
非编码RNAs在肉牛肌肉发育和脂肪沉积中的研究进展
2021-10-18 09:39沈维军万发春
动物营养学报 2021年9期
曾 斌 高 骞 沈维军 万发春
(湖南农业大学动物科学技术学院,长沙 410128)
牛肉肉质鲜美、富含必需氨基酸和微量元素,是我国居民的主要肉类来源之一。根据《2020年度肉牛牦牛产业技术发展报告》,我国在2020年牛肉产量为678万t,居世界第3位。我国也是全球牛肉进口最多的国家,全年牛肉进口量为275万t,存在巨大的牛肉缺口,优质牛肉更是供不应求。牛肉高产优质是当前肉牛营养调控和分子育种研究的重点和热点,其主要涉及肉牛的肌肉发育和脂肪在不同部位的沉积。随着全转录组技术的成熟和广泛应用,非编码RNAs(non-coding RNAs,ncRNAs)相关研究在畜牧生产领域得到迅速发展,越来越多的ncRNAs被鉴定为动物肌肉发育和脂肪形成的关键调控因子。肉牛增量提质需要尽可能增加瘦肉(肌肉)组织以及肌内脂肪(大理石花纹)含量,同时减少价值较低的腹腔脂肪和皮下脂肪含量。因此,这篇综述就ncRNAs对肉牛肌肉发育和脂肪形成的调节作用进行了归纳总结,以期为肉牛高效养殖提供理论参考。
1 肉牛肌肉发育和脂肪沉积概述
肌肉发育和脂肪沉积对肉牛的生产和提质都至关重要。肉牛肌肉细胞、脂肪细胞在胚胎发育过程中都来自一个共同的祖细胞库[1]。骨骼肌作为肉牛躯体最主要的组成部分,与产肉力等经济性状直接相关。骨骼肌的发育可大致分为胚胎期、胎儿期和出生后期[2]。在胚胎期和胎儿期,骨骼肌发育主要包括肌肉纤维细胞的形成和肌肉纤维的进一步增加,即肌细胞的形成和增殖[3]。肉牛肌纤维细胞是在出生前形成的,出生后不再有净增加。出生后肌肉的生长主要通过肌卫星细胞增殖和分化使肌纤维体积增大[4],即肌管和肌纤维的形成,这也表明肌卫星细胞在动物出生后肌肉生长中发挥关键作用。
肉牛的脂肪沉积是一个复杂的生理生化过程,主要依赖于脂肪前体细胞的增殖、分化和成熟。哺乳动物脂肪细胞主要在胎儿和出生后早期形成[3,5],脂肪细胞的总数是在成年期前就达到稳定状态[6]。成年后则主要表现为体积的增加和脂滴的聚集(分化和成熟)[7]。随着动物年龄的增长,祖细胞逐渐减少是产生新脂肪细胞能力下降的主要原因[8]。肉牛脂肪主要存在于皮下、腹腔内脏周边和肌内。其中肌内脂肪的沉积和分布(即大理石花纹等级)是评价牛肉品质的主要依据。动物不同组织部位的脂肪沉积呈现出明显的时序性和组织特异性,肌内脂肪的沉积次序要滞后于腹腔脂肪和皮下脂肪,沉积难度更大[9]。因此,控制脂肪更多的在肌内沉积而不是在其他部位组织沉积一直是肉牛脂肪沉积研究的热点和难点。有证据表明,肌内脂肪细胞与皮下、内脏脂肪细胞存在差异[10-11],在胎儿肌肉发育过程中,肌内脂肪细胞与肌源性细胞具有更加接近的祖细胞。肌内脂肪细胞这种独特的发育起源特性提供了在不提高动物整体肥胖的情况下特异性增强牛肉大理石花纹的靶点[1]。
2 ncRNAs及其作用机制
ncRNAs作为一类表观遗传学调节因子,在复杂的生命活动中参与了众多基因的表达调控。目前,在关于动物生长、发育、免疫等生理功能的ncRNAs研究中,微小RNA(microRNA,miRNA)、长链非编码RNA(long non-codingRNA,lncRNA)和环状RNA(circular RNA,circRNA)这3种ncRNAs的研究最为多见。miRNAs是长度为18~25个核苷酸的ncRNA,其作用方式通常是通过与靶基因的3′非编码区(3′-UTR)结合,抑制蛋白翻译或降解mRNA来调节基因的表达。因为miRNA在物种间保守性强、命名规范、研究方法成熟和数据库完整,是研究最为深入的一种ncRNA。lncRNA的长度大于200个核苷酸,几乎没有蛋白质编码能力[12]。根据现有的研究,lncRNA的作用机制分为以下类型:1)作为miRNA海绵;2)转录和转录后干扰;3)诱导染色质重塑和组蛋白修饰;4)结合特定的蛋白质,调节其活性或改变其定位[13]。