌水条件下的CK、N、NP和灌溉咸水条件下的WCK、WN和WNP共6个处理。灌咸水与施肥时间为春季最干旱时段的3月底至4月上旬,直接抽提当地地下咸水漫灌,灌水量为750 m3.hm-2。施肥量为N 75 kg.hm-2、P2O537.5 kg.hm-2,肥料品种为尿素和二铵,施肥方法采用穴施,即围绕柽柳距树干15 cm地表处挖10~15 cm深的施肥穴4~5个,施肥后立即覆土; 灌咸水处理在施肥后进行。试验第4年(2011年)暂停咸水灌溉,但施肥处理继续进行。
根据灌水与否作裂区设计,每个区组内不同施肥处理顺序排列,重复3次。区组间隔0.9 m,小区之间间隔0.5 m,小区南北向长10 m,东西走向宽9 m,小区面积90 m2,柽柳株行距为40 cm×70 cm。
1.3 调查测定项目与方法
调查时间:于试验第1年(2008年)的7月3日和9月15日分2次调查植株生长性状并采集植株样品用于测定鲜重、干重和植株含水量; 于第4年(2011年)的4月30日、5月30日、7月9日、8月21日和11月7日定株调查柽柳的生长动态,并于8月21日采集植株样品,用于养分和盐分的测定。
调查方法:每个小区选取代表性柽柳植株6株挂牌,定点测定挂牌植株的株高、地茎、分枝、冠幅等生长指标。
样品采集与处理:每个处理小区采集5株长势均匀的地上部植株,剪短,将茎与叶分开,于105 ℃杀青3 h,75 ℃烘至恒量; 植株样品粉碎后过60目筛,用于测定养分和盐分含量,所有指标的测定方法参见《土壤农化分析》[12]。
1.4 数据分析
采用SAS 9.1软件对数据进行方差分析,并用最小显著性差异法(LSD)进行多重比较。
2 结果与分析
2.1 灌溉咸水与施肥处理对柽柳植株生物学性状的影响
试验当年(2008年)7月初调查结果表明(表1),灌咸水极显著地影响柽柳植株含水率,比不灌咸水的处理平均减少13.3个百分点(P<0.001),但对春夏季柽柳的株高、茎粗、分蘖数和鲜重等没有显著影响(表1); 施肥显著促进柽柳的生长,与对照CK相比,N与NP处理的株高平均增长6.6%与17.2%(P<0.01),鲜重增加47.3%和82.5%(P<0.05),干重提高39.7%与69.6%(P<0.01),含水率提高11.6与10.5个百分点(P<0.01)。咸水灌溉下,与对照WCK相比,WN与WNP处理的株高分别增长13.4%与18.4%(P<0.05),基茎增粗25.7%与27.0%(P<0.05),蘖枝数变化不显著,鲜重增加40.7%与75.2%(P<0.05),干重分别提高43.0%与68.0%(P<0.01)。灌咸水与施肥的交互作用对柽柳植株干重(P<0.01)和含水率(P<0.001)有极显著影响。

表1 第1年咸水灌溉与施肥处理对柽柳植株生物学性状的影响(单株)Table 1 Effect of different treatments of fertilization and saline irrigation on biological characteristics of Tamarix chinensis(per plant)in the first year
