
我国部分地区猪繁殖与呼吸综合征病毒遗传变异分析
2021-10-15 12:30梁俊超刘晓东刘红祥马振乾胡继明吉林大学动物医学学院人兽共患病研究所人兽共患病研究教育部重点实验室吉林长春13006动物基因工程疫苗国家重点实验室青岛易邦生物工程有限公司山东青岛66000
家畜生态学报 2021年9期
梁俊超,刘晓东,袁 飞,李 坤,刘红祥,马振乾,胡继明,刘 东*,于 录(1.吉林大学 动物医学学院人兽共患病研究所/人兽共患病研究教育部重点实验室,吉林 长春 13006; .动物基因工程疫苗国家重点实验室/青岛易邦生物工程有限公司,山东 青岛 66000)
猪繁殖与呼吸综合征(porcine reproductive and respiratory syndrome,PRRS)是一种急性、高度接触性传染病,该病病原为猪繁殖与呼吸综合征病毒(porcine reproductive and respiratory syndrome virus,PRRSV),临床发病主要以繁殖障碍及呼吸道症状为主,多表现为公猪精液质量差,母猪流产、产死胎及产后发情不规则,商品猪高烧、呼吸障碍且死淘率高[1-3]。
1987年,美国首次报道发现PRRS[4],1996年我国首次报道从疑似病例中分离得到PRRSV[5],从而也证实PRRS已传入我国。2006年后,我国出现高致病性PRRSV(highly pathogenic PRRSV,HP-PRRSV),该类毒株毒力增强,造成养猪业重大死伤,导致严重经济损失[6]。2010年,广东某猪场仔猪血清中分离得到GM2毒株,GM2毒株以QYYZ毒株和MLV毒株为亲本,但在猪体内的复制能力高于亲本,致病性相对更高[7]。2013年我国已有报道猪群因感染NADC 30-like毒株导致重胎期母猪大面积流产[8]。2014年后,NADC30-like毒株临床检出率逐年升高,国内流行态势加剧,且在某些区域成为优势毒株[9]。目前,国内田间流行蓝耳野毒毒株愈加复杂多样,这也为我国蓝耳病防控带来全新挑战。
GP5蛋白是由PRRSV的ORF5基因编码的高度糖基化的囊膜蛋白,PRRSV的主要中和位点位于GP5蛋白[10]。GP5蛋白是PRRSV中变异程度最大的结构蛋白之一,通常作为 PRRSV变异的主要依据之一,而GP5基因序列的变化可在一定程度反映蓝耳流行毒株的遗传变异情况[11]。因基因组差异,PRRSV被分为欧洲型(基因Ⅰ型,代表毒株LV株)和北美型(基因Ⅱ型,代表毒株VR-2332株)。基于全球PRRSV分类系统和GenBank中的ORF5序列信息,我国普遍存在的北美型(基因Ⅱ型)PRRSV毒株被分为四大谱系:谱系1,谱系3,谱系5和谱系8,谱系1代表毒株为NADC30和MN184,谱系3代表毒株为GM2,谱系5代表毒株为经典毒株VR2332和MLV,而谱系8又被细分为5个亚群,亚群Ⅰ代表毒株为经典毒株CH-1a,亚群Ⅱ为经典和HP-PRRSV的中间亚群,代表毒株为BJ0706,亚群Ⅲ-Ⅴ为HP-PRRSV及其衍生株,代表毒株为JXA1、TJ和HuN4[12]。
对2019年我国8省46份PRRSV阳性样品中PRRSV毒株进行ORF5基因测序,并比对分析PRRSV遗传进化,为以后PRRS的流行趋势及防控提供理论依据和参考。
1 材料与方法
1.1 病毒样品