其中,lncRNA作为miRNA的海绵参与竞争性内源性RNA(competing endogenous RNA, ceRNA),主要是通过降低miRNA活性,导致miRNA的靶基因表达升高,这也是目前lncRNA最为详细的作用机制研究。circRNA是一种具有共价闭环结构的ncRNA,已成为RNA领域的研究热点。与线性的RNA相比,circRNA不具备5′末端帽子结构和3′末端poly(A)尾巴结构[14]。目前的研究表明,circRNA产生机制主要包括直接转录组反向剪切、外显子跳跃、内含子的反义互补序列配对模式以及依赖于RNA结合蛋白的环化模式等[15-16]。目前研究发现,circRNA的作用机制主要包括直接调节亲本基因的转录和翻译、竞争性结合miRNA分子,也有研究表明circRNA还具有编码小肽和蛋白质的能力[17]。在生物学功能上,与lncRNA类似,circRNA作为miRNA海绵分子,参与ceRNA网络调控的报道最为多见。目前,ncRNAs研究的主要方法是首先通过高通量测序或芯片技术找出差异表达、高表达的ncRNA,PCR或者实时荧光定量PCR(qRT-PCR)验证ncRNA表达水平;然后通过生物信息学分析预测靶基因和参与调控的信号通路;最后通过对ncRNA敲减、过表达研究ncRNA的功能,采用RNA pull-down、免疫共沉淀等技术确定ncRNA与蛋白质的互作,采用双荧光素酶报告基因验证miRNA和mRNA/lncRNA/circRNA的互作情况。
3 ncRNAs与肉牛肌肉发育
3.1 miRNA与肉牛肌肉发育
在肉牛肌肉发育的研究中,miRNAs已被广泛报道为骨骼肌性状的重要调控因子。高通量测序技术作为快速筛选差异miRNA的方法,在肉牛肌纤维细胞的增殖和分化的研究中广泛应用。Zhang等[18]和Wang等[19]对牛骨骼肌卫星细胞分化前后的miRNA进行测序分析发现了多个差异的miRNA,功能富集表明这些差异的miRNA主要在细胞代谢、肌动蛋白细胞骨架调控和丝裂原活化蛋白激酶(MAPK)信号通路上富集。Sun等[20]通过对秦川牛胎儿期和成年期的背最长肌的miRNA进行高通量测序分析,共鉴定出521个miRNA。经过qRT-PCR检测到25个高表达的miRNAs,其中miR-206、miR-1、miR-133等5个miRNA在肌肉组织特异性高表达miRNA,提示这些miRNA可能在牛肌肉组织的发育中发挥作用。随后有研究报道了miR-1和miR-206具有相似的种子序列,它们直接靶向牛骨骼肌卫星细胞中配对盒基因7(paired box 7,Pax7)和组蛋白去乙酰化酶4(histone deacetylase 4,HDAC4)来促进细胞成肌分化。在抑制miR-1和miR-206后,可提高Pax7和HDAC4蛋白水平,还能显著促进卫星细胞的增殖[21]。另有研究表明,日本短毛肉牛在圈养期间,股二头肌中miR-206的表达会逐渐下降,而放牧会维持肌肉组织中的miR-206水平,这也表明miR-206参与了放牧诱导的肉牛肌肉类型转化过程[22]。Dai等[23]通过微阵列芯片和qRT-PCR发现miR-128在牛骨骼肌中高表达,同时在牛骨骼肌卫星细胞分化过程中上调。进一步研究发现miR-128通过靶向抑制转录因子-刺激蛋白1(stimulatory protein 1,Sp1),从而抑制牛肌肉卫星细胞增殖和分化。Sp1同时也是成肌分化因子(myogenic differentiation,MyoD)的激活因子和细胞周期素依赖性激酶抑制因子1A(cyclin-dependent kinase inhibitor 1A,CDKN1A)的抑制因子。在肌肉细胞增殖和分化过程中差异变化的miRNA也是研究的聚焦点。