试验当年(2008年)秋季(9月中旬)调查结果表明,与不灌咸水相比,灌咸水显著影响柽柳植株鲜重、茎与叶的干重及植株含水率,植株平均鲜重与叶干重分别下降9.5%和12.3%(P<0.05),而茎干重平均增加2.1%(P<0.01),植株含水率下降13.1个百分点(P<0.01)。与对照CK相比,N与NP处理的株高分别增加11.8%与20.4%(P<0.01),茎增粗29.2%与32.6%(P<0.01),鲜重增加69.6%与1.09倍(P<0.05),蘖枝数增加41.8%与67.4%(P<0.05),柽柳叶干重增加56.9%与93.9%(P<0.05),茎干重增加82.1%与1.16倍(P<0.05),植株干重分别提高71.5%与1.07倍(P<0.01); 灌咸水条件下,与对照WCK相比,WN与WNP处理的株高分别增加14.0%与21.7%(P<0.01),茎增粗27.9%与36.0%(P<0.01),蘖枝数增加37.5%与47.9%(P<0.05),柽柳鲜重增加75.3%与1.11倍(P<0.05),柽柳叶干重增加61.7%与1.00倍(P<0.05),茎干重增加73.3%与1.06倍(P<0.05),植株干重分别提高68.8%与1.04倍(P<0.01)。
2.2 不同水肥处理对柽柳植株生长动态的影响
对柽柳的生长动态调查结果显示(图1),NP处理能显著促进柽柳株高、茎粗和冠幅,比单施氮肥(N)和CK处理株高平均增加19.6 cm和29.4 cm,茎粗增加0.9 mm和2.7 mm,冠幅增加10.5 cm和17.4 cm;比灌溉咸水处理(WNP)的株高、茎粗和冠幅平均分别增加18.9 cm、2 mm和14.2 cm。4-11月的生育期内灌溉咸水组各施肥处理的柽柳株高、茎粗和冠幅比不灌咸水组相对应处理平均下降8.0 cm、1.2 mm和4.86 cm,特别是只灌咸水不施肥处理(WCK)生长状况最差; 但是,灌溉咸水条件的WN和WNP处理的株高、茎粗和冠幅均比WCK显著提高,尤其是WNP处理对柽柳分蘖有极显著的促进作用,整个生育期的蘖枝数均显著高于其他处理(P<0.05),比不灌咸水的NP处理增加27.1%,比CK和WCK分别增加24.6%和38.2%,比N和WN分别增加38.4%和20.9% 。
2.3 不同水肥处理的柽柳植株生长速率
柽柳株高在春季-初冬生育期内的生长速率的变化呈坡状下降趋势(图2A)。以春季生长最快,之后总体呈急剧下降趋势,7-8月间,柽柳株高增长几乎停止,8月下旬,株高的生长率呈缓慢回升态势,灌溉咸水能显著减缓柽柳株高生长速率的下降。不灌咸水的NP处理在春季的生长率下降最快,但株高生长速率下降的拐点比其他处理提前1个月,之后株高平稳生长,至11月初仍能保持1.84 mm.d–1生长速率; 其他处理的株高生长率持续下降,在7月中旬出现拐点值,同期NP处理的生长速率(2.58 mm.d–1)是WNP处理拐点值(0.98 mm.d–1)的2.65倍,是不施肥(CK、WCK)和单施氮处理(N、WN)的5~20倍。