2019年从我国8个省(包括山东、江苏、安徽、宁夏、辽宁、云南等)采集的疑似 PRRSV感染的临床样品按照文献[13]介绍的RT-PCR方法进行检测。利用病毒RNA提取试剂盒(TAKARA公司)提取总RNA,并用RT试剂盒(罗氏公司)进行反转录反应。
利用ORF5基因引物(上游:CATTTCATGACACCTGAGACCA;下游:AGAGCATATATCATCACTGGCG)通过PCR进行扩增。扩增条件如下:94 ℃变性2 min后进入循环:94 ℃15 s,54 ℃30 s,68 ℃1 min,32个循环后,68 ℃10 min。
1.2 主要试剂
RNA提取试剂盒购于TAKARA公司;高保真反转录试剂盒购于罗氏公司;LATaq DNA聚合酶购于宝生物工程(大连)有限公司。
1.3 PRRSV GP5测序
将RT-PCR阳性产物送至生工生物工程(上海)股份有限公司进行测序。
1.4 分子进化分析
通过序列分析软件DNAstar Lasergene对所选毒株的ORF5基因进行序列同源性的比对分析,利用MEGA6软件对所测ORF5序列制作进化树。
2 结果与分析
2.1 RT-PCR检测及测序结果
对送检样品的ORF5基因进行 RT-PCR检测,目的条带大小为603 bp,出现目的条带表明所检样品PRRSV为阳性(图1)。从8省送检的PRRSV抗原阳性样品中,随机挑选46份PRRSV的PCR产物进行ORF5基因测序,经比对后确认得到46个PRRSVORF5基因序列,各省PRRSV样品测序数量详见表1。
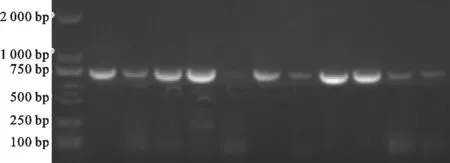
图1 PRRSV RT-PCR检测结果Fig. 1 Results of PRRSV RT-PCR detection
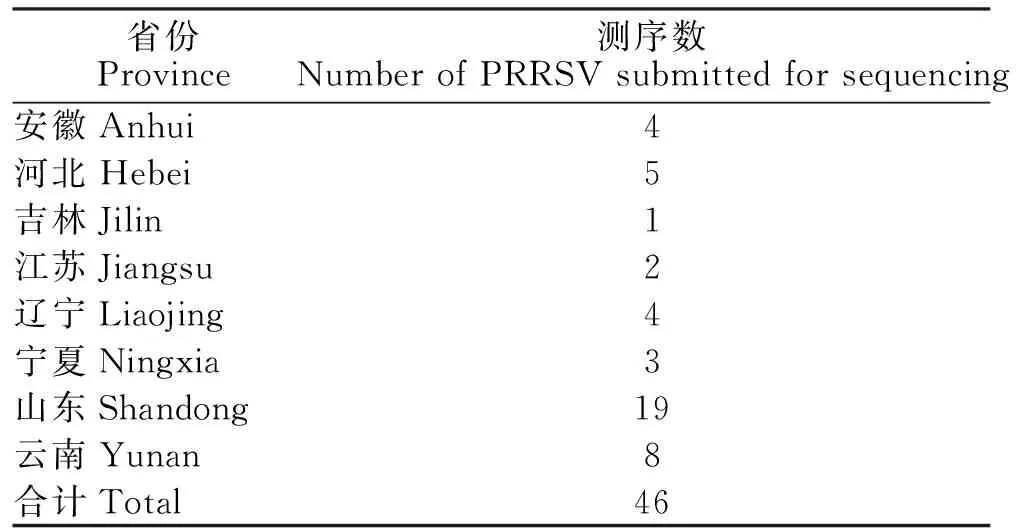
表1 PRRSV送检测序数量及区域分布Table 1 The number and regional distribution of PRRSV submitted for sequencing
2.2 PRRSV ORF5基因的核苷酸同源性比对及进化分析
样品序列结果按照“送检年月-样品编号-送检省份缩写”进行编号。46份样品的PRRSVORF5基因扩增产物经测序所得46个603 bp的ORF5基因序列,与谱系1、3、5和8的标准毒株通过DNAstar和MEGA6软件进行比对,与标准毒株同源性比对结果见表2。46个PRRSVORF5序列与标准毒株的进化树见图2。由表3可见,8个省46个测序样品的PRRSV分布在谱系1、谱系3和谱系8中,其中谱系1有19个,谱系3仅2个,谱系8有25个且全属于亚群Ⅲ-Ⅴ。山东1~6月份谱系1中样品PRRSV仅2个,而7~12月份为10个,NADC 30-like毒株检出量明显增多,而谱系8中样品在1~6月份和7~12月份检出量无明显差异。
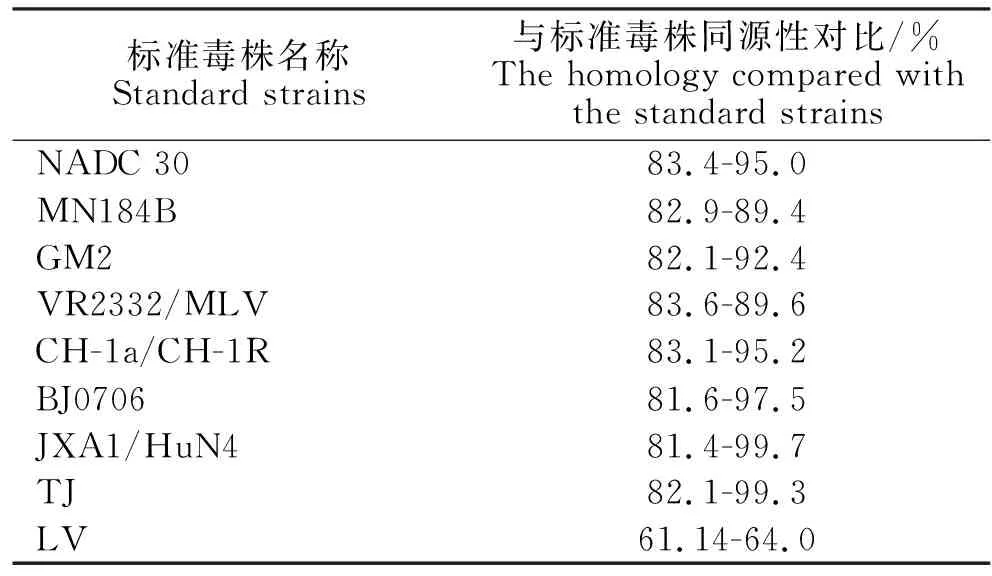
表2 46个PRRSV ORF5基因序列与标准毒株同源性对比Table 2 The homology of 46 PRRSV ORF5 gene sequences compared with the standard strains

表3 46个PRRSV ORF5基因序列谱系及区域分布Table 3 Lineage and regional distribution of 46 PRRSV ORF5 gene sequences