miR-483在牛成肌细胞的增殖和分化阶段的表达量呈现下降趋势,miR-483靶向胰岛素样生长因子1(insulin-like growth factor 1,IGF1),并下调磷酸肌醇3激酶/蛋白激酶(PI3K/AKT)信号通路中关键蛋白的表达,对牛成肌细胞增殖和分化都具有负调控作用[24]。而miR-143表达量在牛骨骼肌卫星细胞分化过程中显著升高,胰岛素样生长因子结合蛋白5(insulin-like growth factor binding protein 5,IGFBP5)作为IGF信号通路的重要组成部分,它在牛骨骼肌卫星细胞中直接受到miR-143的靶向调控。转染miR-143模拟物可以导致IGFBP5的表达水平降低,抑制细胞增殖和分化;转染抑制剂下调miR-143表达量也能抑制细胞增殖,但能促进细胞分化[25]。由此可见,特异性高表达和阶段差异性表达的miRNA更有可能参与肉牛肌肉发育过程,后期研究可通过生物信息学等手段充分挖掘高表达和差异表达的miRNA的靶向基因并进行验证。
最近的研究表明,与miR-143类似,miR-365-3p在肌肉细胞也具增殖和分化双重调节功能[26],进一步功能分析发现,miR-365-3p能抑制牛原代成肌细胞增殖,但促进其分化(肌管形成)过程。其分子机制是miR-365-3p直接靶向激活素A受体Ⅰ型(activin A receptor Ⅰ,ACVRⅠ),从而降低细胞增殖相关分子细胞周期蛋白D1(cyclinD1,CCND1)、细胞周期蛋白依赖激酶2(cyclin-dependent kinase 2,CDK2)和增殖细胞核抗原(proliferating cell nuclear antigen,PCNA)的表达,同时促进肌肉分化标记基因(myogenic differentiation 1,MYOD1)和肌细胞生成素(myogenin,MYOG)的表达水平。此外,miR-378和miR-139可以通过分别靶向DNA聚合酶α亚基B(DNA polymerase α subunit B,POLA2)和二氢叶酸还原酶(dihydrofolater eductase,DHFR)促进牛骨骼肌卫星细胞的分化[27-28]。Zhang等[29]研究证实了miR-2400靶向MYOG促进牛骨骼肌卫星细胞增殖;Wang等[30]发现miR-208b可以提高CCND1、细胞周期蛋白E1(CyclinE1)的mRNA和蛋白表达水平,通过靶向负调控CDKN1A,促进牛原代成肌细胞的增殖。综上所述,不同miRNA因靶向作用的主效基因不同,对肉牛肌肉细胞的功能作用存在差异。在研究肉牛肌肉细胞功能基因变化的同时,关注miRNA等调控因子的变化规律,是深入探索其分子机制的有效途径。
肌肉嫩度、肌纤维类型与牛肉肉品质之间密切联系。Kappeler等[31]比较了不同剪切力的牛肌肉中miRNA表达量的差异,发现在剪切力较低的骨骼肌中miR-182和miR-183高表达,而剪切力较高的骨骼肌中miR-338高表达。Muroya等[32]比较了日本黑牛腱肌(MyHC2x型丰度,快肌)和咀嚼肌(MyHC1型丰度,慢肌)之间差异的miRNA,结果表明miR-196a和miR-885仅在腱肌肌肉中表达,而miR-208b在咀嚼肌中高表达,这些差异表达的miRNA可能在牛肌肉类型特异性组织形成和维持方面发挥关键作用。上述研究大体揭示了miRNA在不同牛肉品质条件下的差异规律,但目前还没有报道指出这些差异miRNA形成的生物学基础和参与的分子机制,因此有必要在今后的研究中加以明确。
3.2 lncRNA与肉牛肌肉发育