柽柳茎的增粗主要在春-夏末(4-8月),这一时段茎粗保持相对较高的增速(图2B),8月入秋之后柽柳增粗的速率急剧下降,8月下旬以后增粗停滞。不同处理对柽柳茎粗增长速率影响差异较大,不灌咸水N处理茎增粗速率最高点发生在春季,之后呈急剧下降; NP和WCK处理的茎在5-6月的增粗速率呈下降趋势,但在暑期(7-8月)增粗速率明显回升; WN处理在春季增粗速率低于其他处理,但在春-夏季迅速增粗,夏季来临前茎粗增速达最大值,之后进入下降趋势; WNP处理在春-初冬季的生育期内呈下降趋势,但4-6月间茎增粗速率显著高于不灌水的NP处理,暑期(7-8月)反被NP处理超越,8月入秋以后增粗速率急剧下降。
冠幅增加速率与株高相近,冠幅增大速率与株高生长速率几乎同步呈坡状下降趋势(图2C)。WNP处理在早春冠幅增加速率最高,但随后冠幅的增加率迅速下降; 而NP处理的冠幅增长速率在春季最低,与WCK的生长速率相同,但在春-夏季有短暂的增加趋势,7-8月暑期冠幅不再增大,8月下旬以后随株高生长而缓慢增长。
柽柳蘖枝的萌发速率呈先升后降的趋势,春季分蘖速率并不高,但春-夏季(5-6月)分蘖迅速升高,至6月中旬分蘖速率达高峰,之后迅速下降,进入暑期(7-8月)分蘖停止。4-5月间CK和WCK处理的分蘖速率最高。初夏季(6月)灌溉咸水组各处理的蘖枝萌发率比不灌咸水平均提高3.26倍,此时WNP处理蘖枝萌发速率是NP处理的5.6倍(图2D)。
2.4 不同水肥处理对柽柳茎杆和叶片养分的影响
灌咸水与施肥处理均对柽柳茎杆的N含量有极显著影响(表2)。灌咸水柽柳茎杆的N含量比不灌咸水处理平均下降13.2%(P<0.01); 不灌咸水情况下N或NP处理分别比CK提高茎杆N含量44.0%和22.0%(P<0.01); 灌溉咸水条件下的WN和WNP处理分别比WCK处理可提高柽柳茎杆N含量14.3%和12.2%(P<0.01); WCK、WN和WNP处理比不灌咸水的CK、N与NP处理的茎杆氮含量分别下降2.0%、23.6%和11.5%(P<0.05)。灌咸水对柽柳茎杆的P、K和C含量没有显著影响,但是灌溉咸水同时施肥则对茎杆有机碳含量产生一定影响(P<0.1)。
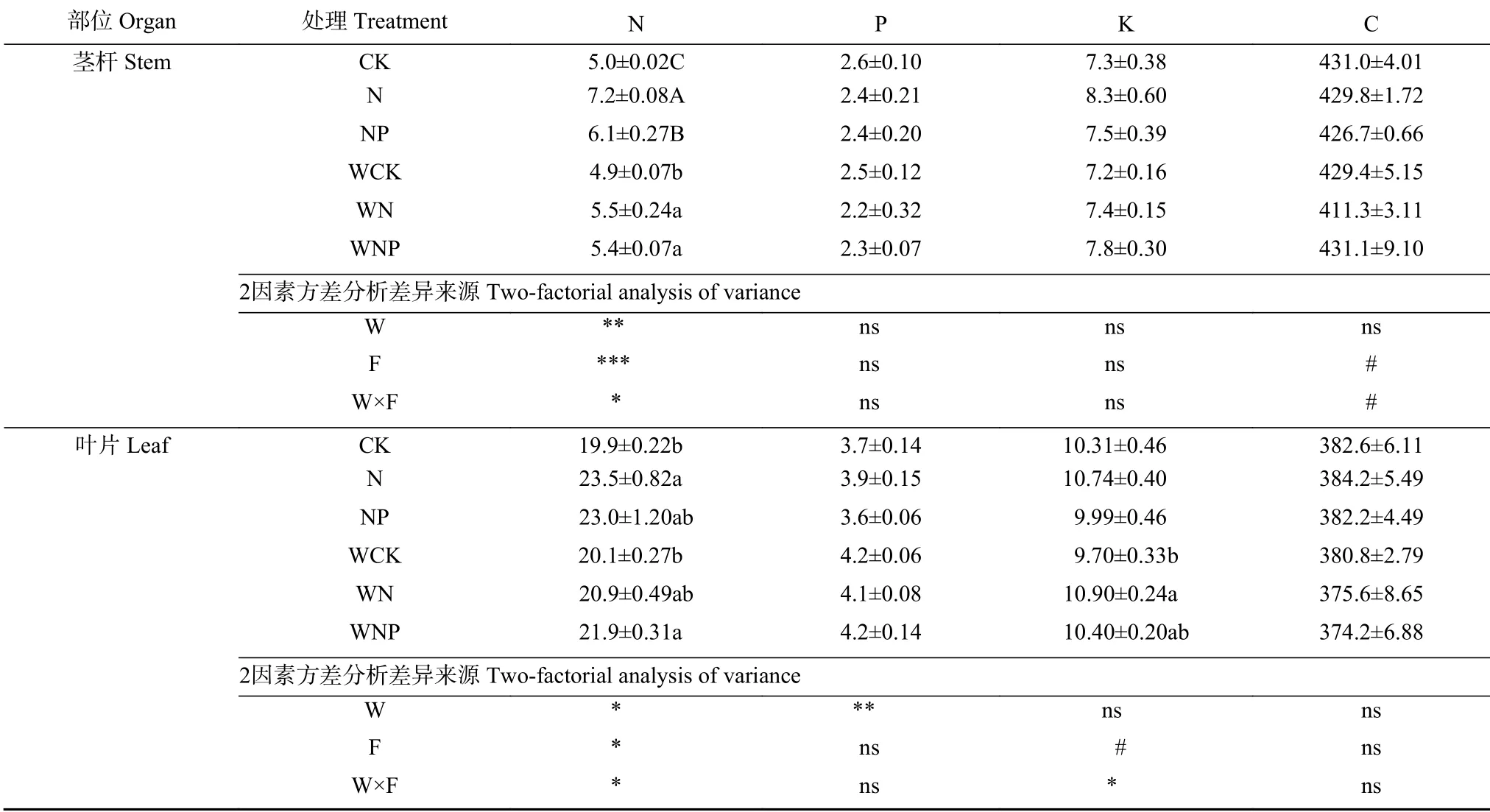
表2 咸水灌溉与施肥处理对柽柳茎叶养分和碳含量的影响Table 2 Nutrients and C contents of stem and leaf of Tamarix chinensis under different treatments fertilization and irrigation of saline water g·kg-1
灌咸水比不灌咸水处理的柽柳叶片N含量平均下降5.3%,WN与WNP处理比N与NP处理的叶片N含量分别下降11.1%和8.7%(P<0.05); 而叶片P含量平均提高11.8%(P<0.01)。施肥显著提高柽柳叶片的N、K含量,N和NP处理比CK的叶片N含量提高18.1%和15.7%; WN和WNP处理分别比WCK处理的N含量提高4.4%和9.1%,同时K含量提高12.7%和8.0%。与不灌咸水相比,灌溉咸水与施肥的交互作用对叶片N、K含量影响显著(P<0.05),但对叶片P和C含量影响不显著。不灌溉咸水时柽柳叶片的N、P、K含量分别是茎杆的3.62倍、1.51倍和1.34倍,灌溉咸水后分别上升至3.92倍、1.79倍和1.38倍; 而C含量变化不明显。