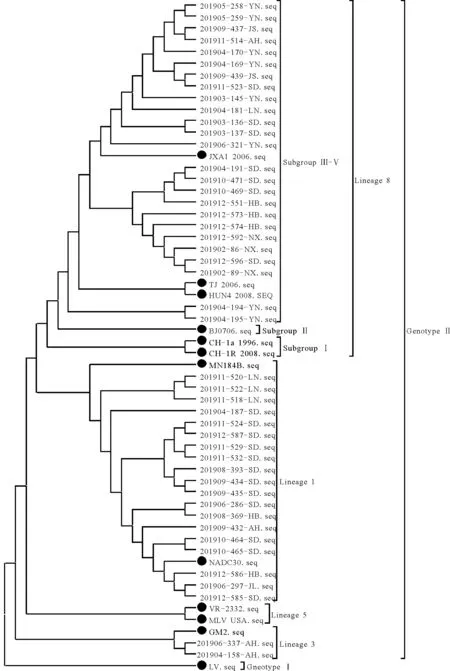
图2 46个PRRSV ORF5基因序列进化树分析Fig. 2 Evolutionary tree analysis of 46 PRRSV ORF5 gene sequences
2.3 PRRSV GP5蛋白氨基酸变异分析
46个PRRSV GP5蛋白的氨基酸序列主要以点突变为主,未见插入突变,氨基酸同源性分析结果见图3。美洲型PRRSV毒株GP5蛋白有2个重要的表位结构,分别是诱饵表位(27~30 aa)和中和表位(37~45 aa),其中诱饵表位是非中和表位[14-16]。在诱饵表位区域,谱系1中14个样品PRRSV在第27位氨基酸发生变异V27A,2个样品PRRSV在第28位氨基酸发生变异L28P和L28F,4个样品PRRSV在第30位氨基酸发生变异N30S,3个样品PRRSV在第29和30位氨基酸发生变异V29A和N30G;谱系3中2个样品PRRSV在第27和30位氨基酸发生变异V27A和N30S;谱系8中2个样品PRRSV在第30位氨基酸发生变异N30D。
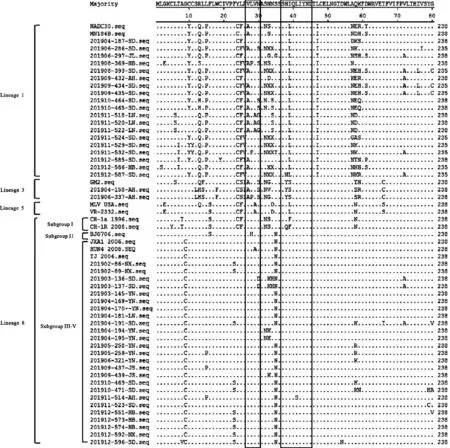
图3 46个PRRSV GP5蛋白氨基酸同源性分析Fig. 3 Amino acid homology analysis of 46 PRRSV GP5 proteins
在中和表位区域,谱系1中19个样品PRRSV在第39位氨基酸均发生变异I39L,其中201912-587-SD毒株在38位氨基酸还发生变异H38N,谱系3中2个样品PRRSV均在第38和39位氨基酸发生变异H38Y和I39S,而谱系8中,201911-514-AH在第41位氨基酸发生变异L41S。
GP5具有多个糖基化位点,N32或N33或N34位于膜外结构域的高变区且只存在于部分毒株中,此外,N44和N51也是糖基化位点[17]。46个PRRSV的GP5蛋白的N-糖基化位点N44N51未发生变化,而不同PRRSV的N-糖基化位点N33N34发生明显变异。谱系1代表毒株NADC 30为N32N34,MN184B为N34,19个样品PRRSV中,8个PRRSV为N32,2个为N33N34,2个为N34,1个为N32N34,2个为N32N33,3个为N33,而21912-586-HB无N33N34;谱系3中代表毒株GM2和2个样品PRRSV均为N32N34;谱系8中代表毒株JXA1、HuN4和TJ均为N33N34N35,25个样品毒株中,20个为N33N34N35,2个为N35,2个为N32N34,1个为N34N35。
3 讨 论
PRRSV是我国养猪业重要防控疾病之一,由于PRRSV基因组易变异,导致新基因型毒株不断出现。随着田间流行野毒毒株多样化,疫苗毒株与野毒毒株同源性差异加大,而野毒毒株抗原表位及中和表位出现突变,最终将导致目前商品化PRRS疫苗免疫效果受限[18-20]。
8省共46个样品PRRSVORF5基因送经测序可知,山东、辽宁、河北的样品PRRSV分布在谱系1和谱系8中,安徽的样品PRRSV分布在谱系1、3和8中,江苏、宁夏、云南的样品PRRSV分布在谱系8中,且均为HP-PRRSV,吉林仅测一个且在谱系1中,谱系1中样品PRRSV共19个,谱系8中样品PRRSV共25个,该结果表明国内HP-PRRSV虽仍占一定优势,但NADC 30-like毒株流行已明显加剧。