肉牛肌肉中lncRNA表达规律的挖掘分析有较为广泛的研究。Jia等[33]对成年夏洛来牛和同江牛的背最长肌的lncRNA进行高通量测序,共鉴定得到4 155个不同的lncRNA,其中有32个lncRNA在这2种肉牛背最长肌存在差异表达。在哈萨克牛和新疆褐牛背最长肌中,有182个lncRNA存在差异表达[34]。而在分析肉牛和广西水牛骨骼肌组织中lncRNA的表达情况时候共发现16 236个候选lncRNA,其中差异的lncRNA共有2 161个[35]。综合以上研究,我们可以发现不同品种肉牛肌肉中lncRNA的表达规律存在差异,这可能与不同肉牛之间肉质性状的差异存在紧密的联系。
越来越多的证据表明,lncRNA是肉牛骨骼肌发育的重要调控因子,通过相应的miRNA调控牛骨骼肌发育过程。Yue等[36]利用他们课题组前期对肉牛骨骼肌转录组高通量测序的结果,构建了lncRNA-miRNA-mRNA的ceRNA调控网络,发现lncRNA在肉牛肌肉发育相关的生物学过程和Wnt信号通路中显著富集。Sun等[37]对肉牛3个不同发育阶段(胎儿期、青春前期和体成熟期)骨骼肌中lncRNA的表达情况进行分析时发现401个lncRNA在3个发育阶段差异表达。其中包括一种新的肌肉特异性lncRNA(lncMD),在成肌细胞分化过程中上调。进一步研究表明,胰岛素样生长因子2(insulin-like growth factor 2,IGF2)是miR-125b的直接靶标,而lncMD可以作为miR-125b的分子海绵,导致IGF2表达增强,从而促进肌肉细胞分化。lncRNA-MEG3在牛骨骼肌组织中高表达,并在牛原代成肌细胞分化中保持稳定,过表达lncRNA-MEG3可以显著促进成肌细胞肌球蛋白重链(myosin heavy chain,MyhC)、MyoG和肌细胞增强因子2C(myocyte enhancer factor 2C,MEF2C)表达,其分子机制是lncRNA-MEG3可通过与miRNA-135和MEF2C相互作用促进牛骨骼肌细胞分化[38]。在对肉牛肌肉miRNA的总结中,已有研究表明miR-133是肌肉组织特异性高表达miRNA[20]。目前,发现lncRNA可以通过miR-133a和miR-133b来发挥作用。例如,lnc133b存在与成熟的miR-133b互补序列。lnc133b可以促进肌肉卫星细胞增殖并抑制其分化,下调miR-133b的表达,同时促进胰岛素样生长因子1受体(insulin-like growth factor 1 receptor,IGF1R)的表达,提示lnc133b通过ceRNA调控肌肉卫星细胞增殖和分化[39]。在肌肉发育过程中表达量变化显著的lnc-MDNCR可以通过海绵miR-133a,解除miR-133a对牛原代成肌细胞中的骨肉瘤B(osteosarcoma B,GosB)的靶向调控作用,从而促进成肌细胞分化并抑制细胞增殖[40]。
通过对基因组中相邻转录组进行转录干扰也是lncRNA的一种作用方式。在肉牛成肌细胞细胞核和肌管中特异性表达的lnc403可以负调控相邻基因——生肌调节因子6(myogenic regulatory factors 6,Myf6)的表达,从而抑制骨骼肌细胞的分化[41]。lncKBTBD10也是定位于细胞核中并影响其邻近基因——Kelch BTB域蛋白10(Kelch BTB domain protein 10,KBTBD10)参与肌发生过程。与其他基因的调控方式存在不同,无论对lncKBTBD10基因进行敲低或过表达,KBTBD10蛋白水平均降低,而且牛骨骼肌卫星细胞的增殖和分化都受到了抑制[42]。Yue等[43]发现在肉牛成肌细胞分化过程中lncYYW的表达逐渐增加,并促进成肌细胞增殖。机制上,lncYYW的过表达可以增加细胞周期DNA合成(S)阶段的细胞数量,促进肌原蛋白和肌球蛋白重链的表达,进一步的芯片分析显示lncYYW在牛成肌细胞中正调控生长激素1(growth hormone 1,GH1)及其下游基因AKT1的表达。lnc-H19已被发现在人类和小鼠中可以促进成肌细胞分化,Xu等[44]研究发现lnc-H19在1周龄及1、6和36月龄的牛骨骼肌中均高表达,并可以通过抑制沉默信息调节因子2相关酶1/叉头转录因子1(Sirt1/FoxO1)信号通路促进牛骨骼肌卫星细胞的分化。