2.5 不同水肥处理对柽柳茎杆和叶片盐分离子的影响
灌咸水与施肥对柽柳茎杆Na+没有显著影响,但灌溉咸水的柽柳茎杆Ca2+与Mg2+含量分别比不灌咸水平均降低12.8%(P<0.05)与21.0%(P<0.01); 相同施肥处理下,WN比N的茎杆Mg2+下降22.4%(P<0.01),WNP比NP处理的茎杆Mg2+下降20.4%(P<0.05); WN和WNP处理比WCK处理的茎杆Ca2+分别提高32.5%和26.7%(P<0.05),Mg2+分别提高22.4%和11.2%(P<0.05); 不灌咸水条件下施N与NP处理比对照CK茎杆Mg2+含量分别提高25.2%与11.3%(P<0.05)。灌溉咸水和施肥打破了柽柳茎杆的盐分离子的平衡(表3)。

表3 咸水灌溉与施肥处理对柽柳茎杆盐基离子含量及离子比的影响Table 3 Effect of fertilization and irrigation of saline water on stem ions contents and ions ratios of Tamarix chinensis
从表3中的离子比也可以看出,灌溉咸水柽柳叶片Na/Ca 和K/Ca比下降,而Ca/Mg上升,其中,灌溉咸水的WNP处理比WCK的柽柳叶片K/Na、K/Ca和Ca/Mg均有下降的趋势。不灌咸水条件的N 和NP处理比CK的茎秆K/Na、Na/Ca 和K/Ca比上升,而Ca/Mg下降; 相反,灌溉咸水后的WN与WNP比WCK处理的茎秆K/Na、Na/Ca 和K/Ca比均下降,而Ca/Mg上升的趋势。
从表3可知,灌溉咸水对叶片Na+含量没显著影响,但施肥均显著提高了柽柳叶片的Na+含量(P<0.01),N与NP处理分别比CK处理的柽柳叶片Na+含量提高26.2%与32.8%(P<0.01),WN和WNP处理分别比WCK处理的叶片Na+含量提高22.7%与20.5%(P<0.01)。灌溉咸水比不灌溉咸水的柽柳叶片Ca2+含量平均提高18.8%(P<0.001),不灌溉咸水条件下,N和NP处理分别比CK处理的柽柳叶片Ca2+含量提高5.88%和15.6%(P<0.05),灌溉咸水条件下WN和WNP处理比WCK处理的叶片Ca2+分别提高7.5%和14.8%(P<0.05)。施肥处理相同条件下,WN与WNP处理分别比N与NP处理的叶片Ca2+提高20.3%和17.6%(P<0.01)。灌溉咸水比不灌咸水的柽柳叶片Mg2+平均提高9.3%(P<0.01),灌溉咸水条件下的WN与WNP处理比对照WCK的柽柳叶片的Mg2+含量分别提高17.9%和22.7%(P<0.05),不灌咸水条件下施肥对Mg2+含量影响不显著(表3)。施肥处理相同条件下,WN与WNP处理分别比N与NP处理的叶片Mg2+含量提高14.7%和13.0%(P<0.01)。
灌溉咸水和施肥后柽柳茎杆与叶的盐分比例发生改变。不灌溉咸水时柽柳叶片的Na+、Ca2+、Mg2+含量分别是茎杆的2.92倍、8.88倍和3.94倍,灌溉咸水后分别为2.88倍、11.86倍和5.41倍。可见,灌溉咸水后叶片中的Ca2+、Mg2+明显提高,Na+略有下降(表3)。