山东送检测序共19个品PRRSV,其中12个属于谱系1,7个属于谱系8,且在7~12月份NADC 30-like毒株检出明显高于1~6月份,该结果表明山东NADC 30-like毒株流行相比HP-PRRSV呈现加剧趋势,且下半年流行态势高于上半年。
46个样品毒株与经典PRRS疫苗毒株如VR2332、MLV及CH-1R同源性相对较远,理论上PRRSV同源性高低影响PRRS疫苗毒株的田间保护效果[21]。GP5蛋白诱饵表位相对于中和表位是优势表位,会优先被机体识别,对中和表位有屏蔽作用,导致中和抗体产生延迟,有利于PRRSV逃避机体免疫清除而大量增殖[22]。46个样品PRRSV中共有17个PRRSV诱饵表位发生变异,氨基酸变异位点分别为V27A、L28P、L28F、V29A、N30S、N30G和N30D,诱饵表位变异可能有利于病毒免疫逃避。GP5蛋白中和表位能够诱导机体产生中和抗体,而该蛋白免疫识别位点的变异可影响中和抗体效果及疫苗的交叉保护率[10,23],46个样品PRRSV中22个PRRSV的中和表位发生变异,氨基酸变异位点分别为H38N、H38Y、I39L、I39S和L41S,22个样品分别属于谱系1、3和8。现在国内流行的NADC-like毒株、GM2-like毒株及HP-PRRSV毒株的中和表位确存在变异而且变异位点存在差异,这可能导致疫苗对田间不同流行野毒毒株的交叉保护力下降。
糖基化屏蔽是PRRSV一种免疫逃避机制,可避免被机体内中和抗体清除,而GP5膜外区多糖位点缺失能够提高抗体的中和能力和其附近中和表位的免疫原性[17,24]。46个样品PRRSV的N-糖基化位点N44N51均未发生变化,而N33N34发生不同变异,其中8个样品PRRSV为N32,2个为N33N34,2个为N34,5个为N32N34,2个为N32N33,3个为N33,20个为N33N34N35,2个为N35,1个为N34N35,1个为N33N34。20个样品PRRSV为HP-PRRSV,且多糖位点增加,这可能降低高中和表位免疫原性及抗体中和能力;14个样品PRRSV为NADC30-like PRRSV,且多糖位点减少,这可能提高高中和表位免疫原性及抗体中和能力。
4 结 论
通过以上研究可知,我国现阶段流行的蓝耳毒株呈现多样化、复杂化,不同谱系毒株间诱饵表位、中和表位及糖基化位点均存在一定的差异,从而疫苗与野毒交叉保护力受限,最终导致单纯依赖疫苗免疫防控难度加大。结合临床,蓝耳病更需综合防控,首先要重视和提升猪场生物安全及饲养管理水平,然后了解猪场自身PRRSV感染实际情况,选择与猪场PRRSV同源性近、安全性高且免疫原性好的疫苗进行免疫,综合防控才能有效防控蓝耳病。
猜你喜欢
动物医学进展(2022年9期)2022-11-26
分子催化(2022年1期)2022-11-02
分析测试学报(2022年9期)2022-09-21
艺术品鉴(2022年16期)2022-07-09
科学大观园(2022年2期)2022-01-23
文萃报·周二版(2021年47期)2021-12-14
北方论丛(2021年2期)2021-05-22
景德镇陶瓷(2021年1期)2021-03-24
当代陕西(2019年19期)2019-11-23
生物学教学(2018年4期)2018-11-29