最近的研究发现lncRNA-IGF2 AS作为胰岛素样生长因子2(insulin-like growth factor 2,IGF2)的反义转录本,可促进牛成肌细胞的增殖和分化。lncRNA-IGF2 AS不仅可以补充IGF2的内含子区域,影响IGF2的稳定性和表达,还可直接与白细胞介素增强因子结合因子3(ILF3)蛋白结合,从而参与调控肌生成[45]。以上研究表明,lncRNA在调控肉牛骨骼肌发育过程的作用形式多样。在通过生物信息学分析lncRNA参与的ceRNA调控网络的同时,也需关注lncRNA的基因组中相邻转录组的表达情况,特别是与相应编码基因具有互补序列的反义lncRNA。
3.3 circRNA与肉牛肌肉发育
Liu等[46]使用RNA-Seq鉴定了山东黑牛和鲁西牛肌肉中的circRNA。在鉴定得到的14 640个circRNA中,差异表达circRNA共有655个。同时分析出有14个circRNA可能在肉牛肌肉生长发育调控中发挥重要作用。在哈萨克牛和新疆褐牛背最长肌circRNA的测序鉴定中共得到5 177个circRNA,其中有46个差异表达。进一步的生物信息学分析表明,差异表达circRNA的来源基因与肌肉的生物学过程有关[47]。这些差异表达的circRNA也可能是新疆褐牛牛肉品质更高的生物学基础。已有研究表明,circRNA参与了肉牛肌细胞增殖与分化过程。circEch1是肉牛肌肉发育过程中上调最多的circRNA之一,体外试验表明circEch1抑制牛成肌细胞的增殖和促进其分化,动物(小鼠)试验研究进一步确认circEch1可刺激骨骼肌再生[48]。更多的体外试验研究表明,circRNA可通过对相应miRNA的海绵作用来参与肉牛肌肉细胞发育过程。circINSR是肉牛胚胎肌肉组织中高表达的circRNA之一,通过miR-34a调控B淋巴细胞瘤-2(B-cell lymphoma-2,Bcl-2)和细胞周期蛋白E2(CyclinE2)的表达来促进肉牛成肌细胞增殖,减少细胞凋亡[49]。miR-133a是肉牛肌肉组织中高表达的miRNA,除了与lncRNA具有结合位点,Li等[50]发现到circFUT10具有miR-133a 3个结合位点,并通过竞争性结合miR-133a,增加MyoD、MyoG和MyhC的蛋白表达水平,诱导牛原代成肌细胞分化,抑制增殖并促进凋亡。miR-107同样在肉牛骨骼肌中高表达,miR-107可以靶向抑制Wnt3a,进而抑制牛成肌细胞的分化,circFGFR4可以通过海绵miR-107改变这一过程来促进成肌细胞的分化[51]。circSNX29也可促进肉牛成肌细胞分化,抑制细胞增殖,其分子机制是circSNX29直接靶向下调miR-744,逆转miR-744对Wnt5a和CaMKⅡ δ的抑制,从而激活了非典型的Wnt5a/Ca2+信号通路[52]。circHUWEl在肉牛成肌细胞中可以通过竞争性结合miR-29b,解除其对AKT的抑制作用,从而激活AKT信号通路,促进牛成肌细胞增殖,抑制细胞凋亡和分化[53]。miR-432作为IGF2的调节因子,circTTN可竞争性结合miR-432,激活IGF2-PI3K-AKT信号通路,促进牛原代成肌细胞的增殖和分化[54]。
综上所述,ncRNA在肉牛肌肉组织中表达丰富,通过不同的靶基因作用和信号通路途径,广泛参与了肉牛肌肉细胞的增殖和分化调控过程,并与肌肉剪切力和嫩度等肉质性状密切相关。表1总结了近年来ncRNA在肉牛肌细胞发育过程中的生物学功能。

表1 ncRNAs在肉牛肌肉发育中的功能Table 1 Function of ncRNAs in muscle development in beef cattle