3 讨论
3.1 灌溉咸水与施NP对柽柳生物学性状的影响
与之前的报道一致[13],本研究也发现灌溉咸水引起柽柳植株鲜重与叶干重的显著下降,但柽柳的茎干重显著增加,表明叶片比茎秆受咸水的影响大; 灌溉咸水的柽柳植株含水率在7月与9月平均分别下降13.3个和13.1个百分点。植株受盐胁迫失水,与蒸腾作用及光合作用密切相关,因为植株叶片通过气孔进行CO2/H2O气体交换,实现植株体内的H2O与环境CO2的相互作用; 盐胁迫条件下植株完成光合作用获得最终产物的过程是以失去更多水分为代价,即通过加大蒸腾来促使有机碳积累和植株生长[14]。本研究结果中灌溉咸水的柽柳茎杆与叶片C含量没有明显变化,植株干重不减反增,也证实了为保证固碳量和茎干重不减,柽柳是以加大植株蒸腾失水为代价的,植株失水首先受影响的就是叶片,这也就解释了灌溉咸水柽柳叶片干重降低的原因。
传统理论认为,N素对植物营养贡献最大,因为N与植株C固持的相关性最大[15]。滨海重盐碱地人工柽柳灌溉咸水条件下的现有施肥技术也仅施氮肥[16]。然而,本研究中灌溉咸水条件下的WNP处理的柽柳株高、茎粗、分蘖、植株干重及含水率均显著高于WN处理(表1),本研究结果与Warren等[17]报道一致,在佐证P对树木植株不可或缺的作用同时,并对滨海重盐碱地人工柽柳现有的单施氮肥的技术提出质疑。
3.2 灌溉咸水与施肥处理对柽柳生长发育动态的影响
本研究发现,春季是柽柳生长的最佳时节,植株长高、茎增粗、冠幅增大和分蘖主要发生在春-夏季,但并不是同步进行(图2)。柽柳的生长与蘖枝萌发速率不同步归因于各组织器官发育成长需要独特的温度所驱动[18]。而灌咸水与NP处理直接影响柽柳生长与分蘖的速率。相同施肥处理条件下灌溉咸水处理的柽柳长势(株高、茎粗)明显低于不灌咸水处理(图1),表明连续灌溉咸水加剧了柽柳的盐胁迫,这可以从土壤盐分含量上升得到解释(数据待发表)。但施NP能显著缓解盐胁迫,表现在不灌水条件下的NP处理的柽柳长势(株高、茎粗、冠幅)明显优于其他处理(图1),且灌溉咸水条件下的WNP可延缓株高和茎粗在春-夏时段内(5-6月)生长速率下降,但能加速这一时段蘖枝萌发(图2)。表明在连续灌溉咸水盐胁迫加剧条件下,WNP处理以加速萌发蘖枝补偿长高和增粗受到的抑制,或者说受连续灌溉咸水胁迫时,柽柳利用WNP的条件蘖枝优先萌发,以使后代能适应新的水盐环境。华北地区春季气候回升,极易返盐,春季灌溉地下咸水可稀释土壤表层高浓度的盐分,有效缓解春旱; 其次,灌溉咸水土壤湿度增加利于养分吸收,特别是P营养的吸收,这与灌溉咸水处理提高柽柳叶片P含量相互印证(表2)。反过来,土壤N、P营养及咸水中必需盐基离子对植株的及时供给,使得柽柳生长速率的下降可得到有效缓解。此前也有研究报道,在盐碱地灌溉微咸水后柽柳的生长、光合能力、蒸腾耗水量和水分利用效率均比淡水效果显著[19]。
3.3 灌溉咸水与施肥处理对植株茎杆和叶片养分、有机碳与盐分含量的影响