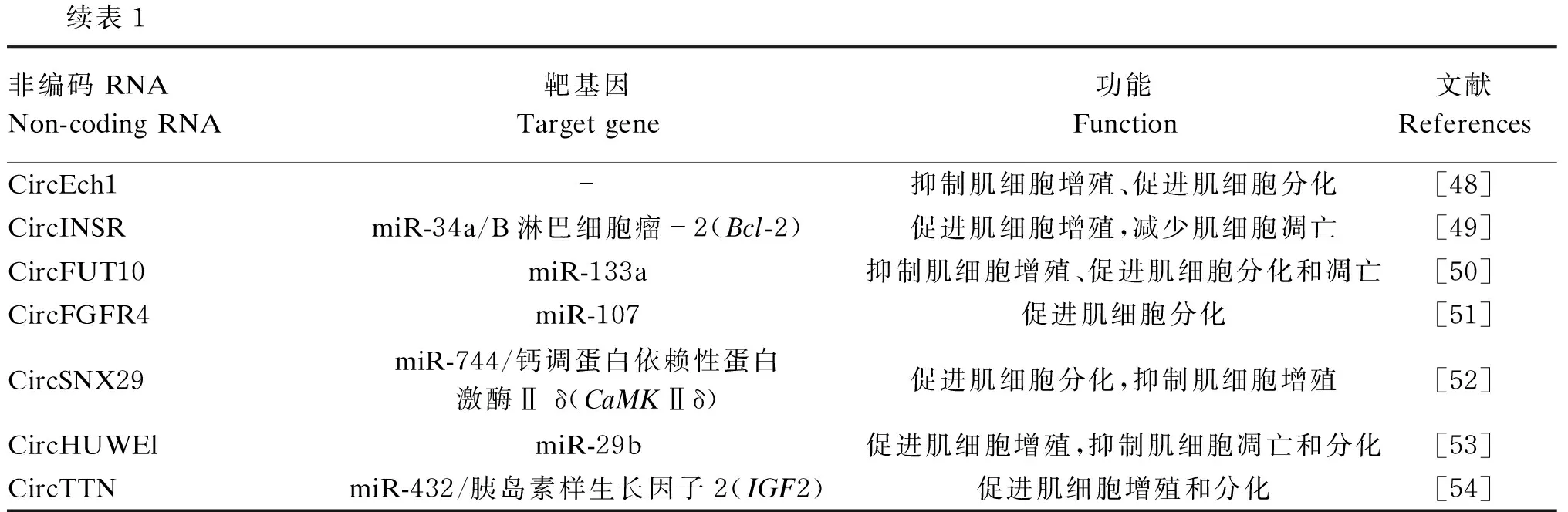
续表1非编码RNANon-coding RNA靶基因Target gene功能Function文献ReferencesCircEch1-抑制肌细胞增殖、促进肌细胞分化[48]CircINSRmiR-34a/B淋巴细胞瘤-2(Bcl-2)促进肌细胞增殖,减少肌细胞凋亡[49]CircFUT10miR-133a抑制肌细胞增殖、促进肌细胞分化和凋亡[50]CircFGFR4miR-107促进肌细胞分化[51]CircSNX29miR-744/钙调蛋白依赖性蛋白激酶Ⅱ δ(CaMKⅡδ)促进肌细胞分化,抑制肌细胞增殖[52]CircHUWElmiR-29b促进肌细胞增殖,抑制肌细胞凋亡和分化[53]CircTTNmiR-432/胰岛素样生长因子2(IGF2)促进肌细胞增殖和分化[54]
4 ncRNAs与肉牛脂肪沉积
4.1 miRNA与肉牛脂肪沉积
分析肉牛不同部位脂肪组织中ncRNA的表达差异可为肉牛脂肪沉积的靶向调控提供新途径。Wang等[55]通过微阵列芯片和生物信息学研究了成年肉牛肌内脂肪和皮下脂肪中miRNA的表达谱,在芯片检测到的213个miRNAs中有88个差异表达的miRNAs。miR-143、miR-145、miR-26a、miR-2373-5p和miR-23b-3p在肌内脂肪中高表达,而miR-26a、miR-2373-5p、miR-2325c、miR-3613和miR-2361在皮下脂肪中高表达;这些差异表达的miRNAs的靶基因参与了不同的信号通路。肌内脂肪作为牛肉品质评价中最重要的指标,在不同肌内脂肪含量的肉牛组织中,其miRNA的表达谱也存在较大差异[56-57]。肉牛脂肪组织中的miRNA的表达谱受多种因素的影响,包括品种[58]、饲粮营养水平[59-60]、饲喂和放牧方式[61]等。阉割作为增加肉牛肌内脂肪沉积,改善肉质的途径之一,同时也会导致肌内脂肪中miRNA表达谱改变。Zhang等[62]对秦川公牛和阉牛肌内脂肪中的miRNA进行分析共鉴定出52个差异表达的miRNA。另有研究证明,阉牛体内组织中miR-27a-5p更高,其可以通过靶向牛体内钙敏感受体(calcium-sensing receptor,CaSR)来增加脂肪沉积,这也可能是阉牛具有更高品质肉质的原因之一[63]。
miRNA广泛参与了肉牛脂肪沉积的调控过程。Yu等[64]为筛选肉牛脂肪形成相关的miRNA,分析了脂肪前体细胞和成熟脂肪细胞(分化后)miRNA的表达谱,发现共有260个差异的miRNA,功能分析表明差异miRNA的靶基因和脂质代谢密切相关。本文总结了近年来对肉牛脂肪细胞具有调控作用的miRNAs。miR-130a和miR-130b都可以调控牛脂肪细胞分化,miR-130a和miR-130b分别通过靶向抑制过氧化物酶体增殖激活受体(peroxisome proliferator activated receptor gamma, PPARγ)和细胞色素P450家族2亚家族U成员1(cytochrome P450 family 2 subfamily U member 1,CYP2U1)导致脂肪细胞中甘油三酯(triglyceride,TG)水平显著降低,并减少脂滴形成[65]。在肉牛胎儿期脂肪细胞分化过程中,miR-210通过靶向沉默Wnt1诱导信号通路蛋白2(Wnt1 inducible signaling pathway protein 2,WISP2)促进细胞的脂肪生成[66]。延边黄牛作为我国五大良种黄牛之一,miR-1271在延边黄牛脂肪细胞中直接靶向激活转录因子3(activating transcription factor 3,ATF3)的3′-UTR,并下调其表达,促进脂肪形成调节因子PPARγ和CCAAT增强子结合蛋白(CCAAT enhancer binding protein alpha,C/EBPα)的表达,从而增加甘油三酯积累和细胞聚脂[67]。miR-149-5p在牛脂肪细胞增殖和分化后期表达量上调,直接靶向CREB调节转录辅激活因子(CREB regulated transcription coactivator,CRTCs)抑制脂肪细胞增殖和脂肪积累[68]。比较转录组分析发现miR-224与脂蛋白脂酶(lipoprotein lipase,LPL)在肉牛肌内脂肪中呈现负相关,进一步的研究证实了miR-224靶向负调控LPL,同时会导致PPARγ、脂肪酸合酶(fatty acid synthase,FASN)、围脂滴蛋白1(perilipin 1,PLIN1)等脂肪形成相关基因的表达水平降低,从而抑制牛前体脂肪细胞的成脂分化[69]。与miR-224的调控功能相反,miR-378可促进牛前体脂肪细胞的分化聚脂[70]。E2F转录因子2(E2F transcription factor 2,E2F2)和RAN结合蛋白10(RAN binding protein 10,RANBP10)都是miR-378靶基因。在肉牛脂肪细胞中过表达miR-378,2个靶基因表达下调,而脂肪细胞分化标记基因PPARγ、固醇调节元件结合蛋白(sterol regulatory element binding protein,SREBP)表达上调,从而导致甘油三酯含量、脂滴体积增加。综上所述,miRNA与肉牛脂肪细胞的增殖、分化聚脂密切相关,但需要对肉牛不同部位脂肪组织中miRNA的表达规律及调控机制进行研究,从而为肉牛不同部位差异化脂肪沉积调控提供靶点和基础资料。
4.2 lncRNA、circRNA与肉牛脂肪沉积
在肉牛脂肪沉积的ncRNA的研究中,lncRNA和circRNA的研究报道相对较少。Jiang等[70]通过测序分析了秦川犊牛和成年牛脂肪组织中lncRNA表达情况,共获得3 716个候选lncRNA,在2个发育阶段有119个lncRNA差异表达。他们同时对2种不同生长阶段肉牛脂肪组织中circRNA表达情况进行了分析,共获得14 274个候选circRNA,其中有307个circRNA差异表达[71]。Li等[72]分析了牛原代脂肪细胞分化前后lncRNA的表达变化,对差异表达最显著的lncRNA ADNCR功能研究发现其抑制脂肪细胞分化标志性基因的表达。机制分析揭示lncRNA ADNCR能够作为竞争性内源RNA吸附miR-204,阻止miR-204对其靶基因-组蛋白去乙酰化酶1(sirtuin 1,SIRT1)的抑制作用,从而抑制脂肪生成。Lnc-MIR221HG作为牛转录中新发现的lncRNA,主要存在于细胞核内并能抑制牛脂肪细胞分化[73]。Lnc-BADLNCR1同样是牛脂肪细胞分化过程中的调节因子,谷氧还蛋白5(glutaredoxin 5,GLRX5)是促进脂滴形成和成脂基因表达的刺激因子,Lnc-BADLNCR1通过负调控GLRX5基因的表达,从而实现抑制牛脂肪细胞分化聚脂[74]。通过对miRNA的海绵作用是lncRNA和circRNA的共同作用方式。Kang等[75]发现circFLT1和lncCCPG1在肉牛前体脂肪细胞和成熟脂肪细胞中的差异化表达,过表达circFLT1和lncCCPG1共同促进脂肪细胞分化并抑制增殖。他们还研究了circFLT1和lncCCPG1在这一生物学过程背后所涉及的复杂分子机制。他们发现circFLT1和lncCCPG1都能对miR-93形成海绵作用,解除miR-93对lncSLC30A9的表达的抑制作用,高表达的lncSLC30A9通过抑制AKT蛋白的表达来抑制细胞增殖,同时通过将FOS蛋白招募到PPARγ的启动子上促进脂肪细胞分化。Jiang等[71]在前期高通量测序的基础上,发现circFUT10的来源基因与脂肪组织发育和沉积密切相关,并在脂肪组织细胞中高表达。功能分析表明circFUT10通过对let-7c的海绵作用,解除let-7c对过氧化物酶体增殖激活受体-γ共激活因子1β(peroxisome proliferation-activated receptor-γ coactivator 1β,PPARGC1B)的抑制,从而促进牛脂肪细胞增殖,抑制细胞分化。