灌溉咸水与施肥处理均对柽柳茎杆的N含量有极显著影响(表1)。灌溉咸水处理的柽柳茎杆和叶片的N含量显著下降。尽管施N或NP可提高茎杆与叶片的N含量,但相同施肥量下咸水灌溉处理的茎杆和叶片N含量显著下降,但WNP处理的下降幅度低于WN处理。可见,灌溉咸水下的施NP可缓解咸水对N吸收的抑制,即灌溉咸水条件下更需要增施磷肥以增加柽柳对N的吸收。这是因为 N的作用与土壤P含量密切相关[19],土壤P不足会限制N的吸收及光合作用的功能发挥[19-21],反过来,增施磷肥则利于N的吸收和光合作用。
灌溉咸水对柽柳茎杆的P、K和C含量没有显著影响,但柽柳叶片P含量显著提高。由于土壤P含量直接关系叶片P和叶片N的含量[19,22],那么灌溉咸水是土壤难溶性P得以活化还是促进了P的吸收并向叶片转移值得进一步研究。
Zhu[23]认为,盐胁迫条件下植株大量吸收Na+,并取代K+,导致K+下降,Na/K比提高。本研究中从施肥后叶片Na+与K+含量的提升比例可以看出,柽柳叶片Na+含量提升高于K+,但在茎杆中不明显。K+既是重要的营养元素,同时也具有渗透调节作用,不是所有部位都因土壤溶液中Na+多而大量吸收或以Na+代K+,这与Wright等[24]的研究结果一致。
与不灌咸水相比,灌咸水后柽柳茎杆Ca2+与Mg2+含量显著降低,但叶片Ca2+与Mg2+含量提高(表3)。表明WNP处理促进Ca2+和Mg2+从茎秆向叶片转移,打破了这2种离子在柽柳茎杆与叶片分布的平衡。这印证了灌溉咸水引起柽柳植株失水是因为蒸腾量加大的报道[14],水分蒸腾过程中同时装载更多Ca2+和Mg2+从茎秆向叶片运转。表明柽柳植株对盐基离子Ca2+、Mg2+和Na+运输具有选择性[25-26]。结合灌溉咸水叶片P含量的显著提升,可以推侧可能与Ca2+和Mg2+同时借蒸腾作用被运输到叶片,但这些离子是否协同吸收、运转还需通过进一步研究证实。
4 结论
首次灌溉咸水极显著地降低柽柳植株含水率、植株鲜重与叶干重,但不影响茎干重。连续灌溉咸水抑制柽柳生长,但蘖枝萌发率显著提高,茎杆与叶片的N含量显著下降,但柽柳叶P含量平均提高11.8%。灌溉咸水条件下施氮磷肥(WNP)处理可减缓春-夏季(5-6月)柽柳长高与增粗速率的下降,加速蘖枝萌发,提高叶片N、K含量,促进Ca2+和Mg2+从茎秆向叶片转移,改变Ca2+和Mg2+离子在茎叶的分布。连续灌咸水抑制柽柳生长的负面作用不可忽视,但灌溉咸水条件下WNP处理可通过加速分蘖、促进养分吸收和盐基离子运输来缓解其负面影响。
致谢:感谢中国科学院遗传与发育生物学研究所农业资源研究中心张秀梅高级工程师协助调查采集部分植株数据!
猜你喜欢
安徽农业科学(2022年19期)2022-10-29
绿色科技(2022年11期)2022-07-02
绿色天府(2022年2期)2022-03-16
云南农业(2021年10期)2021-10-22
云南农业(2021年9期)2021-09-24
云南农业(2021年8期)2021-09-06
云南农业(2021年3期)2021-04-24
山东工业技术(2018年13期)2018-08-20
安徽农业科学(2017年3期)2017-08-13
红领巾·成长(2016年1期)2016-09-10