综上所述,研究肉牛脂肪细胞的增殖和分化的机理是改善肉牛体脂沉积和生产优质牛肉的基础。与肉牛肌肉发育的研究相比,ncRNA在脂肪沉积的研究相对薄弱,但也发现了许多ncRNA对肉牛脂肪细胞的增殖和分化具有调控作用,如表2所示。

表2 ncRNAs在肉牛脂肪沉积中的功能Table 2 Function of ncRNAs in fat deposition in beef cattle

续表2非编码RNANon-coding RNA靶基因Target gene功能Function文献ReferencesmiR-378E2F转录因子2(E2F2)/RAN结合蛋白10(RANBP10)促进脂肪细胞成脂分化[70]LncRNA ADNCRmiR-204/组蛋白去乙酰化酶1(SIRT1)抑制脂肪细胞聚脂[72]Lnc-MIR221HG-抑制脂肪细胞分化[73]Lnc-BADLNCR1谷氧还蛋白5(GLRX5)抑制脂肪细胞分化聚脂[74]LncCCPG1miR-93促进脂肪细胞分化、抑制脂肪细胞增殖[75]CircFLT1miR-93促进脂肪细胞分化、抑脂肪细胞制增殖[75]CircFUT10let-7c/过氧化物酶体增殖激活受体-γ共激活因子1β(PPARGC1B)进促脂肪细胞增殖,抑制脂肪细胞分化[71]
5 小结与展望
目前,ncRNAs在畜牧生产上的研究发展迅速,研究者通过系统的生物学手段筛选出大量参与肉牛肌肉细胞和脂肪细胞增殖和分化过程的ncRNAs,为肉牛的高产优质的营养调控和育种改良提供了理论基础和丰富的调控靶点。随着生物学技术的持续发展和ncRNA数据库不断完善、规范,研究人员将会发现更多参与肉牛生长发育过程的ncRNAs,但是仍然存在以下科学问题有待探索:1)与miRNA相比,lncRNA和circRNA的功能更为广泛,但由于在物种之间保守性差,它们在肉牛中的研究还需要更多的挖掘和验证,尤其是与肉牛脂肪沉积相关的circRNA研究。同时也需要尽快地规范lncRNA和circRNA的命名、完善相关数据库。2)由于肉牛属于大型动物的限制,目前关于非编码RNA在肉牛肌肉和脂肪中的功能验证基本集中在体外细胞水平,缺乏活体试验的验证研究。miRNA在不同动物之间保守性强,并具有相似的生物学功能,通过基因敲除、慢病毒载体等手段在模式动物(例如小鼠)上进行ncRNA功能的活体验证是今后可考虑的研究方法。3)肌纤维类型和肌内脂肪是影响牛肉品质的主要因素,在不同类型肌纤维的形成和转化、不同部位的脂肪沉积过程中ncRNA的变化规律、生物学功能和相应的分子机制的研究应当值得关注。4)饲粮营养作为动物生长的物质基础,在肌肉发育和脂肪沉积过程中具有决定性的作用。在研究不同营养物质和配比对肉牛肌肉和脂肪中关键主效基因产生影响的同时,也应关注相应功能性ncRNA的变化规律,从而为精细化营养调控肉牛的高产优质提供基础。
猜你喜欢
今日农业(2022年2期)2022-11-16
今日农业(2022年1期)2022-06-01
昆明医科大学学报(2021年10期)2021-12-02
今日农业(2021年21期)2021-11-26
天然产物研究与开发(2018年2期)2018-04-04
中国运动医学杂志(2016年3期)2016-07-10
中国运动医学杂志(2016年3期)2016-07-10
医学研究杂志(2015年5期)2015-06-10
当代畜禽养殖业(2014年6期)2014-02-27
中国病理生理杂志(2012年10期)2012-07-31
